Legionella pneumophila-Induced NETs Do Not Bear LL-37 Peptides
Valeria Iliadi, Stefania Marti, Aikaterini Skeva, Konstantinos Marmanis, Theofani Tsavdaridou, Georgios Euthymiou, Eleni Tryfonopoulou, Dimitrios Themelidis, Athina Xanthopoulou, Katerina Chlichlia, Maria Koffa, Theocharis Konstantinidis, Maria Panopoulou

TL;DR
This study shows that neutrophils infected with Legionella pneumophila form extracellular traps (NETs), but these NETs lack the antimicrobial peptide LL-37, which may hinder bacterial elimination.
Contribution
The study reveals that LL-37 is absent on L. pneumophila-induced NETs, suggesting a failure in antimicrobial defense.
Findings
Neutrophils release NETs in response to L. pneumophila, but these NETs lack LL-37.
LL-37 is present on NETs during Clarithromycin treatment, inhibiting bacterial proliferation.
NETs formed during L. pneumophila infection fail to inhibit bacterial growth due to the absence of LL-37.
Abstract
Legionella pneumophila (L. pneumophila) infection is characterized by a wide spectrum of manifestations, from influenza-like illness to life-threatening atypical pneumonia with multiorgan failure. The aim of our study was the assessment of in vitro and ex vivo neutrophil activation in L. pneumophila infections, as well as the role of neutrophils’ peptides such as LL-37 in infection. The ability of neutrophils to form ex vivo extracellular traps (NETs) in response to bacterial infection was examined by immunofluorescence. In parallel, patients’ sera, as well as opsonized standard L. pneumophila strains, were used for in vitro activation of neutrophils from healthy individuals. The serum levels of interleukins were assessed using the LEGENDplexTM Multi-Analyte Flow Assay Kit. Furthermore, citrullinated cf-DNA as a marker of neutrophil extracellular traps (NETs) was detected in the serum…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
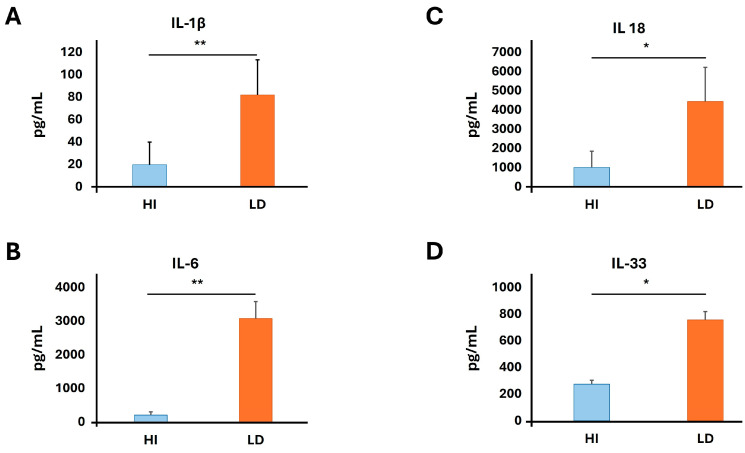 Figure 1
Figure 1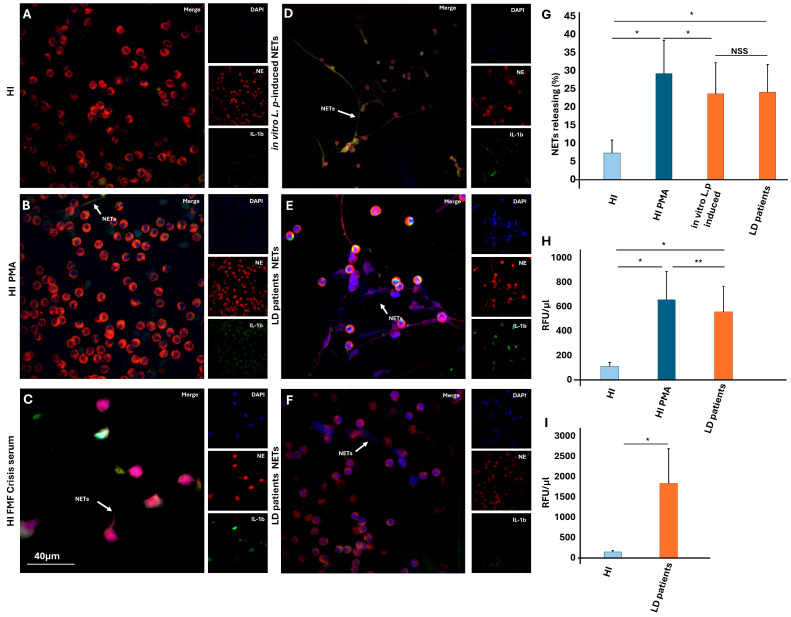 Figure 2
Figure 2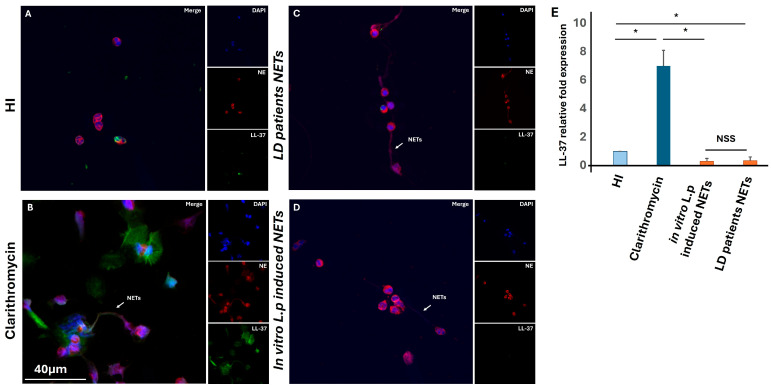 Figure 3
Figure 3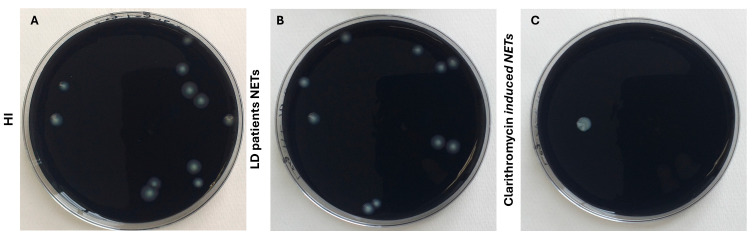 Figure 4
Figure 4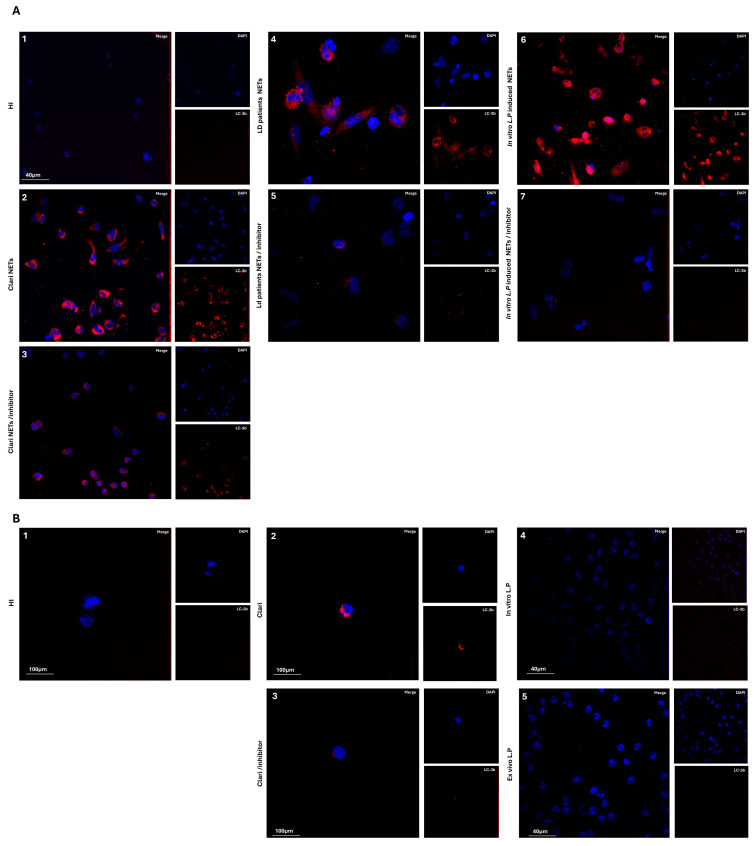 Figure 5
Figure 5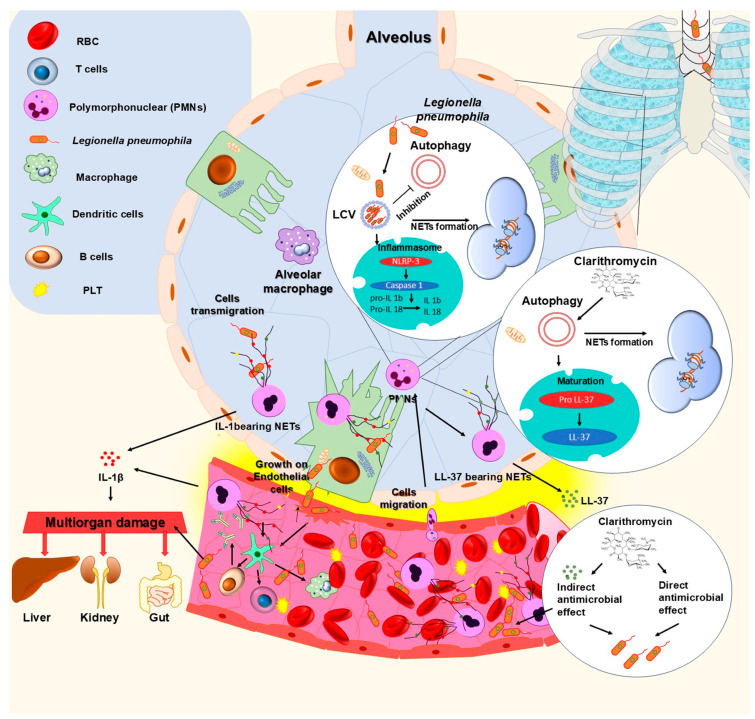 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLegionella and Acanthamoeba research · Neutrophil, Myeloperoxidase and Oxidative Mechanisms · Water Treatment and Disinfection
1. Introduction
The Legionella spp. was first described in 1947 as a “rickettsia-like” microorganism [1], and the first outbreak was reported during the 58th annual convention of the American Legion in Philadelphia [2]. Years later, those strains were characterized as etiological agents of airborne infections, and the microorganism was named Legionella pneumophila (L. pneumophila) [3]. It is a small, Gram-Negative, aerobic, non-sporogenous, non-capsule-forming and motile bacillus that belongs to the family Legionellaceae [4,5]. More than 50 species of Legionella spp., and more than 70 distinct serogroups, have been identified [6]. Legionella pneumophila is the most common pathogenic species, with L. pneumophila serogroup 1 (Lp1) being one of the most commonly detected serotypes that causes outbreaks worldwide [7]. The bacterium is encountered in the environment, in nature as well as in man-made sources, such as air-conditioning systems and car cabin air filters [8,9]. Infection due to L. pneumophila is owed to inhalation of bacteria-contaminated aerosols [10]. The clinical manifestation of infection has two distinct forms: non-pneumonic, a milder febrile flu-like syndrome named Pontiac Fever, and a pneumonic form called Legionnaires’ disease (LD), a severe pneumonia with extrapulmonary manifestations and multiorgan failure, often leading to a life-threatening state and then death [11,12,13].
After inhalation, L. pneumophila arrives at alveolar space, where it enters into the alveolar macrophages. [14]. In addition, bacterial strains may penetrate alveolar epithelial cells by different Legionnaires’ virulence factors [15]. Both alveolar macrophages and epithelial cells, as well as the same microbes, initiate neutrophil chemotaxis to the alveolar space [16]. Neutrophils are the most abundant cell type of circulating leukocyte in human blood, and they migrate through chemotaxis to the site of infection, establishing the groundwork for an innate immune response [17,18]. At the site of infection, neutrophils carry out their antimicrobial activity through interrelated processes—phagocytosis, degranulation and the production of neutrophil extracellular traps (NETs) [19]. NETs are filamentous structures with scaffolds made from DNA, both nuclear and mitochondrial, and decorated by histones and cytoplasmatic proteins, such as neutrophil elastase, myeloperoxidase (MPO), cathepsin G, etc. [20]. NETs exhibit direct antimicrobial activity by bacterial trapping and immobilization, as well as due to antimicrobial peptides such as LL-37 [21,22].
The human cathelicidin, or LL-37, is an antimicrobial peptide. Strong activity against bacteria and viruses results from extracellular activation via serine protease 3 and kallikrein 5. By raising the permeability of bacterial cell walls, LL-37 shows direct bactericidal action; simultaneously, it shows indirect antibacterial effects by binding to and neutralizing bacterial endotoxins, including LPS [23]. Moreover, it can upset bacterial biofilms [22].
Considering all the above, the present study aimed to examine in vitro and ex vivo the role of neutrophils in L. pneumophila infection via NET release and further identification of proteins that may be implicated in the pathophysiology of LD infection.
2. Materials and Methods
2.1. Subjects
To carry out this study, 5 LD patients, as well as 5 (n = 5) healthy individuals (HI), or controls, were enrolled. Regarding the clinical manifestation, it is well recognized that the infection has two distinct forms: the non-pneumonic form, or Pontiac fever, and Legionnaires’ disease, a more severe form of infection which includes pneumonia. All our patients suffered from LD, and three of them had lobular pneumonia, without some extrapulmonary manifestations. Moreover, two patients’ clinical manifestations included extrapulmonary manifestations of LD, with diarrhea and abdominal pain, acute renal injury and elevation of cardiac enzymes (CPK). The study design complied with the Helsinki Declaration and had the approval of the Ethics Committee of the University General Hospital of Alexandroupolis, Greece (Ref. Number 32859/27 June 2025). Routine laboratory tests were performed on all subjects enrolled in this study (Table 1). For diagnosis of LD, a STANDARD F Legionella Ag FIA test system (Analyzer + Test device) for detecting Legionella pneumophila serogroups 1, 3, 5, 6 and 8 antigens via a urine sample was performed (SDBiosensor, Suwon-si, Republic οf Korea). The inclusion criteria for the LD group were a positive test for detecting L. pneumophila serogroups 1, 3, 5, 6 and 8 antigens via urine sample, and absence of other infectious etiology. The inclusion criterion for the HI group (controls) was absence of clinical manifestations of infection. Exclusion criteria included recent surgery, or hospital-acquired infection, except for LD.
2.2. Bacterial Strains
Legionella pneumophila subsp. pneumophila (ATCC 33152™) was used in this study. Initially, the microbial strain was cultivated onto buffered charcoal-yeast extract (BCYE) agar plates. Bacteria were collected and resuspended into sodium saline (0.9%NaCl) to adjust the turbidity of bacterial suspensions to 0.5 McFarland standards. Then, opsonization of bacteria was performed by incubating with serum from healthy donors for 30 min at 37 °C [24].
Bacterial strains, a clinical isolate of P. aeruginosa, were preserved in glycerol broth at −80 °C. Bacteria from an overnight culture on MacConkey agar were suspended in saline to an optical density of 0.5 McFarland, corresponding to a concentration of approximately 108 CFU/mL To specify the functionality (antimicrobial activity) of LL-37 peptides, bacterial strains of P. aeruginosa and L. pneumophila were cocultured with NETs structures, generated ex vivo from LD patients, or in vitro by clarithromycin [22,24].
2.3. Cell Types
2.3.1. Neutrophil
Venous blood from healthy individuals (HI) and LD patients were obtained in heparinized blood collection tubes (BD Vacutainer^®^ Heparin Tubes, Becton, Dickinson and Company, Franklin Lakes, NJ, USA). Then, neutrophils were isolated by performing Histopaque double-gradient density centrifugation (11,191 and 10,771, Sigma-Aldrich, St. Louis, MO, USA), as previously reported [25]. The purity of the isolated cells’ type (neutrophils) exceeded 98%, as observed by May–Grünwald–Giemsa staining. Neutrophils were seeded and cultured in 24-well plates at a density of 1–2 × 10^5^ cells per well in RPMI.
2.3.2. THP1 Cells
Cell culture
The human monocytic THP-1 cell line (ATCC TIB202; American Type Culture Collection, Rockville, MD, USA) was routinely cultured in a humidified CO_2_ incubator at 37 °C in RPMI 1640 medium, supplemented with 10% heat-inactivated fetal bovine serum, L-glutamine, 100 U/mL penicillin and 100 μg/mL streptomycin (Biosera, Boussens, France).
THP-1 differentiation
THP-1 cells were seeded at a density of 2.5 × 10^5^ cells/well in 24-well plates with sterile glass coverslips fully attached to the bottom of each well. Concomitantly, THP-1 monocytes were differentiated into macrophages with phorbol myristate acetate (PMA, Sigma-Aldrich, St. Louis, MO, USA) at a final concentration of 50 ng/mL at 37 °C with 5% CO_2_ for 48 h in a humidified atmosphere. After a 24 h resting period without PMA, THP-1 cells were treated with serum samples from HI and LD.
2.4. Stimulation and Inhibition Studies
2.4.1. Neutrophils’ Stimulation and NET Generation
Isolated neutrophils from LD patients, HI neutrophils, were incubated at 37 °C with 5% CO_2_ at a density of 1–2 × 105 cells per well in Roswell Park Memorial Institute 1640 medium (RPMI-1640, Thermo Fisher Scientific, Waltham, MA, USA), supplemented with 2% v/v FBS. Subsequently, they were incubated for 3 h at 37 °C with 5% CO_2_.
In addition, to investigate the role of neutrophils in the LD, neutrophils from healthy individuals (HI) were stimulated with: (a) 5% serum obtained from LD patients, (b) control serum (untreated condition), (c) Clarithromycin (Anfarm Hellas, Kifissia, Greece) was dissolved at a concentration of 50 mg/mL per stock, (d) opsonized L. pneumophila at a concentration of ~10 bacteria as previously reported and (e) serum from Familial Mediterranean Fever (FMF) patient as positive control for IL-1β decorated NETs [22,24,25].
2.4.2. Autophagy Study
For in vitro or ex vivo autophagy induction, samples were stimulated and incubated as described above and incubated for 90 min [22,25].
2.4.3. NET Structures Generation and Isolation
For in vitro or ex vivo NET structures generation, 2 × 10^6^ HI neutrophils were cultured in a 6-well cell culture plate (SPL Life Sciences, Kyonggido, Republic of Korea) in RPMI, supplemented with 2% v/v heterologous healthy donor serum, and stimulated with serum from LD patients or clarithromycin. Following a 3 h incubation (37 °C, 5% CO_2_), the cell culture medium was removed, cells were washed once with pre-warmed RPMI and fresh prewarmed RPMI was added. The cell culture plate was vigorously agitated, and NET structures were isolated. HI neutrophils that were not stimulated with patient serum served as control (control NETs) [22,25].
2.5. Immunofluorescence
2.5.1. Neutrophil Seeding
Peripheral blood neutrophils were seeded on poly-L-lysine-coated glass coverslips (Biocoat, New York, USA) in a 24-well cell culture plate (SPL Life Sciences, Kyonggi-do, Republic of Korea) and incubated for 3 h (37 °C, 5% CO_2_) to evaluate their NET release capacity and examine the NET protein profile. In addition to studying autophagy induction, samples were incubated for 90 min. Cells were fixed with 10% formaldehyde solution (Biognost, Zagreb, Croatia) for 15 min at room temperature. Nonspecific binding sites were blocked with 6% normal goat serum (Thermo Fisher Scientific, Waltham, MA, USA) in 1x PBS (blocking solution).
2.5.2. Antibodies
Following this, samples were stained with a primary antibody solution, consisting of a rabbit polyclonal anti-human neutrophil elastase (NE) poyclonal antibody (1:300 dilution, OriGene Technologies GmbH, Herford, Germany), a mouse anti-human IL-1β monoclonal antibody (mAb) (1:300 dilution, R&D System, Minneapolis, MN, USA) and a mouse anti-human LL-37 mAb (1:300 dilution, Santa Cruz Biotechnology, Dallas, TX, USA) in blocking solution, for 1 h at room temperature (RT). A polyclonal gout anti-rabbit IgG AlexaFluor 647 antibody (Invitrogen, Waltham, MA, USA) or a polyclonal rabbit anti-mouse IgG AlexaFluor 488 antibody (Invitrogen, Waltham, MA, USA). Finally, DAPI solution (Sigma-Aldrich, St Louis, MO, USA) was used for DNA counterstaining [22,25].
For protein detection by immunofluorescence, the following antibodies were used: mouse monoclonal antibody anti-LL-37 protein (1/300 dilution; Santa Cruz), mouse anti-IL1β and rabbit polyclonal antibody antι-Neutrophil Elastase (NE) (1/300 dilution; OriGene Technologies GmbH, Herford, Germany).
2.5.3. Autophagy
To study autophagy induction, samples were stained with anti-LC3b polyclonal antibody (1:100 dilution, OriGene Technologies GmbH, Herford, Germany), followed by a polyclonal anti-mouse IgG AlexaFluor488 antibody (Invitrogen, Waltham, MA, USA) as a secondary antibody. DNA was counterstained using DAPI [22]. In addition, to clarify, autophagy inhibition by Bafilomycin, and more specifically Bafilomycin A1 (BafA1), which inhibits the final stage of autophagy, was used.
2.5.4. Visualization
Imaging was performed on a customized Andor Revolution Spinning Disk Confocal system (Yokogawa CSU-X1; Yokogawa, Tokyo, Japan) built around an Olympus IX81 (Olympus Shinjuku, Tokyo, Japan) with 20× 0.45NA air lens, 40× 0.95NA air lens or 100× 1.40NA oil lens (UPlanXApo; Olympus Shinjuku, Tokyo, Japan) and a digital camera (Andor Zyla 4.2 sCMOS; Andor Technology Ltd., Belfast, Northern Ireland) (CIBIT-Bioimaging Facility, MBG-DUTH). The system was controlled by Andor IQ3.6.5 software (Andor Technology). Images were acquired as z-stacks with selected optical sections every 1 μm, through the entire cell volume.
2.5.5. Quantification
Quantification of NETs with PicoGreen dye was performed by fluorescence measurements on a multimode plate reader (EnSpire, PerkinElmer, Waltham, MA, USA) and presented as Relative fluorescence units (RFU).
2.6. RNA Extraction, cDNA Synthesis and qPCR
Neutrophils from LD patients, HI neutrophils stimulated with Lp serum, as well as with clarithromycin, were processed for RNA extraction, cDNA synthesis, and qPCR, as previously detailed [25,26]. To verify LL-37 expression in Lp-serum-treated neutrophils in vitro, expression of LL-37 (forward primer: 50TGGTGTCACTGCTACTG30, reverse primer: 50CATTGCGGTGGAGATTC30) was evaluated via qPCR. GAPDH (forward primer: 50GGGAAGCTTGTCATCAATGG30, reverse primer: 50CATCGCCCCACTTGATTTTG30) was utilized to normalize LL-37 expression, following the housekeeping gene method of normalization. qPCR was performed, and the 2-DDCt method was applied for data analysis [27].
2.7. Bead-Based Multiplex Immunoassay
The serum levels of interleukin (IL)-1β, IL-6, IL-10, IL-18, IL-23 and IL-33, were assessed using the LEGENDplexTM Multi-Analyte Flow Assay Kit (Biolegend, San Diego, CA, USA) [28,29]. The samples were prepared following the manufacturer’s instructions, and data acquisition was performed by the Attune NxT Flow Cytometer (Invitrogen by Thermo Fisher Scientific Inc.).
2.8. Statistical Analysis
For quantitative variables, age and interleukin levels, we calculated mean ± standard deviation (SD). Nonparametric tests (Mann–Whitney and Kruskal–Wallis) and the Chi-square (χ2) test were performed. The data for qualitative variables are presented as percentages. Statistical analysis was performed using IBM SPSS 26. Statistical significance was set to p < 0.05.
3. Results
3.1. Inflammatory Cytokines Are Detected in the Circulation of LD Patients
Considering our recently published data, which demonstrate that L. pneumophila-induced NETs express a major proinflammatory cytokine, IL-1β [24], that plays a cornerstone role in multi-organ failure during the LD, we determined cytokines in the sera. To assess the inflammatory milieu of L. pneumophila infection, serum samples from patients were analyzed using a bead-based multiplex immunoassay. We observed that the levels of proinflammatory cytokines IL-1β, IL-6, IL-18 and IL-33 were elevated in LD patients compared to HI in a statistically significant manner (Figure 1). On the other hand, there was no difference between the patients and the control group for TNF-α, IL-8, IL-12p70, IL-17A, 40.05 ± 18.8 vs. 37.6 ± 19.4, p = 0.9, 77.8 ± 5.6 vs. 60.3 ± 7.6 p = 0.21, 27.8 ± 6.1 vs. 14.9 ± 7.8 p = 0.2 and 7.2 ± 1.9 vs. 4.4 ± 2.5, p = 0.12, respectively.
3.2. Neutrophils Release NETs as a Response to L. pneumophila Ex Vivo and In Vitro
Considering the neutrophils’ ability to generate NETs as a mechanism to eliminate infection, we investigated whether activation of neutrophils during L. pneumophila infection releases NETs. We initially studied the ability of L. pneumophila to induce NETs formation ex vivo. NETs released by neutrophils from LD patients have been observed by immunofluorescence (Figure 2A–F), as well as the percentage of NETs formation (Figure 2G), and a fluorescence-based method using the PicoGreen assay (Figure 2H,I). Next, to support our ex vivo observations, neutrophils isolated from healthy individuals (HI) were stimulated with LD serum samples, representing the inflammatory disease microenvironment. This experiment highlights that the disease’s microenvironment can induce neutrophils to release NETs (Figure 2D). Furthermore, HI neutrophils were also pretreated with bacterial strains, as previously described [24]. We found that L. pneumophila pre-treated control neutrophils, as well as LD serum-treated cells, form NETs, as assessed via immunofluorescence microscopy. Several previous studies have demonstrated that potent inflammatory proteins expressed in NETs can determine the clinical manifestation of diseases [26,28,30,31]. In this context, to examine the proteinous components of NETs, and more specifically, the expression of IL-1β and correlation with disease severity, ex vivo and in vitro studies were performed (Figure 2D–F). In addition, HI neutrophils in vitro stimulated with FMF crisis serum were used as a positive control for IL-1β-bearing NETs (Figure 2C) [30]. We observe that the percentage of NETs released in LD patients with severe pneumonia and extrapulmonary manifestations was higher compared to patients with pneumonia and without extrapulmonary manifestations. Moreover, the IL-1β serum level was correlated with disease severity in a statistically significant manner 97.1 ± 24.8 vs. 32.1 ± 57.1, p = 0.003.
3.3. L. pneumophila-Induced NETs Are Unable to Express the Antimicrobial Peptide LL-37
Considering that LL-37 is one of the potent neutrophils’ antimicrobial peptides with direct bactericidal properties that may prevent the spread of infection [23], we examined whether these NETs are decorated with the antimicrobial peptide LL-37 and whether these NET structures can inhibit bacterial growth in vitro. First, to detect the expression of LL-37 on the NETs, both ex vivo and in vitro studies were performed. Untreated neutrophils were exemplified as a negative control, while clarithromycin was used as a control for LL-37-positive NETs (Figure 3A,B). Surprisingly, we observed that ex vivo and in vitro L. pneumophila-induced NETs do not decorate with LL-37 (Figure 3C,D). Moreover, producing LL-37 poor NETs during the LD was also detected by a decrease in the mRNA LL-37 level in neutrophils, activated in vitro with both LD serum and ex vivo LD (Figure 3Ε).
Next, given the bactericidal properties of NETs, we examined whether in these LD NETs, poor antimicrobial peptide LL-37 can inhibit bacterial growth in vitro. As a positive control in this experiment, clarithromycin-induced NETs were used. We found that only clarithromycin-induced NETs and LL-37-decorated NETs were able to reduce bactericidal growth in vitro. On the contrary, LD NETs were unable to restrain L. pneumophila growth in vitro (Figure 4).
3.4. L. pneumophila-Induced NETs Are Unable to Activate Autophagy
Previous reports demonstrated that NET release triggered by L. pneumophila is ROS-dependent [24]. Next, given the role of autophagy in NETs release, we examined whether L. pneumophila utilizes autophagy to induce NET generation ex vivo or in vitro. Moreover, we examined whether autophagy utilization takes place in different types of cells, macrophages and neutrophils. To verify this, macrophages were stimulated with NETs structures, as assessed by LC3b staining. Surprisingly, we observed that L. pneumophila does not utilize autophagy to induce NET generation ex vivo and in vitro. In parallel, we found that L. pneumophila-generated NETs structures activate autophagy machinery in vitro in macrophages (Figure 5).
3.5. Dual Role of Macrolides in Legionella pneumophila Infection
It is well recognized that quinolones and macrolides have excellent activity against Legionella spp. strains in vitro and reach adequate inhibitory concentration within phagocytes (both macrophages and neutrophils) [32,33]. Previously, in our study, we demonstrated the in vitro and in vivo ability of macrolide antibiotics to induce NET formation. Moreover, we show that clarithromycin-induced NETs are decorated by the antimicrobial peptide LL-37, which is in its active form and can act against multidrug-resistant A. baumannii in vitro [22]. In this study, we observed that ex vivo and in vitro L. pneumophila-induced NETs do not decorate with LL-37 (Figure 3C,D). In addition, we found that LD NETs are unable to restrain L. pneumophila growth in vitro (Figure 4). Taking into account the antimicrobial effect of clarithromycin-induced NETs, both clarithromycin-sensitive strains of L. pneumophila and resistance strains of P. aeruginosa were analyzed. We confirm that LL-37-decorated NETs were able to reduce bactericidal growth in vitro, both in clarithromycin-sensitive strains of L. pneumophila and in resistance strains of P. aeruginosa (Figure S1).
4. Discussion
This article demonstrates that L. pneumophila induces NETs formation in vitro and ex vivo; however, the ability to be externalized on NETs antimicrobial peptides like LL-37 and their functionality is still unknown. In our study, LD-driven NETs were shown to be unadorned LL-37. The ability of microorganisms to induce neutrophil extracellular traps, which lack antimicrobial activity, was previously reported by Jhelum et al. Authors observe that the abundance of the antimicrobial peptide cathelicidin (CAMP) was lower on NETs induced by PVL-positive S. aureus, and as such, they were unable to kill S. aureus [34]. These data were in line with our results. In addition, the treatment of HI neutrophils with clarithromycin increases the levels of LL-37 on NETs, leading to significant in vitro induction of antibacterial activity of LL-37-bearing NETs on L. pneumophila strain, as much as P. aeruginosa strain. Previous studies have reported that LL-37 function is not limited to antimicrobial activity. When bound to DNA in the NETs, LL-37 protects DNA from degradation. Using immunofluorescence microscopy, Neumann et al., demonstrate that LL-37 decorated NETs are distinctly more resistant to S. aureus nuclease degradation [35]. Moreover, in our previous study, we demonstrated that upregulation of LL-37 on NETs improves the wound-healing capacity of fibroblasts [22,36]. Furthermore, its lipopolysaccharide (LPS)-neutralizing effect was reported [37].
Since neutrophils have been shown to express IL-1β under L. pneumophila. activation in vitro [24], we investigated its presence on L. pneumophila ex vivo-induced NETs. We demonstrate that during the LD, neutrophils produce NETs, which express a major proinflammatory cytokine IL-1β. It was shown that IL-1β expression depends on clinical manifestation. Moreover, it was shown that other cytokines of the IL-1 cytokine superfamily, IL-1β, IL-18 and IL-33, were also elevated. Similarly, a proinflammatory cytokine IL-6 was elevated in LD patients compared to HI, in a statistically significant manner. Our results are in line with previously reported data that show IL-18 or IL-33 elevation in Legionella pneumophila infection [38]. Rakebrandt et al. report a protective effect in subsequent heterologous challenges with Legionella pneumophila [38].
Taking together the data presented in this article, we proposed the pathophysiological mechanisms of LD (Figure 6).
The bacteria, after the entry to the host cell, are within a vacuole (Legionella-containing vacuole—LCV). Afterwards, the intracellular replication of Legionella spp. is advanced. Furthermore, L. pneumophila binds via Mip protein to the collagen, which allows bacterial transmigration to the blood. In addition, L. pneumophila possesses autophagy inhibitory mechanisms in neutrophils. Further to this, L. pneumophila induces NETs formation, which are decorated with proinflammatory cytokines IL-1β that play a critical role in multiorgan damage in LD. On the other hand, the abundance of the antimicrobial peptide cathelicidin (CAMP), or LL-37, was lower on L. pneumophila-induced NETs. Finally, Clarithromycin-induced NETs bear LL-37. In this way, clarithromycin plays a dual role in Legionella infection.
Our study has limitations that we would like to mention. Initially, we had a small group of patients with LD. Consequently, we could not perform analysis with different disease phenotypes. Furthermore, we were unable to assess the quantitative determination of LL-37 in serum. Lastly, taking into account the role of IL-1β in inflammation and its elevated level in LD, we did not perform experiments with IL-1β inhibition. Hence, the next essential goal is to analyze our results in a large group of patients with LD, as well as the role of IL-1β inhibition by Kineret in the pathogenesis of multiorgan failure in LD.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Jackson E. Crocker T. Smadel J. Studies on Two Rickettsia-like Agents Probably Isolated from Guinea Pigs Bacteriol. Proc.195252119
- 2Mc Dade J.E. Shepard C.C. Fraser D.W. Tsai T.R. Redus M.A. Dowdle W.R. Legionnaires’ Disease: Isolation of a Bacterium and Demonstration of Its Role in Other Respiratory Disease N. Engl. J. Med.19772971197120310.1056/NEJM 197712012972202335245 · doi ↗ · pubmed ↗
- 3Del Piano M. La Palombara P. Nicosia R. Picchiotti R. The Legionellosis Boll. Dell’istituto Sieroter. Milan.19846387996380526 · pubmed ↗
- 4Sanford J.P. Legionnaires’ Disease—The First Thousand Days N. Engl. J. Med.197930065465610.1056/NEJM 197903223001205763287 · doi ↗ · pubmed ↗
- 5Viasus D. Gaia V. Manzur-Barbur C. CarratalàJ. Legionnaires’ Disease: Update on Diagnosis and Treatment Infect. Dis. Ther.20221197398610.1007/s 40121-022-00635-735505000 PMC 9124264 · doi ↗ · pubmed ↗
- 6Yu V.L. Plouffe J.F. Pastoris M.C. Stout J.E. Schousboe M. Widmer A. Summersgill J. File T. Heath C.M. Paterson D.L. Distribution of Legionella Species and Serogroups Isolated by Culture in Patients with Sporadic Community-Acquired Legionellosis: An International Collaborative Survey J. Infect. Dis.200218612712810.1086/34108712089674 · doi ↗ · pubmed ↗
- 7Brady M.F. Sundareshan V. Legionnaires’ Disease Stat Pearls Stat Pearls Publishing Treasure Island, FL, USA 2022
- 8Alexandropoulou I.G. Ntougias S. Konstantinidis T.G. Parasidis T.A. Panopoulou M. Constantinidis T.C. Environmental Surveillance and Molecular Epidemiology of Waterborne Pathogen Legionella Pneumophila in Health-Care Facilities of Northeastern Greece: A 4-Year Survey Environ. Sci. Pollut. Res. Int.2015227628764010.1007/s 11356-014-3740-825712880 · doi ↗ · pubmed ↗