Characterization of Gut Bacteria in Natural Populations of Sand Flies (Diptera: Psychodidae) from Endemic and Non-Endemic Areas of Leishmaniasis in Morocco
Mohamed Daoudi, Abdelkrim Outammassine, El Mahdi Redouane, Souad Loqman, Mohamed Hafidi, Ali Boumezzough, Martin Olivier, Samia Boussaa, Momar Ndao

TL;DR
This study explores the gut bacteria of sand flies in Morocco, finding differences between areas with and without leishmaniasis, and identifies new bacterial species in these insects.
Contribution
The first characterization of sand fly gut microbiota in Morocco, revealing species- and location-specific bacterial communities.
Findings
Bacillus pumilus was the most frequently detected gut bacterium in sand flies.
Bacillus simplex, Nocardia ignorata, and Serratia spp. were identified for the first time in specific sand fly species.
Significant differences in bacterial richness and CFU counts were observed between sand fly species and collection sites.
Abstract
Leishmaniasis is a vector-borne parasitic disease caused by Leishmania spp., transmitted to humans by phlebotomine sand flies. The development of Leishmania into infective metacyclic promastigotes occurs within the sand fly gut, where the bacterial microbiota plays a pivotal role in parasite development and transmission dynamics. This study aimed to characterize the gut bacterial composition of phlebotomine sand flies collected from both endemic (Lalla Aaziza) and non-endemic (Marrakech) regions of leishmaniasis in Morocco. We investigated the microbiota of Phlebotomus papatasi, P. sergenti, P. perniciosus, and P. longicuspis, all proven vectors of cutaneous and visceral leishmaniasis in the Old World, including Morocco, as well as Sergentomyia minuta, a potential vector in the Mediterranean basin. Gut bacteria were isolated using conventional microbiological techniques and identified…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1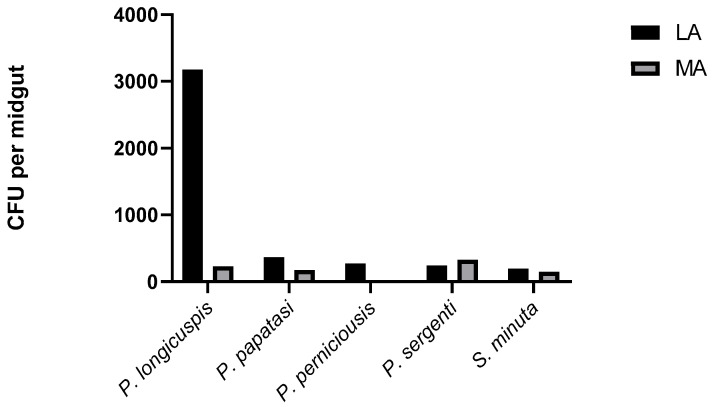 Figure 2
Figure 2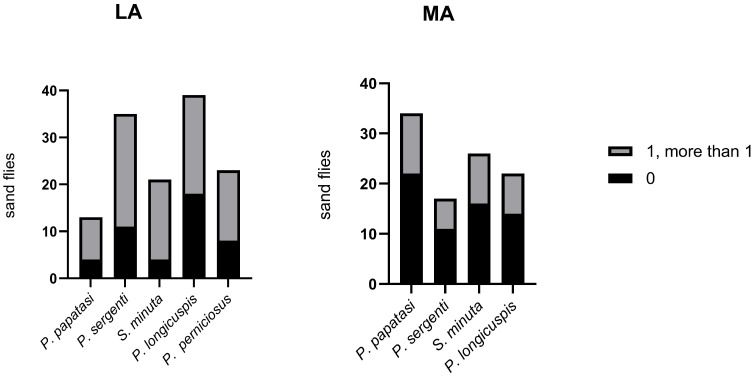 Figure 3
Figure 3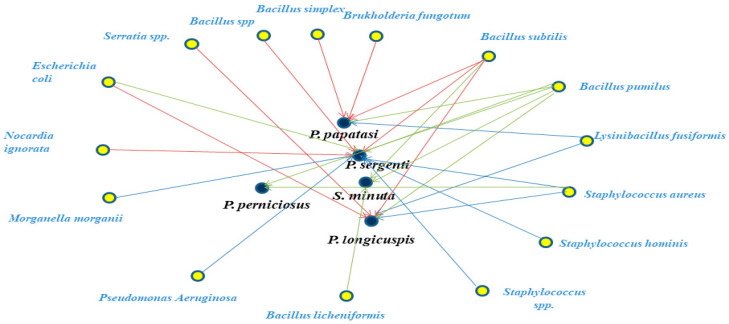 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsResearch on Leishmaniasis Studies · Insect symbiosis and bacterial influences · Insect and Pesticide Research
1. Introduction
Phlebotomine sand flies (Diptera: Psychodidae) are the exclusive natural vectors of leishmaniasis, a group of parasitic diseases caused by protozoan Kinetoplastid flagellates belonging to the genus Leishmania. Leishmaniasis affects between 12 and 15 million people in many countries located in the Mediterranean, tropical and subtropical regions, with around 350 million people at risk [1]. In Morocco, leishmaniasis remains a significant public health concern in both endemic and emerging regions. Cutaneous leishmaniasis (CL) caused by L. tropica has notably expanded since the 1980s across central and arid areas, becoming the most prevalent form of the disease [2]. Between 1997 and 2018, over 80,000 cases of human leishmaniasis were reported nationally [3]. In the northern Tangier–Tétouan–Al Hoceima region, 1255 cases were recorded, representing about 1.56% of the national burden, with Leishmania infantum responsible for most infections [4]. Visceral leishmaniasis (VL) remains endemic in northern Morocco, with approximately 100 cases reported annually, mainly affecting children under five years old [5]. The disease also has serious significant social and psychological consequences, as CL often results in permanent scarring and associated stigma [6]. Sand flies bear different microorganisms in their body and in their gut [7]. The sand fly gut hosts many microorganisms, including bacteria. Several studies have focused on the microbial community of both Phlebotomus and Lutzomyia species [8,9,10]. Bacterial symbionts significantly control some aspects of the physiology of their host [11]. Characterizing studies on insects’ gut microbiota and their role have been well investigated in recent years [12,13]. Studies focused on diverse mosquito species microbiota, including a potential influence on their vector competence [14,15]. Microorganisms, including bacteria, fungi, and protozoan parasites, can modulate insect vectorial capacity [16,17,18,19]. Furthermore, some investigations have shown a significant impact of the mosquito midgut microbiota in defense against malaria parasites [20]. However, several articles have been published on the effect of sand fly microflora on biology and sand fly–pathogen interaction. Volf et al. (2002) showed that Phlebotomus dubosqui had an associated gut bacterial cluster immediately after adult emergence [17]. Moreover, vector gut microbiota influences the development of virulent parasites, as demonstrated in L. infantum infection [21]. Some investigations on the midgut bacterial flora have been conducted on Lutzomyia longipalpis, Phlebotomus papatasi, P. perniciosus, P. tobbi, P. argentipes, P. duboscqi and Sergentomyia spp. [8,22,23,24]. Studies have shown that Phlebotomus papatasi harbors a low-diversity gut microbiota dominated by Enterobacteriaceae (e.g., Pantoea, Enterobacter, Serratia), which may influence Leishmania major development [25]. In contrast, P. perniciosus exhibits a richer and seasonally variable microbiota, including Ochrobactrum, Serratia, and Bacillus, potentially impacting L. infantum transmission [24]. A recent study by Cecilio et al. (2025) [26] demonstrated that the bacterium Delftia tsuruhatensis TC1, when introduced into the gut of Phlebotomus duboscqi sand flies, disrupts the development of L. major parasites. This disruption is attributed to gut dysbiosis induced by the bacterium, leading to reduced vector competence and decreased transmission potential [26]. These findings suggest that gut microbial composition may modulate vector competence and serve as an infection biomarker in Moroccan populations.
Our study aimed to examine the diversity of gut bacterial community colonization in field-caught sand flies through a culture-dependent method and to discuss its potential implications in sand fly vectorial competence.
2. Materials and Methods
2.1. Sand Fly Collection, Species Identification, and Gut Dissection
Sampling was carried out in Marrakech city (MA) (31°39′ N 8°01′ W) and Lalla Aaziza (LA) in Chichaoua province (31°04′ N 8°42′ W), located in the central region of Morocco (Figure 1). Lalla Aaziza is well-known as an active focus of cutaneous leishmaniasis due to Leishmania tropica in Morocco. In contrast, Marrakech is considered free of leishmaniasis [27].
At each site, six CDC miniature light traps were deployed and placed inside human dwellings. Traps were set in fixed indoor and outdoor locations in randomly selected habitations within the two study locations. Sand flies were collected throughout the activity season, between May and November 2018, yielding a total of 307 specimens. Sampling was conducted monthly over a seven-month period. Traps were deployed before sunset, sterilized with ethanol prior to use, and retrieved the following morning.
In this step, we followed the protocol described by Akhoundi et al. [28]. Each female sand fly was placed in a 1.5 mL microtube containing 30 µL of absolute ethanol and gently vortexed for 1 min to remove surface bacterial contaminants. Using sterile entomological needles, specimens were transferred into new sterile microtubes containing 30 µL of phosphate-buffered saline (PBS) to eliminate residual ethanol. Sand flies were then dissected under a stereomicroscope on a sterile slide with a drop of PBS. The head was removed, and the midgut was isolated, transferred into 30 µL of PBS into a new sterile microtube, and homogenized using a sterile glass pestle. Finally, 30 µL of the resulting suspension was used for bacterial analysis [28].
The genitalia and head of the sand flies were mounted on a slide, and species identification was performed morphologically using Boussaa’s identification key [29].
2.2. Bacterial Isolation and Identification
Bacterial identification was performed individually for each sample using standard microbiological techniques, including cultivation, isolation, and identification. From each 30 µL bacterial suspension, 15 µL was directly plated onto Plate Count Agar (PCA) medium (Cat. No. 105463, E. Merck Co., Darmstadt, Germany) to assess colony-forming units (CFU). The remaining 15 µL was inoculated into Brain Heart Infusion (BHI) broth and incubated at 37 °C for 24 h. PCA plates were incubated at 35 °C for 48 h, and the total number of colonies was counted. The CFU for each sample was calculated based on colony counts and dilution factors.
Following incubation in BHI, enriched cultures were plated onto various selective and differential media, including MacConkey Agar, Mannitol Salt Agar, Bile Esculin Agar, Cetrimide Agar, Trypticase Soy Agar, and Blood Agar supplemented with Amphotericin B (2 µg/mL). All plates were incubated at 37 °C for 24–48 h [28]. Isolated colonies were then analyzed by macroscopic (colony size, shape, edge, and optical properties), microscopic (Gram staining using the HiMedia Gram-stain Kit, HiMedia Laboratories Pvt. Ltd., Mumbai, Maharashtra, India), and biochemical methods (Catalase, Oxidase, DNase tests, and API 20E system). Final species-level identification was confirmed using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS), following established protocols and the manufacturer’s recommendations [30].
2.3. MALDI-TOF Assay
We employed Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry (MALDI-TOF MS) to identify bacterial strains isolated from sand flies. A single colony of each bacterial strain was directly smeared onto a designated spot on a metal target plate and allowed to air-dry. Subsequently, 1 µL of a saturated solution of α-cyano-4-hydroxycinnamic acid (CHCA) matrix was added to each spot, and the plate was again air-dried to facilitate co-crystallization of the matrix with bacterial proteins. The prepared target plate was then introduced into the MALDI-TOF MS instrument (Bruker Daltonics, Bremen, Germany), where it was subjected to laser irradiation under vacuum conditions. The ionized proteins were accelerated in an electric field, and their time-of-flight (TOF) to the detector was measured, generating a mass spectrum for each isolate. These spectra were analyzed using Biotyper 2.0 software (Bruker Daltonics), which compared them against a reference database to identify bacterial species based on their unique protein fingerprints (see Supplementary Material).
2.4. Statistical Analysis
Two-way analysis of variance (ANOVA) was performed based on colony counts and bacterial species richness across localities, using SPSS software ver. 18 to detect statistical differences in CFU and bacterial populations isolated from sand fly guts. Cytoscape software (version 3.9.1; http://www.cytoscape.org; accessed on 2 September 2020) was employed to construct and visualize networks of associations between sand fly species and their gut bacteria. In these networks, nodes represented either a bacterial species or a sand fly host, while edges indicated confirmed isolation links. Graphical representations and statistical analyses were generated using Microsoft Excel and GraphPad Prism ver. 10.2.3.
3. Results
A total of 175 specimens belonging to 7 species were collected in Lalla Aaziza (LA), Phlebotomus sergenti (45%), P. perniciosus (15%), P. longicuspis (11%), P. papatasi (9%), Sergentomyia minuta (10%), S. fallax (6%), and S. dreyfussi (4%), with a sex ratio of 0.8. In Marrakech (MA), 134 specimens representing five species were captured: Phlebotomus papatasi (40%), P. sergenti (18%), P. longicuspis (6%), S. minuta (23%), and S. fallax (13%), with a sex ratio of 0.6.
Using a culture-dependent methodology, we successfully isolated the bacteria colonizing the midgut of sand flies. Two-way ANOVA revealed a statistically significant interaction between sand fly species and locality (p = 0.033). At the species level, significant differences in CFU counts were observed for P. papatasi (p < 0.05) and P. longicuspis (p < 0.05). In contrast, no significant differences were detected for P. sergenti and S. minuta. The mean number of colonies per midgut varied among species, with P. longicuspis from LA showing the highest CFU (3.1 × 10^2^) (Figure 2). The distribution of sand flies carrying no bacteria, a single bacterium, or multiple bacteria also varied among species (Figure 3). Overall, bacterial infection rates appeared higher in LA compared to MA. However, this difference could not be statistically assessed due to unequal sample sizes between localities (Figure 3).
MALDI-TOF identification of bacterial species allowed us to detect fifteen different strains belonging to three phyla (Firmicutes, Proteobacteria, and Actinobacteria) in five sand fly species (P. sergenti, P. papatasi, P. longicuspis, P. perniciosus, and S. minuta) from the Lalla Aaziza and Marrakech localities. Among these phyla, Firmicutes was the most dominant, followed by Proteobacteria and Actinobacteria (Table 1).
Table 2 shows that bacterial species richness varies according to both locality and sand fly species. ANOVA revealed significant differences between localities and sand fly species for P. papatasi and P. sergenti (p < 0.05). Notably, P. sergenti exhibited the highest bacterial richness among the five species studied (7 bacterial species, BS) (Table 2).
Of the fifteen bacterial strains identified, seven were Gram-negative and eight were Gram-positive. Members of the genus Bacillus were the most frequently isolated, with Bacillus pumilus, Bacillus subtilis, Bacillus licheniformis, and Lysinibacillus fusiformis being the most common species (Table 1). These taxa also represented the most abundant midgut colonizers across the four sand fly species examined from both localities (Table 3). Results obtained using culture-dependent methods are summarized in Table 3.
At the species level, bacterial composition differed between the two localities (Table 2 and Table 3). In P. sergenti, the gut bacterial profile from Lalla Aaziza consisted of B. pumilus (38.5%), Staphylococcus hominis (7.7%), Staphylococcus spp. (7.7%), Staphylococcus aureus (15.4%), Escherichia coli (15.4%), Morganella morganii (7.7%), and Pseudomonas aeruginosa (7.7%). In contrast, specimens from Marrakech harbored B. pumilus (33%), B. subtilis (16.6%), Bacillus sp. (16.6%), E. coli (16.6%), and Nocardia ignorata (16.6%).
For P. longicuspis, the gut composition in Lalla Aaziza included B. pumilus (71.4%), S. aureus (14.6%), and L. fusiformis (14%). In Marrakech, isolates included B. pumilus (40%), B. subtilis (20%), E. coli (20%), and Serratia spp. (20%) (Table 3).
In P. papatasi, bacteria isolated from LA included B. pumilus (50%) and L. fusiformis (50%), while in MA, the gut contained B. pumilus (47.4%), B. subtilis (14.2%), Bacillus simplex (14.2%), and Burkholderia fungorum (14.2%) (Table 3). From S. minuta, B. pumilus (50%) and B. licheniformis (50%) were isolated in LA, whereas B. pumilus (75%) and B. subtilis (25%) were found in MA (Table 3). In P. perniciosus from LA, the gut microbiota consisted of B. subtilis (60%), S. hominis (20%), and S. aureus (20%) (Table 3).
A comparison of taxonomic composition between the two localities at the species level revealed 11 bacterial species in Marrakech and 8 in Lalla Aaziza. Interestingly, five species B. subtilis, B. simplex, Serratia spp., Burkholderia fungorum, N. ignorata, and B. licheniformis were found exclusively in Marrakech. Notably, B. simplex was detected for the first time in P. papatasi. Similarly, N. ignorata was reported for the first time in P. sergenti and Serratia spp. in P. longicuspis.
Overall, Gram-positive bacteria were more prevalent across the sand fly species examined, except in P. longicuspis, where Gram-negative bacteria were dominant. Network analysis further illustrated the associations between sand fly species and the bacterial taxa isolated in this study (Figure 4).
4. Discussion
Morocco is considered one of the countries where leishmaniasis is a major public health concern, and sand fly species exist within a significant geographical range [31,32]. The genus Phlebotomus includes the vectors of leishmaniasis in Morocco: Phlebotomus papatasi, the proven vector of zoonotic cutaneous leishmaniasis due to L. major [33]; P. ariasi, P. longicuspis and P. perniciosus, vectors of visceral leishmaniasis due to L. infantum [34]; and P. sergenti, the proven vector of anthroponotic cutaneous leishmaniasis caused by L. tropica [35].
Sand fly populations in the Mediterranean region, including Morocco, are recognized vectors of both Leishmania spp. and phleboviruses [36]; however, their vectorial competence, particularly regarding bacterial associations, remains underexplored. To investigate the bacterial composition of natural sand fly populations in Morocco, field studies were conducted in two central regions: Lalla Aaziza, an active focus of cutaneous leishmaniasis caused by L. tropica [37] with occasional cases of visceral leishmaniasis due to L. infantum [38], and Marrakech, which is considered a non-endemic area for leishmaniasis [39].
Sand flies are the only proven vectors of Leishmania species for humans [40]. Their vectorial capacity can be significantly influenced by symbiotic microorganisms [41,42,43], with bacteria representing promising targets for potential disease control strategies [44]. Understanding sand fly–bacteria–Leishmania interactions may provide new approaches for vector control and reduction in transmission. We therefore evaluated the bacterial composition of Moroccan phlebotomine sand flies and described the abundance of gut bacteria in different species and localities using a culture-dependent method and MALDI-TOF identification.
This identification technique has shown high potential for microbial identification in clinical and food sectors, as it is fast, easy to handle, and highly discriminatory down to the strain level [45]. Our results identified the gut bacterial flora of sand flies, which may contribute to better knowledge of their microbiota and to the development of new biological tools for disease control [46]. Several studies on the gut microbiome of sand flies in South America have applied classical molecular ecology techniques such as DGGE and 16S rRNA sequencing [10,18]. In Morocco, only one study has examined the intestinal microbiome of sand flies (P. papatasi compared to P. duboscqi) using DNA sequencing [47]. This is the first study to use a culture-dependent method coupled with MALDI-TOF to investigate the gut microbiota of Moroccan sand flies.
Our study on the gut flora of wild-caught sand flies (P. papatasi, P. sergenti, P. longicuspis, P. perniciosus, and S. minuta) demonstrated the presence of a wide diversity of bacterial strains belonging to the phyla Firmicutes (8 strains), Proteobacteria (6 strains), and Actinobacteria (1 strain). This result confirms previous investigations on sand fly gut microbiota, showing that Proteobacteria and Firmicutes are the predominant phyla [16,48,49]. Several studies carried out on the gut of natural populations or laboratory-reared sand flies have revealed the presence of diverse bacterial genera, including Bacillus, Staphylococcus, Burkholderia, Escherichia, Pseudomonas, Serratia, Cellulomonas, Chloroflexi, Citrobacter, Microbacterium, Enterobacter, Flavimonas, Gordonia, Klebsiella, Maltophila, Brevibacterium, Micrococcus, Morganella, Ochrobactrum, Oligella, Pantoea, Shigella, Sphingobacterium, Stenotrophomonas, Streptococcus, Acinetobacter, and Weeksella [8,24,25]. The genera Bacillus, Staphylococcus, Burkholderia, Escherichia, Pseudomonas, Serratia, and Morganella were also detected in our samples. We observed that variations in sand fly gut microbiota between species and localities (Figure 2 and Table 2) may be explained by the influence of microhabitats and physicochemical properties such as pH, oxygen availability in the insect gut, and food sources [50]. In the present study, Gram-positive strains (8) were isolated more frequently than Gram-negative strains (7). Additionally, in P. longicuspis, Gram-negative bacteria were the most prevalent strains, particularly in the Marrakech locality. Previous investigations have reported that the presence and prevalence of Gram-negative bacteria can completely or partially influence pathogen development in the insect midgut [25]. Moreover, the high prevalence of microbial infections in sand fly guts could negatively affect Leishmania transmission in endemic areas [51].
Notable variations in gut bacterial composition were observed among sand fly species across the two study localities. Among the dominant strains, Bacillus pumilus (Gram-positive) was detected in all sand fly species except P. perniciosus. The widespread occurrence and high abundance of B. pumilus may be attributed to its resistance to extreme environmental conditions, as previously reported by Kempf et al. (2005) [52]. Interestingly, members of the Bacillus genus exhibited host specificity and geographical variation. For example, Bacillus subtilis was exclusively isolated from sand flies collected in Marrakech (P. sergenti, P. papatasi, P. longicuspis, and S. minuta), whereas in Lalla Aaziza, it was only found in P. perniciosus. Although B. subtilis is considered non-pathogenic to humans [53], it is widely used in agriculture as a biofertilizer [54] and has been reported to possess probiotic properties [55,56]. These findings suggest that both environmental factors and host species influence the composition and distribution of sand fly gut microbiota, with Bacillus species showing ecological and biological adaptability. The isolation of B. subtilis from P. papatasi highlights its potential as a candidate for paratransgenic approaches, although other genera were detected less frequently. P. sergenti, P. papatasi, P. perniciosus, and P. longicuspis are well-known vectors of Leishmania spp. in Morocco and elsewhere [34,35]. S. minuta could also be a potential vector of L. infantum and L. tropica in Morocco [57].
The high bacterial diversity observed in sand flies may be linked to vectorial competence. For example, the gut microbiota of Lutzomyia longipalpis (the vector of VL in the New World) has been shown to play an important role in Leishmania development [21]. A study by Fraihi et al. (2017) reported that seasonal variation in P. perniciosus gut microbiota was associated with L. infantum transmission periods [24]. The identification of Serratia spp. and B. subtilis in sand flies in our study suggests these species may be suitable candidates for paratransgenic strategies [14,19,58]. Interestingly, B. subtilis was identified only in Marrakech, a non-endemic area of leishmaniasis in Morocco. This bacterium has already been used in paratransgenic mosquitoes and sand flies [55,56].
Some bacteria detected, such as Escherichia coli, are commonly associated with humans. Their presence may reflect environmental contamination from human or animal waste, or contact with contaminated soil, rather than stable colonization of sand fly guts. Although our study provides valuable insights into the bacterial communities of sand flies from endemic and non-endemic areas of Morocco, some limitations should be noted. Sampling was conducted monthly from May to September, covering only the peak sand fly activity season, so seasonal variations outside this period were not assessed. Moreover, the use of culture-dependent bacterial identification may underestimate the full microbial diversity present. These factors should be considered when interpreting the potential role of sand fly microbiota in vectorial competence.
5. Conclusions
This study provides the first characterization of gut bacterial communities in Moroccan sand flies from both endemic and non-endemic regions of leishmaniasis. A total of fifteen bacterial strains from three phyla were identified, with Bacillus pumilus being the most frequently detected species. Notably, Bacillus simplex (in P. papatasi), Nocardia ignorata (in P. sergenti), and Serratia spp. (in P. longicuspis) were reported for the first time in these vectors, highlighting previously unrecognized microbial associations. Significant differences in bacterial richness and colony-forming units were observed between sand fly species and localities, underscoring species and environment-dependent variation in gut microbiota. These findings provide a critical baseline for future studies on the role of gut bacteria in sand fly physiology and vector competence, and they open promising avenues for paratransgenic strategies to control leishmaniasis transmission in Morocco.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Torres-Guerrero E. Quintanilla-Cedillo M.R. Ruiz-Esmenjaud J. Arenas R. Leishmaniasis: A review F 1000 Research 2017675010.12688/f 1000 research.11120.128649370 PMC 5464238 · doi ↗ · pubmed ↗
- 2El Idrissi Saik I. Benlabsir C. Fellah H. Transmission patterns of Leishmania tropica around the Mediterranean basin: Could Morocco be impacted by a zoonotic spillover?P Lo S Negl. Trop. Dis.202216 e 001000910.1371/journal.pntd.001000935025884 PMC 8757988 · doi ↗ · pubmed ↗
- 3Hakkour M. Badaoui B. El Hamiani Khatat S. Sahibi H. Fellah H. Sadak A. Sebti F. Genetic diversity in Leishmania infantum and Leishmania tropica isolates from human and canine hosts in northern Morocco Gene 202492114848410.1016/j.gene.2024.14848438642857 · doi ↗ · pubmed ↗
- 4Hakkour M. El Alem M.M. Hmamouch A. Rhalem A. Delouane B. Habbari K. Fellah H. Sadak A. Sebti F. Leishmaniasis in Northern Morocco: Predominance of Leishmania infantum Compared to Leishmania tropica Biomed. Res. Int.20192019532728710.1155/2019/532728731485441 PMC 6702844 · doi ↗ · pubmed ↗
- 5Mouttaki T. Maksouri H. El Mabrouki J. Merino-Espinosa G. Fellah H. Itri M. Martin-Sanchez J. Soussi-Abdallaoui M. Chiheb S. Riyad M. Concomitant visceral and localized cutaneous leishmaniasis in two Moroccan infants Infect. Dis. Poverty 201873210.1186/s 40249-018-0413-829642944 PMC 5896130 · doi ↗ · pubmed ↗
- 6Bennis I. Thys S. Filali H. De Brouwere V. Sahibi H. Boelaert M. Psychosocial impact of scars due to cutaneous leishmaniasis on high school students in Errachidia Province, Morocco Infect. Dis. Poverty 201764610.1186/s 40249-017-0267-528385151 PMC 5383955 · doi ↗ · pubmed ↗
- 7Mc Carthy C.B. Diambra L.A. Rivera Pomar R.V. Metagenomic analysis of taxa associated with Lutzomyia longipalpis, vector of visceral leishmaniasis, using an unbiased high-throughput approach P Lo S Negl. Trop. Dis.20115 e 10310.1371/journal.pntd.0001304 · doi ↗
- 8Hillesland H. Read A. Subhadra B. Hurwitz I. Mc Kelvey R. Ghosh K. Das P. Durvasula R. Identification of aerobic gut bacteria from the kala azar vector, Phlebotomus argentipes: A platform for potential paratransgenic manipulation of sand flies Am. J. Trop. Med. Hyg.20087988188610.4269/ajtmh.2008.79.88119052297 · doi ↗ · pubmed ↗