Crithidia fasciculata Shows Non-Pathogenic Behavior in Leishmania Co-Infection Related to Temperature Stress, In Vitro and In Vivo Infections, and Amphotericin B Susceptibility
Julia Fernandes Barbosa dos Santos, Carolina Boucinha Martins, Valter Viana Andrade-Neto, Thais Lemos-Silva, Rosiane Freire dos Santos, Silvia Amaral Gonçalves da-Silva, Yara Maria Traub-Csekö, Rubem Figueiredo Sadok Menna-Barreto, Eduardo Caio Torres-Santos

TL;DR
This study shows that Crithidia fasciculata does not behave as a pathogen in leishmaniasis co-infections, both in lab and animal models.
Contribution
The study provides new insights into the non-pathogenic nature of C. fasciculata in co-infections with Leishmania under various temperature and infection conditions.
Findings
Crithidia fasciculata strains showed morphological changes and reduced survival at elevated temperatures.
Crithidia failed to infect macrophages and was not detected in mouse or hamster tissues.
Crithidia established infection in sandflies but did not cause disease in co-infection models.
Abstract
There is increasing evidence on the occurrence of Crithidia spp. in patients presenting either cutaneous or visceral leishmaniasis, solely or associated with Leishmania. We analyzed growth, morphology, and temperature tolerance of two C. fasciculata strains, the reference strain COLPROT048 and patient isolate COLPROT606. We also evaluated their co-cultivation with L. braziliensis, macrophage infectivity, and infections in hamsters, BALB/c mice, and sandflies. In culture, both Crithidia strains survived at 32 °C for 96 h, showing major morphological alterations and decreased mitochondrial membrane potential, with ΔΨm reducing to 52% in COLPROT606. At 34 °C, the patient isolate showed an 80% reduction in cell number. Mixed cultivation of Crithidia-Leishmania led to recovery of only Crithidia. In macrophages, C. fasciculata alone was virtually eliminated, and in co-infection only…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
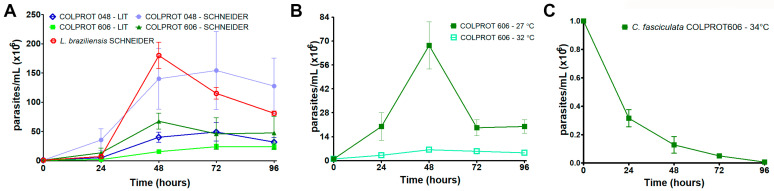 Figure 1
Figure 1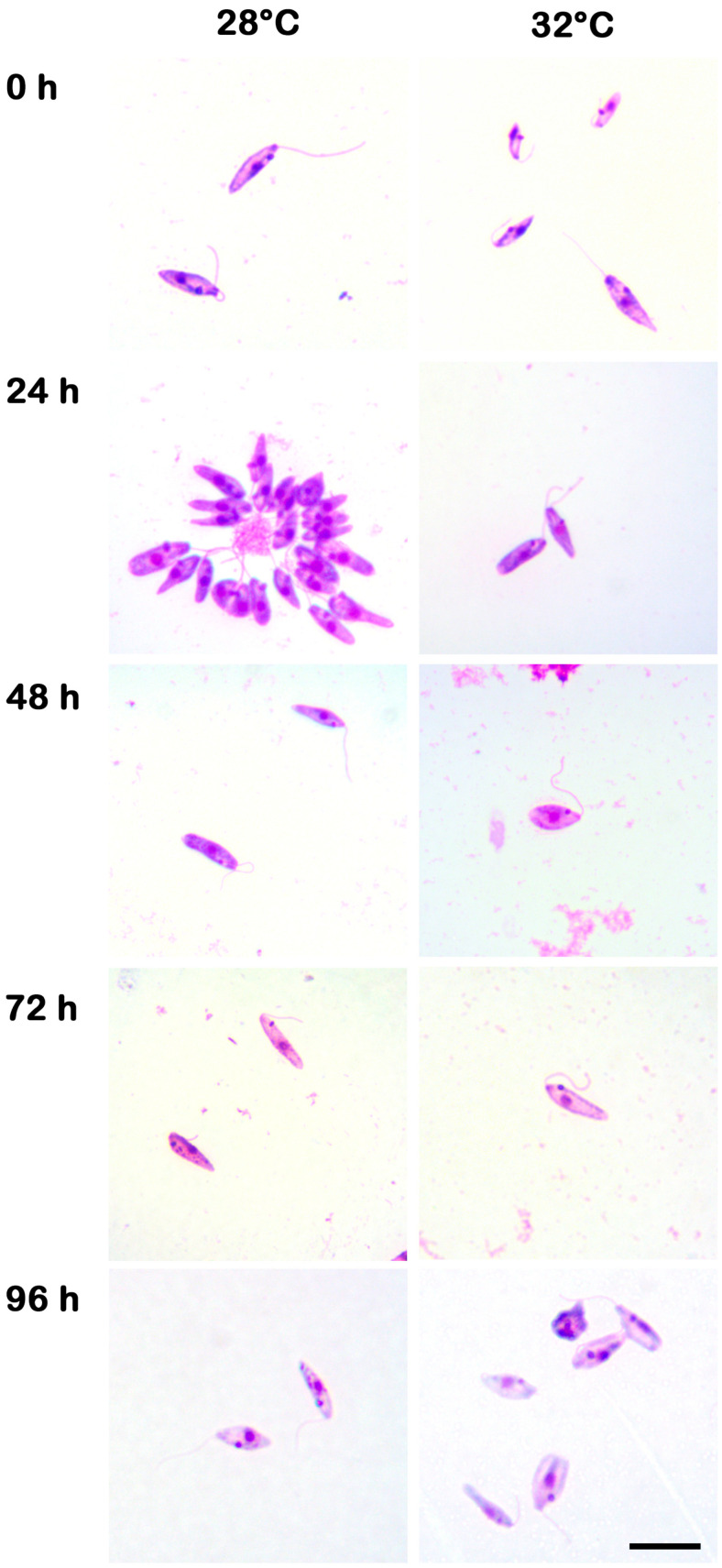 Figure 2
Figure 2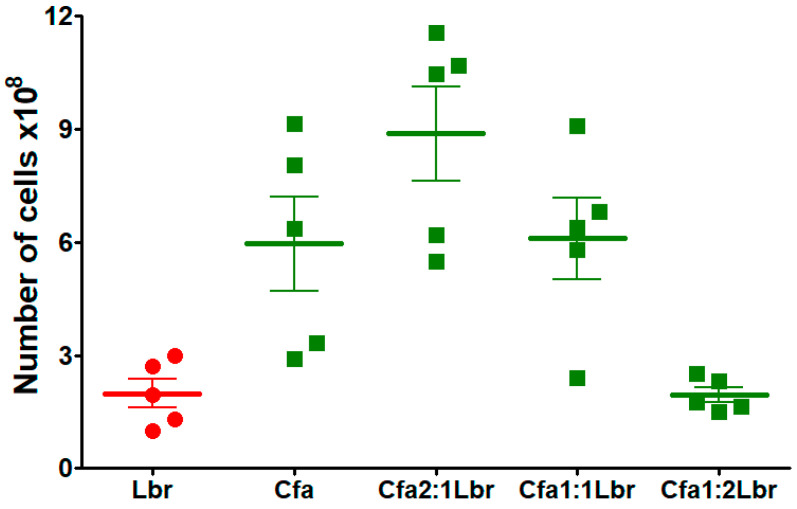 Figure 3
Figure 3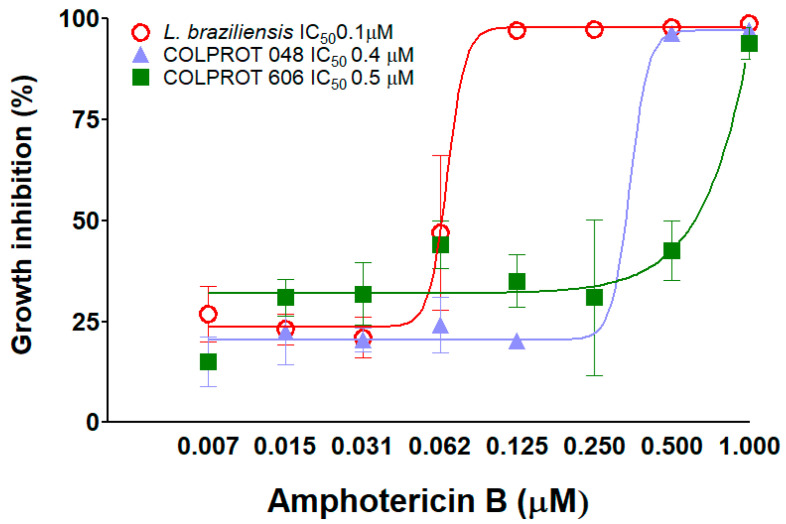 Figure 4
Figure 4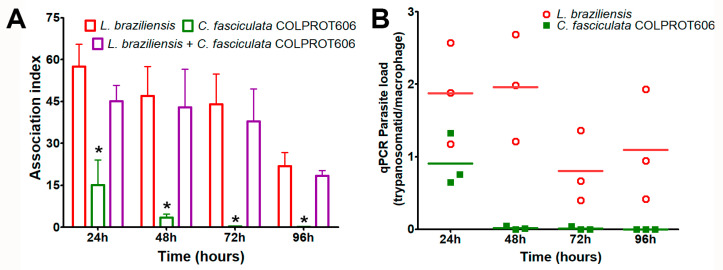 Figure 5
Figure 5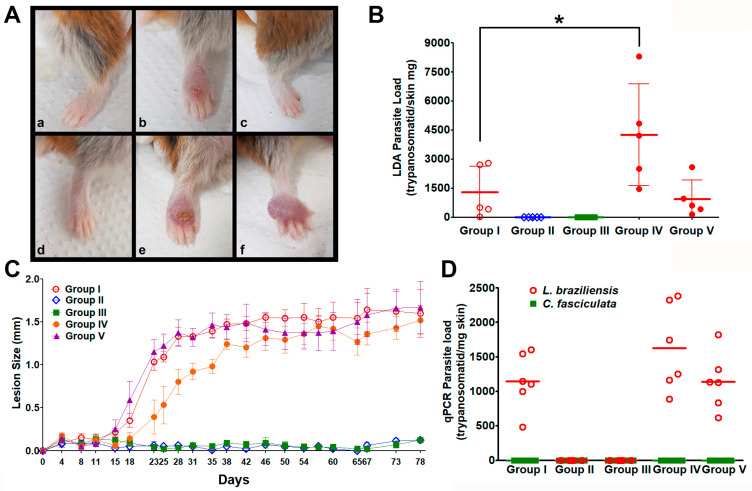 Figure 6
Figure 6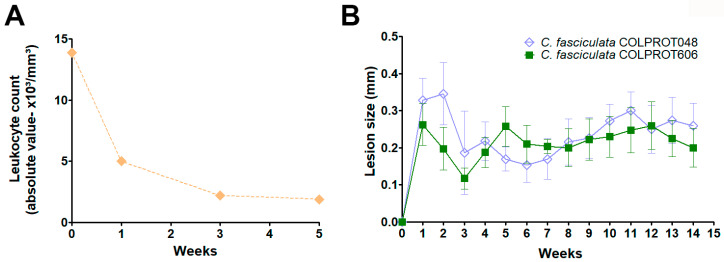 Figure 7
Figure 7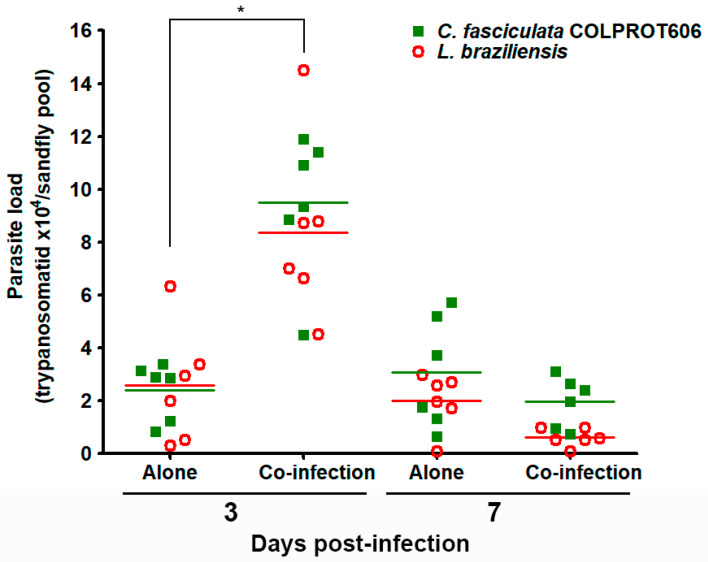 Figure 8
Figure 8- —Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ)
- —Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)
- —Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
- —Instituto Oswaldo Cruz (IOC)
- —Fundação Oswaldo Cruz (FIOCRUZ)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsResearch on Leishmaniasis Studies · Trypanosoma species research and implications · Helminth infection and control
1. Introduction
Trypanosomatid protozoa constitute a diverse group that includes the etiological agents of several neglected tropical diseases (NTDs) [1]. These parasites are transmitted by a variety of arthropod vectors and cause a wide spectrum of diseases associated with substantial morbidity and mortality [2]. Conventionally, trypanosomatid species are classified into dixenous parasites, which complete their life cycle, alternating between invertebrate and vertebrate or plant hosts; and monoxenous trypanosomatids, assumed to be restricted to invertebrate hosts, mainly insects. While dixenous species have received more attention due to their considerable medical, veterinary, and economic relevance [3], typically nonpathogenic monoxenous trypanosomatids have gained increasing interest due to growing reports of their occurrence in mammalian hosts, including humans [4,5,6,7,8,9,10,11,12,13,14,15,16].
Most of the recent reports of monoxenous infections in mammals have emerged from the atypical nature of some cases, prompting further investigations into the pathogenic species involved. In most instances, these infections occur in co-infection with HIV or other trypanosomatids, such as Leishmania [7]. These findings disclose ongoing controversies surrounding their taxonomy and potential pathogenicity in vertebrate hosts. Nevertheless, the impact of monoxenous trypanosomatid in the pathogenesis of leishmaniasis remains poorly understood, raising critical questions about how mixed infections affect disease severity, diagnosis, and treatment.
In this sense, our research group received an isolate from a cutaneous lesion of an immunocompetent man in Cusco, Peru [4]. The parasite was initially identified as Leishmania braziliensis based on clinical presentation and geographic distribution, but further studies revealed atypical features, such as non-fastidious cultivation. However, more accurate genomic analysis employing the molecular markers gGAPDH and V7V8 ribosomal RNA region identified the parasite as Crithidia fasciculata, which was subsequently deposited in the Protozoa Collection in the Oswaldo Cruz Foundation (COLPROT-Fiocruz) [14].
The isolation of C. fasciculata from a human lesion led us to hypothesize that the monoxenous parasite may persist in mammalian hosts and modulate the outcomes of co-infection with L. braziliensis. To elucidate this issue, a comprehensive analysis of the growth dynamics and infectivity of C. fasciculata isolated from a human patient were performed. The parasite’s behavior in different culture media was assessed, as well as its in vitro and in vivo infectivity against mouse and hamster models. Finally, we examined the potential effect of the co-infection of C. fasciculata and L. braziliensis on the colonization of the sandfly vector Lutzomyia longipalpis, thereby contributing to a better understanding of its potential as a vector-borne pathogen. This study provides valuable information into the ecological and pathological roles of monoxenous trypanosomatids in human infections and advances our knowledge of co-infection mechanisms and the pathogenic potential of C. fasciculata.
2. Materials and Methods
2.1. Parasite Cultivation
The following strains from our Protozoa Collection (COLPROT—Fiocruz) were used: (i) the type strain of C. fasciculata (COLPROT048), isolated from Anopheles quadrimaculatus in 1926; (ii) C. fasciculata (COLPROT606), isolated from a human patient in 1994; and (iii) L. braziliensis (ThorMCAN/BR/97/P142) obtained from LBTq/IOC and maintained through passages in BALB/c mice. C. fasciculata strains were initially cultivated in NNN’/LIT medium with 10% inactivated FBS, while L. braziliensis was maintained in Schneider’s medium (Sigma-Aldrich, St. Louis, MO, USA) with 20% inactivated FBS and 2% sterile human filtered urine at 27 °C. C. fasciculata strains were subsequently maintained in Schneider’s medium with 10% inactivated FBS (Gibco) at 27 °C. Passages were performed twice a week until the fifth passage.
2.2. Growth Curves
Growth curves were established by inoculating C. fasciculata (COLPROT606 and COLPROT048) and L. braziliensis (MCAN/BR/97/P142) cultures, starting with 1.0 × 10^6^ parasites/mL in LIT and Schneider’s medium supplemented with 10% and 20% inactivated FBS, respectively. Parasites were maintained in triplicate and monitored every 24 h during five days. Parasite viability was assessed using a Neubauer chamber and Trypan blue (Sigma-Aldrich) exclusion. The effect of temperature on COLPROT606 was evaluated at 27 °C, 32 °C and 34 °C. Replicates were passed during the logarithmic growth phase [17].
Alternatively, to evaluate mixed cultures, L. braziliensis and COLPROT606 were co-cultured in Schneider’s medium with 20% inactivated FBS and 2% sterile male human urine at 27 °C in the following ratios: L. braziliensis (control), COLPROT606 (control), and three mixed ratios (1:1, 2:1, 1:2). In all mixed cultures, the total number of parasites was equivalent to the corresponding single-species control, with the number of each species adjusted according to the ratio, e.g., 1 × 10^6^ each in 1:1; 1.33 × 10^6^ of one species and 0.67 × 10^6^ of the other in 2:1 or 1:2, totaling 2 × 10^6^ parasites. Cultures were maintained in 5 mL medium, monitored every 3 days for contamination and growth, and 300 µL culture was transferred into fresh medium. After five passages, the cultures were washed three times with sterile PBS, centrifuged, and stored in TRIzol^®^ (Invitrogen, Waltham, MA, USA) at −80 °C for RNA extraction and qPCR analysis [18].
2.3. Morphological Analysis
For cell morphology analysis, C. fasciculata COLPROT606 and COLPROT048 were stained with Giemsa (Sigma-Aldrich) and observed under a Zeiss AxioObserver M1 optical microscope with a 100 × objective. Smears were prepared from log phase samples (5.0 × 10^6^ parasites/mL), fixed in 100% methanol for 10 min, air-dried, treated with 5N HCl for 10 min, washed, and stained with Giemsa for 1 h.
2.4. Mitochondrial Membrane Potential (ΔΨm) Assay
To evaluate the impact of temperature on C. fasciculata strains, the mitochondrial membrane potential (ΔΨm) of parasites incubated at 32 °C was evaluated by flow cytometry. The parasites were incubated with 50 nM tetramethylrhodamine (TMRE) (Molecular Probes) for 15 min at 28 °C, using 50 μM carbonyl cyanide m-chlorophenyl hydrazone (CCCP) (Sigma, St. Louis, MO, USA) as a control for ΔΨm dissipation. Variations in TMRE fluorescence were quantified using an index of variation (IV) following the equation IV = (ME − MC)/MC, where ME corresponds to the median fluorescence of parasites under experimental conditions of 32 °C and MC corresponds to the median fluorescence of parasites under control conditions at 27 °C. Negative IV values indicate depolarization of the mitochondrial membrane. Fluorescence values of CCCP-treated parasites were subtracted from both ME and MC values [19]. In parallel, the parasites were labeled with 0.1 µM TO-PRO3 iodide (Invitrogen) for 30 min to evaluate the plasma membrane integrity. Parasites incubated in 0.01% Triton X-100 (Sigma) were used as positive control. The parasites (10,000) were kept on ice until the acquisition on Beckman Coulter’s CyAn Flow Cytometer, and finally analyzed in the CytExpert 2.5 software.
2.5. Murine Peritoneal Macrophages In Vitro Infection
Peritoneal macrophages were extracted from BALB/c mice and adhered to coverslips (3 × 10^5^ cells) with RPMI 1640 medium (Gibco, Waltham, MA, USA) in a 4% CO_2_ atmosphere at 37 °C for 24 h, and were then washed thrice with PBS. Subsequently, macrophages (1.0 × 10^6^ cells/well) were infected with L. braziliensis (MCAN/BR/97/P142 strain) and both C. fasciculata strains (COLPROT048 and COLPROT606) at a 5 parasites per macrophage ratio. For co-infections, a 1:1 ratio was employed, yielding 2.5 parasites of each species per macrophage, thereby maintaining the total ratio of 5 parasites per macrophage used in monoinfections. Cells were cultured in RPMI medium supplemented with 10% FBS, 1% glutamine, and 1% pyruvate for 24, 48, 72, and 96 h at 35 °C with 5% CO_2_. After incubation, slides were stained with Panotic Fast (Laborclin, Pinhais, Brazil), and the infection index was determined by counting under an optical microscope using the formula: (% infected macrophages × amastigotes count)/total macrophages [20].
2.6. Amphotericin B Sensitivity Assays
For Amphotericin B assays, 4 × 10^6^ parasites/mL were used in 96-well plates. The drug was tested in a serial dilution range starting at 1 μM. After 72 h of incubation at 26 °C, cell viability was assessed by fluorometry using 50 µM Alamar Blue^®^resazurin. Readings were taken with a SpectraMax GEMINI XPS (Molecular Devices, San Jose, CA, USA), with an excitation of 560 nm to an emission detection at 590 nm. All assays were performed in triplicate. The results were plotted as the 50% inhibitory concentration (IC_50_/72 h) for parasite growth, using a nonlinear regression analysis on a semi-logarithmic scale obtained from GraphPad Prism 5.0 software [21].
2.7. In Vivo Experimental Infection Models
In vivo experiments were conducted in two models. Female BALB/c mice (4–8 weeks old, n = per group) from ICTB Fiocruz, housed at the Oswaldo Cruz Institute (Animal Ethics Committee approval: L002/2022), were infected in the right ear with 2.0 × 10^6^ parasites in the following groups: Group I—L. braziliensis (control), Group II –C. fasciculata COLPROT048, Group III—C. fasciculata COLPROT606, Group IV—L. braziliensis + COLPROT 048 (1:1 mixture, 1.0 × 10^6^ of each), and Group V—L. braziliensis + COLPROT606 (1:1 mixture, 1.0 × 10^6^ of each). Lesion size was monitored twice weekly for 35 days with a dial caliper (Mitutoyo, Kawasaki-shi, Japan). The animals had the lesion size measured for 78 days post-infection, and were euthanized at the end of the experiment. The infected paw and popliteal lymph nodes were removed and macerated for analysis by limiting dilution (LDA) and qPCR absolute quantification. Parasitic load was evaluated by limiting dilution and RT-qPCR. The second in vivo model was Golden hamsters (6–8 weeks old) from the UERJ animal facility (Animal Ethics Committee approval: 023/2022), which were used in similar experiments, i.e., infected in the right paw with 1.0 × 10^6^ parasites and monitored for lesion size. After euthanasia, the infected paw and popliteal lymph nodes were removed for analysis [21].
For the immunosuppression assays, BALB/c mice (n = 10 per group) were treated with cyclophosphamide (150 mg/kg) intraperitoneally, and subsequently infected with 1 × 10^7^ C. fasciculata COLPROT048 or COLPROT606. Lesions were monitored weekly, and parasitic load was assessed by limiting dilution.
2.8. RT-qPCR Quantification
Parasitic load was quantified using reverse transcriptase quantitative PCR (RT-qPCR). Primers and probes (IDT Inc., Newark, NJ, USA) were designed based on conserved regions of the SSU rRNA gene sequences of L. braziliensis and C. fasciculata using MUSCLE and Primer Express software v3.0.1. RNA was extracted with TRIzol and RNeasy Mini Kit (Qiagen, Hilden, Germany), treated with DNase (Sigma), and converted to cDNA using the SuperScript IV VILO kit (Invitrogen). The absence of genomic DNA amplification was confirmed by conventional PCR with actin primers in the no-reverse transcriptase control provided by the kit [17]. qPCR was performed on an ABI Prism 7500 Fast system, with the primers and probes described in Table 1. Standard curves were generated from serial dilutions of cDNA (10^7^ to 10^2^ parasites equivalents) to quantify parasitemia and assess primer efficiency. All the reactions were performed in triplicate.
2.9. Sandflies Experimental Infection
Infections of sandflies were performed using 5–8-day-old female L. longipalpis by artificially feeding on mice blood containing 5.0 × 10^6^ parasites/mL of L. braziliensis (MCAN/BR/97/P142), C. fasciculata (COLPROT606), or a 1:1 mixture of both parasites (2.5 × 10^6^ of each species). Pools of 10 sandflies from 3- and 7 days post-infection had their RNA extracted and cDNA synthesized by the protocols described previously. The parasite load was evaluated by qPCR using SYBR Green (Invitrogen) and employing the same SSU rRNA primers for the trypanosomatids’ detection and L. longipalpis ribosomal protein (RP49) as a normalization control [22]. The parasite load was represented by the number of quantified parasites (L. braziliensis or C. fasciculata) × 10^4^ per the number of sandflies in the pools (trypanosomatid × 10^4^/sandfly pool).
2.10. Statistical Analysis
All data were analyzed using GraphPad Prism 5.0 software (San Diego, CA, USA). For growth curves, viability assays, and drug sensitivity tests, normally distributed data with equal variances were compared using two-way ANOVA followed by Bonferroni’s multiple-comparison post hoc test. For macrophage infection indices and parasite load in animal tissues, comparisons between two groups were performed using Student’s t-test when data were normally distributed, or the Mann–Whitney U test otherwise. For sand fly infection and colonization experiments, parasite load was analyzed by two-way ANOVA with Bonferroni’s correction. Values of p < 0.05 were considered statistically significant.
3. Results
3.1. Growth Kinetics of C. fasciculata COLPROT60
The presence of monoxenous co-infections with Leishmania should imply several adaptations to the environment of the human body, such as temperature. Pathogenic Leishmania species exhibit intracellular amastigotes in mammalian cells and extracellular promastigotes in the vector, while Crithidia species typically display the choanomastigote form, but promastigotes and intermediate forms may appear in cultures. To investigate the growth kinetic of the isolated C. fasciculata COLPROT606, parasites were cultured in LIT medium with 10% FBS and Schneider’s medium with 20% FBS and 2% sterile human urine at 27 °C (Figure 1A). Growth analysis revealed faster growth of the reference strain (COLPROT048) in Schneider’s medium compared to LIT, and a slight increase in cell number for C. fasciculata COLPROT606 in Schneider’s medium. L. braziliensis failed to grow in LIT medium, demanding the use of Schneider’s medium for subsequent experiments. Additionally, C. fasciculata COLPROT606 survived and replicated at 32 °C, although in fewer numbers than at 27 °C (Figure 1B). However, the parasites were not capable of surviving at 34 °C, resulting in an 80% cell death within 24 h (Figure 1C). Similar results were observed using C. fasciculata COLPROT048. Additionally, 32 °C is the temperature used to induce Leishmania amastigogenesis, and so it may mimic the temperature within mammalian host cells.
Morphological parameters at different temperatures of the growth kinetics were assessed using Giemsa staining prepared every 24 h (Figure 2). Cells with subtle morphological differences were observed at 32 °C, whereas at 27 °C they maintained a less altered morphology with rosette formation, which indicates high cellular growth and adaptation. To assess C. fasciculata survival at elevated temperatures, mitochondrial membrane potential (ΔΨm) was measured, showing that both strains maintained mitochondrial function more effectively than L. braziliensis, indicating moderate thermotolerance (Table S1).
3.2. Cell Growth Evaluation in Leishmania-Crithidia Co-Cultures
A common approach to isolate parasites from clinical lesions requires the inoculation of the biopsy material in culture for taxonomic identification following cell growth. However, this methodology raises the concern of whether one parasite might overcome the other in the culture. To explore this question, we simulated an in vitro mixture under the following conditions: L. braziliensis alone (control), C. fasciculata COLPROT606 alone (control), a 1:1 mixture of L. braziliensis and C. fasciculata, and a 2:1 mixture of the two parasites The cultures were subcultured every 3 days, and after 5 passages the cells had their RNA extracted for quantitative RT-qPCR analysis.
To quantify the number of each trypanosomatid in the co-infection’s assays, a sensitive and reproducible qPCR was developed. We standardized parasite load quantification by RT-qPCR employing the small subunit of the ribosomal RNA (SSU rRNA) primers and probes, which provides a higher specificity and sensibility to quantify each trypanosomatid without unspecific amplification (Figure S1). In the mixed cultures, the results demonstrated that C. fasciculata successfully outgrew L. braziliensis in all mixed infection conditions (Figure 3). The absolute quantification did not reveal the presence of L. braziliensis in any co-infection condition.
3.3. Susceptibility to Amphotericin B
Due to the increasing presence of non-pathogenic species in vertebrate hosts, which may exacerbate symptoms or promote resistance to leishmaniasis treatment [5,6], we evaluated C. fasciculata susceptibility to amphotericin B, a drug commonly used for leishmaniasis treatment. L. braziliensis showed an IC_50_/72 h of 0.1 ± 0.02 μM, while the values were 0.4 ± 0.02 and μM for and 0.5 ± 0.01 μM for COLPROT048 and COLPROT606, respectively (Figure 4).
3.4. In Vitro Coinfection of C. fasciculata and L. braziliensis
Macrophages are key immune cells controlling Leishmania infections, and the failure to control parasite replication within these cells is critical for disease progression. Here, we analyzed the ability of C. fasciculata parasites to infect macrophages, and maintain the infection for 96 h, by optical microscopy. As expected, L. braziliensis was internalized by macrophages and maintained infection for 4 days (Figure 5). In contrast, C. fasciculata strains (COLPROT048 and COLPROT606) showed a significantly lower number of parasites inside macrophages at 24 h post infection. This number persisted low at 48 and 72 h, decreasing even more at 96 h (Figure 5A). No statistical difference was observed between C. fasciculata COLPROT048 and COLPROT606 strains.
Considering that L. braziliensis might be able to modulate the immune response to support the survival of C. fasciculata, in vitro coinfection of murine macrophages was also evaluated by RT-qPCR. As observed previously, L. braziliensis alone was successfully internalized and maintained the macrophage for 96 h, but C. fasciculata COLPROT606 failed to persist alone in macrophages over the same period. In coinfected murine macrophages, 5 parasites of each species were added per macrophage, keeping the total number equal to that of the single-species control. L. braziliensis RNA was detected in high number at the 96 h of infection, but C. fasciculata was only detected at 24 h pos-infection (Figure 5B). These findings suggest that L. braziliensis does not enhance the survival of C. fasciculata within macrophages or exacerbate infection, but rather eliminated C. fasciculata from the vertebrate host cells.
3.5. In Vivo Experimental Infections
The infectivity of C. fasciculata was evaluated in both BALB/c mice and golden hamsters, two established experimental models for Leishmania spp. infection. The golden hamsters are the most susceptible in vivo experimental model for L. braziliensis infection, capable of developing large and self-resolving lesions [23], Therefore, female hamsters were infected in the dorsum of their right hind paw with 1.0 × 10^6^ parasites according to the following protocol: Group I—L. braziliensis alone, Group II—C. fasciculata COLPROT048, Group III—C. fasciculata COLPROT606, Group IV—L. braziliensis co-infected with C. fasciculata COLPROT048, and Group V—L. braziliensis co-infected with C. fasciculata COLPROT606 (Figure 6).
Our results reported apparent lesions in control Group I, and in the co-infections of L. braziliensis with C. fasciculata COLPROT048 and COLPROT606, Groups IV and V, including the typical ulceration of this lesion in this experimental model (Figure 6A,C). However, no apparent lesions were observed on the paws of the hamsters infected only with C. fasciculata, Groups II and III. Moreover, the LDA parasite burden analysis did not report any parasite growth of in C. fasciculata strains infected alone, Groups II and III (Figure 6B). In the systems in which it was possible to observe a LDA positive parasite growth, Groups I, IV and V, the paw samples had a higher burden compared to popliteal lymph node.
In order to improve our co-infection analysis, we performed a qPCR quantification of the removed paws to detect L. braziliensis and C. fasciculata RNA, which could indicate the presence of live parasites at the end of the experiment. No C. fasciculata RNA was detected by RT-qPCR in the co-infection’s groups after 78 days of infection, or Crithidia alone (Figure 6D). However, L. braziliensis was detected in all the three groups where this parasite was used. A mean of 1145 parasites/mg skin equivalents were observed in Group I, 1625 parasites/mg in the co-infected Group IV, and 1141 parasites/mg in the co-infected Group V (Figure 6D). The changes in the qPCR parasite load were not statistically significant, suggesting that the co-infected were not capable of improving the hamster infection. The absence of C. fasciculata RNA in Groups II and III leads us to assume that this parasite is not able to maintain itself in hamsters.
In the BALB/c mouse model, infection with L. braziliensis and C. fasciculata COLPROT048 and COLPROT606 induced significant inflammation, as measured by increased ear thickness (Figure S2A). However, in hamsters C. fasciculata COLPROT048 and COLPROT606 alone failed to induce any visible signs of infection or growth, indicating that C. fasciculata are also not capable of surviving or establishing an infection in BALB/c mice (Figure S2B). In addition, no C. fasciculata growth or RNA were observed. Notwithstanding, we decided to investigate the persistence of C. fasciculata in immunosuppressed BALB/c mice treated weekly with cyclophosphamide (3 mg/animal, intraperitoneally). The animals were infected in the left hind paw with C. fasciculata COLPROT048 and with the clinical isolate C. fasciculata COLPROT606. A control group received the same treatment without infection. Immunosuppression efficacy was confirmed by a decrease in leukocyte count from 13.9 to 1.9 over 5 weeks (Figure 7A). The infection was monitored by measuring paw thickness, and after 14 weeks, both the paws and draining popliteal lymph nodes were collected for LDA parasite burden analysis. An initial increase in paw thickness was observed, followed by a stabilization in the size of the lesion (Figure 7B), but no parasite growth was detected in cultures from the paws or lymph nodes of cyclophosphamide-treated mice. These results suggest that C. fasciculata is unable to persist in BALB/c mice, even under immunosuppression.
3.6. Co-Infection of Sandflies with L. braziliensis and C. fasciculata COLPROT606
To assess the transmissibility of C. fasciculata in an invertebrate host, we tested its ability to colonize the L. longipalpis sandfly, a known permissive vector of Leishmania spp. [24]. Female sandflies (100 per group) were fed with blood containing: (i) L. braziliensis, (ii) C. fasciculata (COLPROT606), or (iii) both parasites (1:1). A pool of 10 insects was collected on days 3 and 7 post-infection and analyzed by RT-qPCR. Results showed that C. fasciculata and L. braziliensis were present at the two time points, both alone and in coinfection, (Figure 8). These findings suggest that C. fasciculata can colonize L. longipalpis, offering insights into the transmission dynamics of these trypanosomatids in vertebrate hosts. Moreover, the co-infection significantly increased the number of both parasites on the third day, rising from 2.4 × 10^4^ to 9.5 × 10^4^ C. fasciculata/sandfly and 2.5 × 10^4^ to 8.3 × 10^4^ L. braziliensis/sandfly. On day 7, the number of both parasites decreased and no statistical difference was observed both alone and in the mixed infection (Figure 8).
4. Discussion
Recent reports of human infections by monoxenous trypanosomatids, previously assumed to be insect-exclusive, are raising concerns about their pathogenic potential in vertebrates, including humans. Leptomonas seymouri has been frequently identified in co-infections with Leishmania donovani in the Indian subcontinent, often associated with more severe clinical outcomes and possible drug resistance [15,16]. Similarly, Crithidia spp. have been reported in both humans and animals, with cases documented in Brazil and Iran [4,14]. In humans, monoxenous infections are primarily described in immunocompromised individuals co-infected with HIV and L. major or L. infantum, but have also been detected in immunocompetent hosts as the sole pathogen [4,5,6,7,8,9,10,11,12,13,14]. Additionally, species of Herpetomonas and Blechomonas (formerly classified as Leptomonas) have been sporadically detected in humans [7]. These emerging reports suggest an expanding host range and increased adaptability of monoxenous trypanosomatids, reinforcing the need for intensified surveillance and continued investigation into their pathogenic mechanisms. Therefore, in this study, we further investigated the biological and pathogenic characteristics of a C. fasciculata isolate obtained from a skin lesion of a patient from Cusco, Peru [4,17], to better understand its behavior under experimental conditions and its potential role in human infections.
Our findings demonstrate that C. fasciculata was capable of surviving and proliferating in two media specifically designed for monoxenous trypanosomatids, i.e., LIT and Schneider’s medium, the last typically used for Leishmania species. Furthermore, the parasite’s limited growth at 32 °C, a temperature usually employed for Leishmania axenic amastigogenesis [25], indicates a certain degree of thermotolerance, although the reduction in C. fasciculata ΔΨm at 32 °C after 96 h suggests the loss of parasites’ viability in this adverse condition. The capability to withstand a higher temperature supports previous observations of the parasite’s ability to grow in environments with low nutrient availability and inconstant temperatures, such as the conditions in the regions where C. fasciculata has been reported causing mammal infections [4]. Interestingly, in co-culture experiments, C. fasciculata outgrew L. braziliensis, highlighting its faster growth rate under laboratory conditions. However, in contrast to prior studies with Crithidia coinfection, we observed that C. fasciculata COLPROT606 failed to survive at higher temperatures, such as 34 °C, suggesting that environmental temperature may limit some isolates to thrive in warmer conditions [14,26].
Following these findings, it is important to consider the “environment-biased selection hypothesis”, which suggests that in co-infection scenarios, laboratory culture conditions may favor the faster-growing monoxenous trypanosomatids, such as C. fasciculata, potentially overshadowing the growth of Leishmania species [27]. In our co-culture experiments, C. fasciculata consistently outgrew L. braziliensis, making it unfeasible to detect Leishmania parasites after extended incubation periods. This result supports the hypothesis mentioned, indicating that C. fasciculata outgrew the pathogenic species in laboratory conditions due to the monoxenous faster growth, which could lead to potential diagnostic misinterpretations in co-infections. On the other hand, in the vertebrate model, Leishmania outcompetes the monoxenous trypanosomatids, which may be eliminated by host immune responses, or even by the lack of other mechanisms to establishes infection [27].
Unlike pathogenic Leishmania species, C. fasciculata did not survive or proliferate within murine peritoneal macrophages despite an initial infection, indicating the absence of immune evasion mechanisms commonly associated with Leishmania infections [28,29]. However, Crithidia sp. CLA-KP1, isolated from the biting midge Culicoides peregrinus, was cleared from murine peritoneal exudate macrophages (PEMs) by 48 h [26]; whereas, the Brazilian clinical isolate Crithidia sp. LVH60 infected THP-1 cells for up to 72 h [11]. Another C. fasciculata isolated in Iran infected both J774 and THP-1 cells, yet the persistence was not specified [6]. The results presented here are aligned with findings on Leptomonas seymouri, which also failed to persist in mammalian macrophages during co-infection with L. donovani [15]. Together, these findings suggest that C. fasciculata isolated from Peru could lack the virulence factors required to cause disease in vertebrate hosts. Furthermore, its inability to establish infection in murine or golden hamster models, even under immunosuppressed conditions, further supports the conclusion that C. fasciculata could be non-pathogenic under the experimental conditions assayed here. Nevertheless, we emphasize that only a single mammalian isolate was tested and that experiments were conducted with a limited number of replicates, which restricts the generalization of our conclusions. Further in vivo and in vitro studies with additional isolates are necessary to clarify whether our findings are consistent across different strains and host contexts.
Another key aspect of our study was assessing C. fasciculata resistance to Amphotericin B, a drug commonly used for the treatment of leishmaniasis [30]. Our results showed that C. fasciculata parasites isolated from a human patient exhibited greater resistance to Amphotericin B compared to both L. braziliensis and the reference C. fasciculata strain. This result suggests that C. fasciculata may harbor intrinsic mechanisms of drug resistance, as supported by a previous study comparing growth inhibition between C. fasciculata and Leptomonas, where C. fasciculata required 3 to 6 times higher concentrations of phenanthridines and diamidines to achieve 50% inhibition compared to Leptomonas [31]. Although this result is intriguing, it should be interpreted with caution, since resistance was assessed only for Amphotericin B and exclusively in vitro. Additional experiments using different drugs and clinical isolates are required to evaluate the potential clinical implications of this phenomenon, especially in co-infection scenarios where resistance traits could be transferred or masked.
In the sandfly insect host, C. fasciculata was detected in all samples after artificial blood infection, indicating its ability to establish in the vector. A meta-analysis has shown that insects exhibit a higher prevalence of monoxenous trypanosomatids, likely due to a trade-off favoring dissemination over complex host adaptation [32]. This increased prevalence could favor contact with vertebrate hosts, presenting a challenge which should be overcome by the monoxenous parasite. In the presence of a dixenous parasite the survival of the monoxenous counterpart could be favored by a synergistic interaction between both trypanosomatids. In this context, studies reporting sandflies naturally infected with monoxenous trypanosomatids suggest that human infection with C. fasciculata could share similar transmission dynamics with pathogenic species [33,34,35,36,37]. Notwithstanding, the passage of the parasite through the vector could select for more virulent populations [37], and factors such as immunomodulatory molecules presented in sandfly saliva may also influence the success of infection [38]. Additional research should aim to evaluate these natural transmission conditions more closely to better understand how the vector influences the pathogenic potential of C. fasciculata, and to clarify the interaction dynamics of trypanosomatids in their natural environments.
Taken together, these findings underscore the need for improved diagnostic accuracy. Clinical isolates of C. fasciculata can be easily mistaken for Leishmania due to morphological similarity and the limitations of conventional tests, such as direct smears or immunological assays, especially in endemic areas [7,11]. Therefore, the use of molecular methods, including PCR and sequencing, is essential, as they have proven more effective in distinguishing C. fasciculata from pathogenic Leishmania, as demonstrated by recent qPCR-based studies in clinical isolates [7,11].
5. Conclusions
Over the last few years, an increased number of reports of monoxenous trypanosomatids in vertebrate hosts have emerged. However, the mechanisms involved in the pathogenesis and transmission of such parasites in co-infection with pathogenic trypanosomatids remain poorly understood. Our findings suggest that the Crithidia fasciculata COLPROT606 isolate from an atypical human infection can grow in vitro under moderate temperatures, colonize permissive sandfly vectors, outcompete L. braziliensis in co-culture, and exhibits partial resistance to Amphotericin B. However, it fails to persist or proliferate within mammalian macrophages and does not establish infection in immunocompetent or immunosuppressed BALB/c mice or golden hamsters, nor does co-infection with L. braziliensis enhance its survival. These results indicate that C. fasciculata COLPROT606 lacks the virulence and adaptive mechanisms required for sustained infection in vertebrate hosts, underlining the importance of molecular screening of atypical lesions and further investigation into its transmission and ecological interactions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hamida A. Ilyas D. Zahra M. Shehnaz G. Leishmaniasis: A Neglected Tropical Disease Glob. Immunol. Infect. Dis. Rev.20194172310.31703/giidr.2019(IV-I).03 · doi ↗
- 2Karmakar S. Volpedo G. Zhang W.-W. Lypaczewski P. Ismail N. Oliveira F. Oristian J. Meneses C. Gannavaram S. Kamhawi S. Centrin-deficient Leishmania mexicana confers protection against Old World visceral leishmaniasis NPJ Vaccines 2022715710.1038/s 41541-022-00574-x 36463228 PMC 9719514 · doi ↗ · pubmed ↗
- 3Maslov D.A. Votýpka J. Yurchenko V. LukešJ. Diversity and phylogeny of insect trypanosomatids: All that is hidden shall be revealed Trends Parasitol.201329435210.1016/j.pt.2012.11.00123246083 · doi ↗ · pubmed ↗
- 4Toledo P.D. Comportamiento In Vitro de um Kinetoplastido Trypanosomatidae Aislado de uma Lesion Cutanea Licenciate Thesis Universidad Nacional Siglo XX Llallagua, Bolivia 2007
- 5Maruyama S.R. de Santana A.K. Takamiya N.T. Takahashi T.Y. Rogerio L.A. Oliveira C.A. Milanezi C.M. Trombela V.A. Cruz A.K. Jesus A.R. Non-Leishmania parasite in fatal visceral leishmaniasis-like disease, Brazil Emerg. Infect. Dis.2019252088209210.3201/eid 2511.18154831625841 PMC 6810192 · doi ↗ · pubmed ↗
- 6Ghobakhloo N. Motazedian M.H. Naderi S. Ebrahimi S. Isolation of Crithidia spp. from lesions of immunocompetent patients with suspected cutaneous leishmaniasis in Iran Trop. Med. Int. Health 20192411612610.1111/tmi.1304229446852 · doi ↗ · pubmed ↗
- 7Boucinha C. Andrade-Neto V.V. Ennes-Vidal V. Branquinha M.H. Santos A.L.S. Torres-Santos E.C. A stroll through the history of monoxenous trypanosomatids infection in vertebrate hosts Front. Cell. Infect. Microbiol.20221280470710.3389/fcimb.2022.80470735242719 PMC 8886219 · doi ↗ · pubmed ↗
- 8Fakhar M. Derakhshani-Nia M. Gohardehi S. Karamian M. Hezarjaribi H.Z. Mohebali M. Akhoundi B. Sharbatkhori M. Domestic dogs carriers of Leishmania infantum, Leishmania tropica, and Crithidia fasciculata as potential reservoirs for human visceral leishmaniasis in northeastern Iran Vet. Med. Sci.202282329233610.1002/vms 3.92936063538 PMC 9677403 · doi ↗ · pubmed ↗