A DNA Barcode Inventory of Austrian Dragonfly and Damselfly (Insecta: Odonata) Species
Lukas Zangl, Iris Fischer, Marcia Sittenthaler, Andreas Chovanec, Patrick Gros, Werner Holzinger, Gernot Kunz, Andrea Lienhard, Oliver Macek, Christoph Mayerhofer, Marija Mladinić, Martina Topić, Sylvia Schäffer, Kristina M. Sefc, Christian Sturmbauer, Elisabeth Haring

TL;DR
This study creates a DNA barcode library for 78 Austrian dragonfly and damselfly species to support biodiversity monitoring and conservation efforts.
Contribution
The study provides a comprehensive DNA barcode library for Austrian dragonflies and damselflies using COI and 16S markers.
Findings
More than 90% of the DNA sequences for COI and 16S markers were successfully obtained and identified.
Both COI and 16S markers proved effective for species identification in most odonate families.
Challenges were observed in closely related genera with low genetic differences, but overall results were reliable.
Abstract
Dragonflies and damselflies are good indicators of the health of rivers, ponds, and wetlands. In Austria, 78 species of these insects have been recorded. While we know a lot about where they live and how threatened they are, genetic data have been largely lacking. This study begins to fill that gap by creating a DNA reference library for Austrian dragonflies and damselflies. We collected many dragonfly and damselfly individuals from across Austria and sequenced two key DNA markers: COI, commonly used for species identification, and 16S, which is often used to detect species from environmental samples like water (a method known as eDNA metabarcoding). More than 90% of the samples were successfully identified using both markers. Some challenges appeared with closely related species, where the genetic differences are very small, but overall, the results show that both markers work well for…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2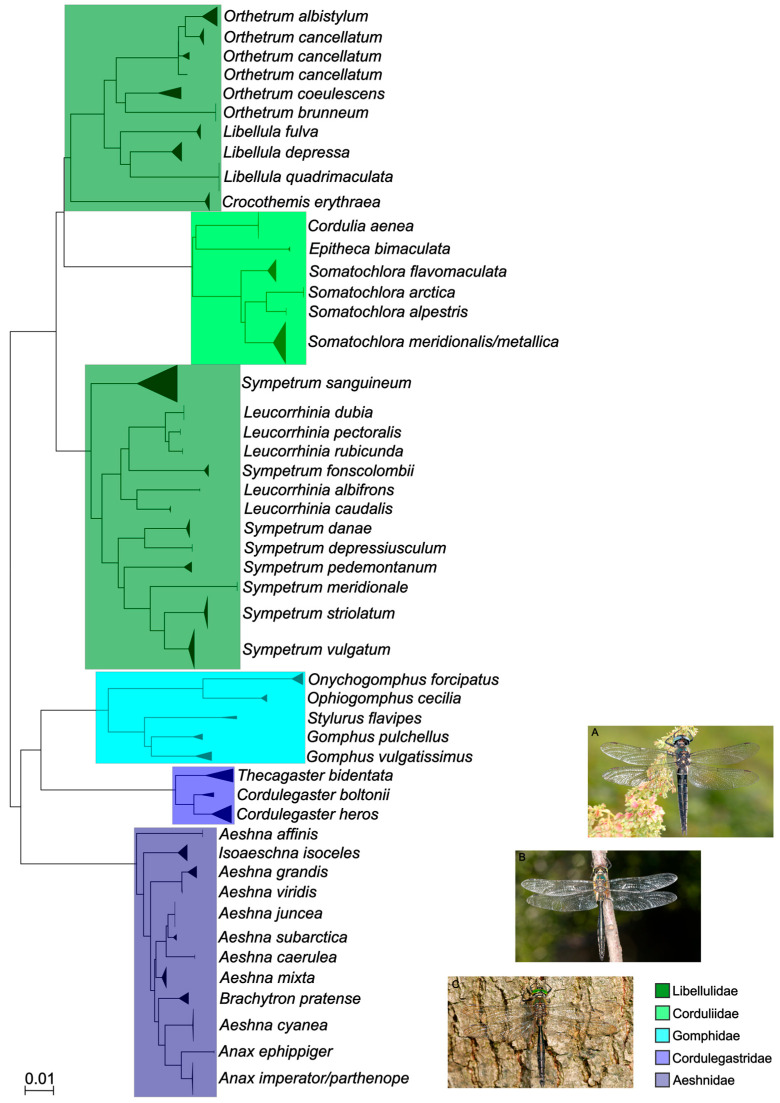 Figure 3
Figure 3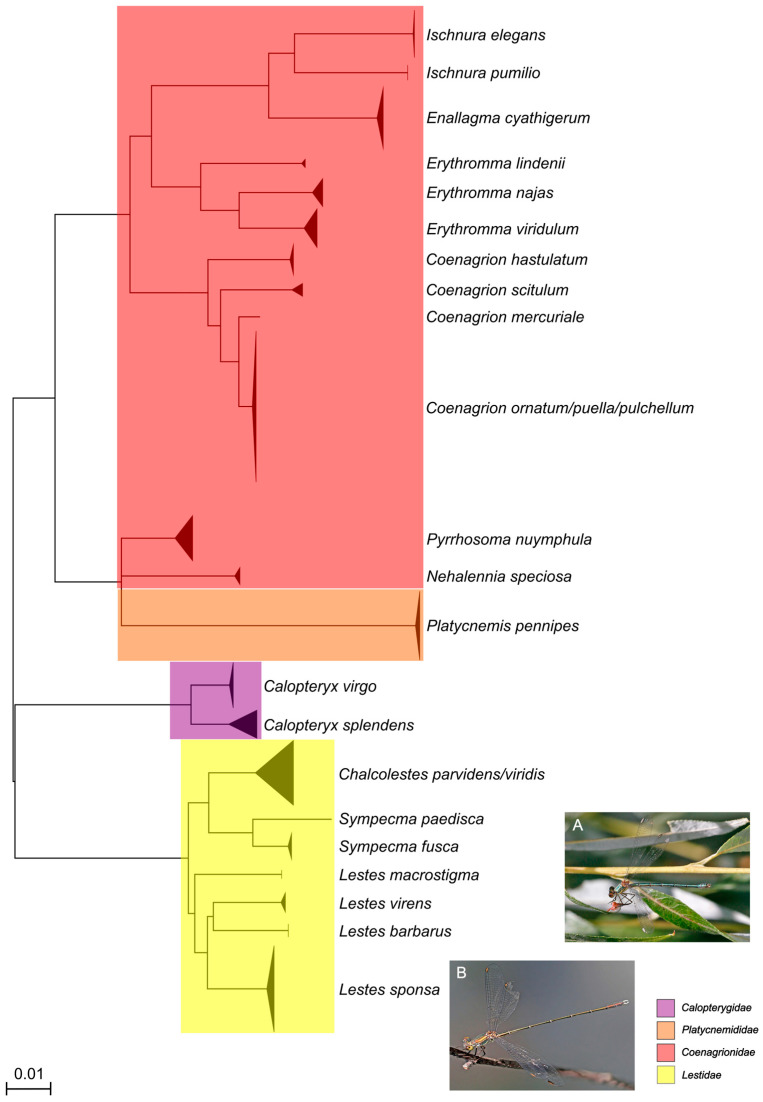 Figure 4
Figure 4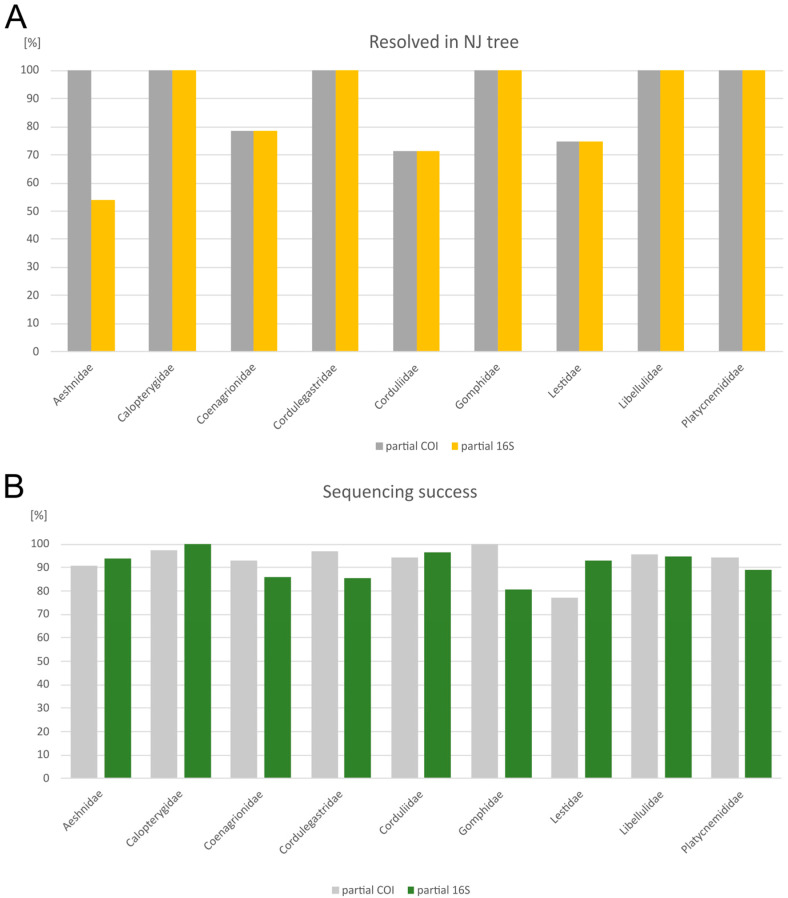 Figure 5
Figure 5- —Austrian Federal Ministry of Science, Research and Economy in the frame of ABOL (Austrian Barcode of Life) and an ABOL associated project within the framework of the “Hochschulraum-Strukturmittel”
- —Municipal Department 22—Environmental Protection (MA22)
- —European Agricultural Fund for Rural Development 2014–2020
- —University of Graz
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEnvironmental DNA in Biodiversity Studies · Freshwater macroinvertebrate diversity and ecology · Species Distribution and Climate Change
1. Introduction
The hemimetabolous insect order Odonata comprises three suborders, two of them—Anisoptera (dragonflies) and Zygoptera (damselflies)—are represented by 143 species throughout Europe [1,2,3]. Their relatively large body size and attractive coloration as well as their fascinating biology make them appealing to nature enthusiasts and scientists and have led to a relatively comprehensive knowledge about their diversity, distribution, reproductive behavior, and environmental requirements [1,3,4,5,6,7,8,9]. Based on this advanced state of knowledge, particularly about their amphibiotic life history and ecological preferences, Odonata are also used for environmental assessments of (semi-)aquatic habitats and monitoring within the European Water Framework Directive [10]. For Austria, several indices based on Odonata have been developed for assessing the ecological status of different aquatic habitats and the evaluation of restoration measures [11,12,13]. All this work has also led to fairly up-to-date Red List assessments and national checklists. According to the latest version of the checklist of Austrian odonates [14], 78 species belonging to nine families were recorded in Austria, of which 44 are listed in one of the three “threatened” categories in the Red List from 2006 [15]. These numbers, however, will soon be updated as the new version of the Red List for Austria is currently in preparation. Furthermore, 16 species are listed in the Annexes II and/or IV of the Habitats Directive 92/43/EEC by the European Union [2,16]. Contrasting this comprehensive knowledge about their general biology, comparatively little genetic data was available for Odonata until recently. Most of the existing genetic data was generated in the context of phylogenetic studies focusing mainly on the genus/family level [8,17,18]. In addition, character-based DNA barcoding was proposed for species discovery in odonates [19]. However, until recently, genetic reference data for national or Europe-wide species assemblages of Odonata were scarce and incomplete [20]. Those data were generated only recently following the progression of DNA barcoding becoming widely used in biodiversity research and species identification (e.g., [21,22,23,24,25,26]). The first comprehensive European odonate reference data were made publicly available in 2021 [3,27,28]. However, since geographic coverage increases species identification success as well as accuracy of genetic diversity estimation [29], more DNA sequence data of European dragonflies and damselflies is needed. This especially applies in the light of increasing (non-invasive) DNA-based approaches to study and/or monitor biological diversity, i.e., through DNA extracted from exuviae [30] or environmental DNA (eDNA) metabarcoding approaches [31,32]. These approaches may either investigate whole communities [33] or target specific species of interest [32]. However, contrary to classical DNA barcoding based on the mitochondrial cytochrome c oxidase subunit 1 gene (COI) as the standard marker for animal taxa, the marker of choice for (eDNA) metabarcoding is still open for discussion and may be dependent on the research question [34,35]. While COI has become the standard marker for metabarcoding of insects (e.g., [36,37,38], (eDNA) metabarcoding studies that target a larger taxonomic diversity including both invertebrates and vertebrates often use other markers that contain less primer site variation even across higher taxonomic levels, thus increasing the likelihood of successful amplification of all components of bulk samples containing large taxonomic diversity [34,39]. One occasionally used marker for assessing aquatic biodiversity, often in combination with COI, is a region of the mitochondrial 16S rRNA gene [40,41,42,43,44].
In this study, we present a comprehensive and almost complete reference COI DNA barcode inventory of Austrian Odonata as well as their respective 16S sequences. We compare the performance of both markers with regard to sequencing success, species identification, and discrimination and ponder the suitability for eDNA approaches.
2. Materials and Methods
The present study includes sequences of 892 specimens (Table S1) from all nine federal states of Austria covering all extant native odonate families. From 2017 to 2020, 839 of them were collected under the following permits: ABT13-53S-7/1996-156, ABT13-53W-50/2018-2, ABT13-198250/2020-9, RU5-BE-1489/001-2018, RU5-BE-64/018-2018, RU5-BE-1489/002-2021, MA22-169437/2017, A4/NN.AB-10097-5-2017, A4/NN.AB-10200-5-2019, SP3-NS-3375/2019 (005/2019), FE3-NS-2650/2019 (009/2019), N-2020-68581/4-Has, and 08-NATP-845/1-2019 (007/2019). Imagines were caught with an insect net and larvae were collected by hand. Six specimens (Aeshna viridis Eversmann, 1836 and Stylurus flavipes (Charp, 1825), three specimens each, collected between 1968 and 1988) were obtained from the entomological collection of the Natural History Museum Vienna and an additional 53 sequences of Austrian odonates (24 sequences from [45], and 29 sequences from [3]) were retrieved from BOLD. Specimens were photographed and tissue samples (legs) were stored in pure ethanol. The specimens, either mounted dry or preserved in a water–ethanol–glycerol solution (5:85:10), were transferred to scientific collections. All collection and storage metadata of the 892 specimens analyzed here are available on BOLD (https://dx.doi.org/10.5883/DS-BAAZ).
Tissue samples for genetic investigations were processed at the Central Research Laboratories of the Natural History Museum Vienna (NHM Vienna) and the University of Graz. Sequencing efforts followed a three-pronged approach: (1) For the COI gene, we tried to amplify major parts or even the complete COI gene in order to potentially gain additional species-specific genetic information not contained in the Folmer region constituting the ‘classical’ DNA barcode sequence. (2) Alternatively, at least the Folmer region of the COI gene was recovered, and (3) a partial fragment of the mitochondrial 16S (16S) gene was sequenced for all newly caught specimens as another widely used eDNA marker. As protocols varied between the two laboratories, procedures for generating sequences are described separately in the following paragraphs.
For samples processed at the NHM Vienna: For DNA extraction, the coxa and a small piece of the femur were used as starting material. For freshly collected samples, a standard DNA extraction was performed with the DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol, with a final elution volume of 60 µL AE buffer. For DNA extraction of museum specimens (dry collection), as lower DNA yields were expected, the QIAamp DNA Micro Kit (Qiagen, Hilden, Germany) was used according to the manufacturer’s protocol with a final elution volume of 40 µL AE buffer. For COI, mostly PCR primers binding close to or within the flanking tRNA genes (tRNA Tyr gene and tRNA Leu gene) were used, amplifying a ~1600 bp long fragment at once (Table 1). Alternatively, a COI sequence was generated by combining two shorter, overlapping amplicons (between ~500 and 800 bp), which were generated in separate PCRs with different primers depending on the species. All primers used for PCR are listed in Table 1. Regardless of whether the final COI sequence was amplified in one piece or composed of two amplicons, the consensus sequences included the ~650 bp long standard DNA barcoding region (the Folmer region). PCRs of the 1600 bp long fragment were performed with the Qiagen Taq Polymerase (Qiagen, Hilden, Germany) in a volume of 50 μL containing 0.5 μL Taq polymerase (5 units/μL), 5 μL of 10 × PCR Buffer (Qiagen, Hilden, Germany), 10 μL of 5 × Q-Solution (Qiagen, Hilden, Germany), 1.5 mM MgCl_2_, 2.5 mM dNTP Mix, 0.5 μM of each primer, and 1 μL DNA template. The PCR cycling protocol included an initial denaturation at 94 °C for 3 min, followed by 35 cycles of denaturation at 94 °C for 60 s, annealing (for primer-specific annealing temperatures see Table 1) for 30 s and extension at 72 °C for 60 s. The final step was an extension at 72 °C for 10 min. For PCR amplification of the partial 16S fragment, the primer pair 16S-Odo-F1/16S-Odo-R2 (Table 1) resulting in an amplicon length of ~560 bp (60 °C annealing temperature) was used. PCRs with primers for 16S and the shorter overlapping COI amplicons were performed in a volume of 25 µL containing 0.25 µL Qiagen Taq Polymerase (5 units/µL; Qiagen, Hilden, Germany), 2.5 µL 10 × PCR Buffer (Qiagen, Hilden, Germany), 5 µL 5 × Q-Solution (Qiagen, Hilden, Germany), 1.5 µL of MgCl_2_ (1.5 mM), 0.5 µL of dNTP Mix (2.5 mM), 0.5 µL of each primer (50 pmol/µL), and 1 µL template DNA. The PCR cycling protocol was the same as for the COI amplification (see above). For museum specimens (Aeshna viridis, and Stylurus flavipes), however, only the 560 bp long 16S fragment and for COI the standard barcoding region, using PCR primer combinations amplifying two shorter (~200 and ~600 bp), overlapping fragments (ODO_LCO1490d/CO1-OdoCol-R1 and CO1-Zyg-F1/ODO_HCO2198d [45,46], both with an annealing temperature of 57 °C) were amplified. PCR reactions with DNA stemming from museum material were performed with the Multiplex PCR Kit (Qiagen, Hilden, Germany) in a volume of 25 µL, containing 12.5 µL Multiplex PCR Master Mix, 0.5 µM of each primer, and 2 µL of template DNA. The PCR cycling protocol included an initial denaturation at 95 °C for 15 min, followed by 40 cycles of denaturation at 94 °C for 30 s, annealing for 90 s and extension at 72 °C for 60 s. The final step was an extension at 72 °C for 10 min. For all PCR reactions, PCR success was checked on 1% agarose gels, and PCR products were subsequently purified with the QIAquick PCR Purification Kit (Qiagen, Hilden, Germany). All samples were sequenced in both directions (Microsynth, Balgach, Switzerland) using the PCR primers as well as two additional internal primers (Table 1). For the smaller fragments (16S and COI, from museum specimens), only PCR primers were used for sequencing.
For samples processed at the University of Graz: Extraction of whole genomic DNA followed a rapid Chelex protocol described in [47]. Subsequent PCRs for the amplification of COI were conducted in a total of 10 µL containing 7.05 µL of water, 1 µL of 10 × buffer BioTherm containing 15 mM MgCl_2_ (Gene Craft, Lüdinghausen, Germany), 0.35 µL of 1 mM dNTPs (1 mM), 0.1 µM of each primer, 0.5 units of BioTherm DNA polymerase (Gene Craft, Lüdinghausen, Germany), and 1 µL of template DNA using the primers ODO_LCO1490d, ODO_HCO2198d, Tyr-Odo-F, and Leu-Odo-R [45,46] (Table 1). Cycling conditions were as follows: three minutes of initial denaturation at 95 °C, followed by 45 cycles of denaturation at 95 °C for 30 s, varying annealing temperatures (Table 1) for 30 s and extension at 72 °C for one minute as well as a final extension phase at 72 °C for seven minutes. Identical settings were also used for amplifying the 16S fragment with the only exception being the primers which were the same as being used by the NHM Vienna. Success of PCRs were checked via a 2% agarose gel electrophoresis and successful PCR products were cleaned using Exo-Sap-IT Express PCR Product Cleanup (Applied Biosystems by Thermo Fisher Scientific, Waltham, MA, USA). Bidirectional Sanger sequencing was performed following [48] using the primers ODO_LCO1490d and ODO_HCO2198d or Tyr-Odo-F and Leu-Odo-R for COI and 16S-Odo-F1 and 16S-Odo-R2 for 16S (Table 1).
Trace files were checked with MEGA version 6 [49] and forward and reverse reads were combined to consensus sequences and aligned using the built-in MUSCLE algorithm. All newly generated COI and 16S sequences were uploaded to BOLD (https://dx.doi.org/10.5883/DS-BAAZ) and subjected to subsequent clustering (COI and 16S), genetic distance, and BIN assignment analyses (only COI). Furthermore, additional sequences of dragonfly and damselfly species from Austria available on BOLD [3,30,45] were appended to the dataset resulting in alignments of 839 COI sequences (658/1536 bp) and 867 partial 16S sequences (547 bp), respectively (Table S1).
The ‘Taxon ID Tree’ tool implemented on BOLD (applying the Kimura 2 Parameter distance model and the BOLD aligner including all codon positions and the pairwise deletion option) was used to illustrate species-specific clustering on separate neighbor joining trees (NJ) for Anisoptera and Zygoptera based on sequence similarity of partial COI sequences (covering the Folmer region). Maximum intra- and minimum interspecific genetic distances were calculated using the ‘Distance Summary’ and ‘Barcode Gap Analysis’ tools (K2P distance model, complete deletion of ambiguous characters or missing data, and BOLD aligner), also implemented on BOLD. Furthermore, assignment of individual COI sequences to BINs was also checked on BOLD. Additionally, separate alignments for Anisoptera and Zygoptera containing only full-length COI sequences (>1000 bp) were created in MEGA and used for NJ tree inference and for comparison with the shorter dataset covering the Folmer region (using the pairwise deletion option for ambiguous or missing data). These datasets, however, did not contain all available species anymore as full-length sequences were not available for all species (Anisoptera: 194 sequences, 38 species; Zygoptera: 113 sequences, 21 species). For 16S, sequences of Anisoptera and Zygoptera were aligned separately with MEGA and subsequently used for NJ tree inference and for mean between-group (species) genetic distance calculation. Finally, the overall performance of the two markers (as well as the standard and full-length dataset for COI) was compared for the distinct species based on sequencing success (ratio samples/sequences) and discriminatory power (resolution in phylogram).
3. Results
In this study, 786 new COI DNA barcode sequences and 867 partial 16S sequences were generated, covering all nine families, 27 genera, and 74 out of 78 species of dragonflies and damselflies reported from and/or present in Austria. All species, except Anax ephippiger (Burmeister, 1839) (2), Coenagrion mercuriale (Charpentier, 1840) (2), Sympecma paedisca (Brauer, 1877) (1), and Leucorrhinia albifrons (Burmeister, 1839) (2) were represented by at least three specimens. Only Coenagrion hylas (Trybom, 1889), C. lunulatum (Charpentier, 1840), Lestes dryas (Kirby, 1890), and Sympetrum flaveolum (Linnaeus, 1758) could not be collected or sequenced at all. Overall, the sequencing success (ratio of successfully recovered sequences compared to the overall number of specimens sampled) amounted to 94 percent of specimens for COI and 96 percent for 16S. The 74 morphologically identified species were represented by 73 distinct BINs (Table 2) and 71 distinct clusters or singletons in the COI NJ trees based on the short fragments (Figure 1 and Figure 2). BIN sharing was detected in Coenagrion ornatum (Selys, 1850)/C. puella (Linnaeus, 1758)/C. pulchellum (Vander Linden, 1825), Anax imperator Leach, 1815/A. parthenope Selys, 1939, Somatochlora meridionalis Nielsen, 1935/S. metallica (Vander Linden, 1825), and Chalcolestes parvidens (Vander Linden, 1825)/C. viridis (Vander Linden, 1825) (Figure 1 and Figure 2, Table 2). Chalcolestes parvidens and C. viridis shared two BINs (BOLD:AAI7225; BOLD:ADR7794). Coenagrion mercuriale, on the other hand, was represented by two different BINs (BOLD:ADS2145, BOLD:ACG0797). Furthermore, whereas C. ornatum/C. puella/C. pulchellum, S. meridionalis/S. metallica and C. parvidens/C. viridis constituted mixed clades in the NJ trees, Austrian Anax imperator and A. parthenope occupied two distinct monophyletic clades in the NJ tree and were clearly resolved as distinct entities despite their shared BIN. K2P distances within species ranged from 0 to 3.17% (10.21% in cases of deep intraspecific divergences, usually together with BIN sharing; mean: 0.55%) and between species from 2.30 to 16.91% (0% in cases of BIN sharing) resulting in a pronounced DNA barcoding gap for all species except (i) Coenagrion ornatum/C. puella/C. pulchellum, (ii) Somatochlora meridionalis and S. metallica, and (iii) Chalcolestes parvidens and C. viridis (Figure S1).
Similar results were also obtained for 16S. Somatochlora meridionalis/S. metallica, Anax imperator/A. parthenope, Coenagrion ornatum/C. puella/C. pulchellum, and Chalcolestes parvidens/C. viridis and could not be distinguished based on their respective 16S sequences either and were consequently forming mixed clades in the 16S trees (Figure 3 and Figure 4). Additionally, Orthetrum cancellatum (Linnaeus, 1758) was not recovered as a monophyletic group, but was divided in three clades which, however, exclusively contained O. cancellatum sequences (Figure 3). All other species could be clearly recognized by their respective 16S sequence and, consequently, were occupying distinct clades in the 16S NJ tree. Mean within- and between-group (species) genetic distances (Table S2) mirrored the results of COI genetic distances (Table 2) although 16S distances were approximately five times lower than COI distances.
The comparison between the standard (Folmer region; Figure 1 and Figure 2) and full-length COI dataset (Figures S2 and S3) revealed an equal performance in terms of species distinction in the NJ tree. In particular, unresolved morphospecies like in the genera of Somatochlora, Chalcolestes, or Coenagrion could also not be resolved with the full-length data set.
The comparison between the COI and the 16S datasets revealed that, in terms of sequencing success, both markers performed almost equally well (Figure 5). The sequencing success was very high in general (mostly above 90%) with the only drop below 80% for COI in the family Lestidae. For species discrimination, both genetic markers performed similarly. Sequences from Calopterygidae, Cordulegastridae, Gomphidae, Libellulidae, and Platycnemididae were unambiguously assigned to morphospecies level using both COI and 16S. However, Anax imperator and A. parthenope were resolved as two distinct clades sharing a single BIN in the COI tree, but formed a single mixed clade in the 16S tree. Both markers failed to differentiate among Coenagrion ornatum/puella/pulchellum, Somatochlora meridionalis/metallica, and Chalcolestes parvidens/viridis.
4. Discussion
In this study, we compiled an almost complete genetic reference database of Austrian dragonflies and damselflies consisting of 839 COI barcode sequences and 867 partial 16S sequences (Table S1). BIN assignment, genetic distance analysis, and clustering of COI sequences are generally in line with results obtained by [3,27,28], but a further increase in regional representation is important for reliable genetic species identification [3,20,29]. No new BINs were detected. Conflicting cases between morphological and genetic identification, which point to incidental hybridization or introgression, were generally corroborating those found by [3,27]. Somatochlora meridionalis and S. metallica shared BINs/haplotypes. Interestingly, there are already indications for potential hybridization between different (not Central European) Somatochlora species in the literature [50,51,52,53]. The patterns of BIN and haplotype sharing between Somatochlora meridionalis and S. metallica, however, does not necessarily imply hybridization, but could equally likely be due to incomplete lineage sorting of two recently diverged species. Indeed, the two species are morphologically very similar but also distinct [54] and closely related based on genomic data [55], and the species status of S. meridionalis is still disputed by some researchers [56]. Clear evidence for introgressive hybridization was found in Chalcolestes parvidens and C. viridis [3,8,27]. Unfortunately, these ambiguities could not be further resolved by adding more information from the COI region as we sequenced large parts or even the entire COI gene. Also, in Coenagrion ornatum/puella/pulchellum distinction at the species level was not achieved. For this group, although mtDNA data show BIN sharing across large parts of the species’ distribution range [3,27,57], nuclear data successfully separate the three species into distinct entities [27,58], corroborating the assumption that the observed BIN sharing might be due to introgressive hybridization and potentially mitochondrial capture over large parts of the species distribution ranges, rather than incomplete lineage sorting, which, in theory, could also be responsible for BIN sharing. Reports of heterospecific pairing and hybridization between C. puella and C. pulchellum have been published previously [59,60], adding to the plausibility of such a scenario. Regarding Chalcolestes, southeastern Austria represents the northernmost edge of the distribution range of C. parvidens [61], and hybrids are known from places where C. parvidens co-occurs with C. viridis, including the very eastern part of Austria [62]. Thus, hybridization followed by back-crossing might indeed be responsible for BIN-sharing between these two species.
Interestingly, while some taxa cannot be clearly resolved at species levels based on their DNA barcodes across their entire geographic ranges, our Austrian data show that accurate identification is still possible locally. For instance, on a Holarctic scale, both Aeshna juncea (Linnaeus, 1758) and A. subarctica Walker, 1908 comprise several BINs with some BIN sharing between the two species. In addition, the two species are not reciprocally monophyletic based on DNA barcodes. These patterns not only indicate geographic structure but also introgressive hybridization and or (ancient) incomplete lineage sorting, if we assume morphological identification of the specimens was correct [63]. In Austria, as for the rest of Europe [3,27], the two species comprise one single BIN each with no indication of interspecific hybridization. BIN sharing between Anax imperator and A parthenope has been shown before, with all A. imperator and A. parthenope constituting a single BIN [3,27,28] and evidence for haplotype sharing [59]. Even though, in our Austrian data, the two species also form a single BIN, they fall into two reciprocally monophyletic groups. However, considering the range-wide pattern of some haplotype sharing and phenotypic evidence for hybridization between the two species [64], DNA barcode-based species assignment in this species pair cannot be performed with 100% confidence in Austria, too.
In general, however, sequencing and species identification success based on COI barcode sequences was very high for most specimens confirming its applicability for genetic biodiversity assessments [3,27,65]. On the other hand, sequencing and identification success was also comparably high for 16S, which is a marker often considered for eDNA and metabarcoding approaches [66,67,68]. Naturally, the choice of marker is dependent on the research question and aspects like the universality of the primers and the species-level resolution as well as the coverage of respective reference databases (like BOLD or NCBI GenBank) which may influence the decision [29,66,69]. Despite ongoing discussions about primer choice in eDNA studies [68,70,71], the results of this study seem to underscore the suitability of 16S as a marker for eDNA studies in odonates, due to both the discriminatory power on the species level as well as the availability of highly conserved primer regions (Table 1).
Both, DNA barcoding as well as eDNA metabarcoding, gain increasing importance for the identification and monitoring of (rare and/or threatened) odonate species, especially with regard to non-invasive sampling [30,32,72]. Considering that Austria is home to 11 of the 16 species protected by the EU Habitats Directive (Directive 92/43/EEC, annexes II and/or IV) and that 56% of the 78 species reported from Austria fall into one of the three ‘threatened’ categories [15], proper monitoring is not only a legal obligation but a conservation necessity. For the present study, four species (Coenagrion hylas, Coenagrion lunulatum, Lestes dryas, and Sympetrum flaveolum) which have been reported from Austria until the beginning of the 21st century [72,73,74,75,76] were not found and/or could not be sampled at all and for another four species, fewer than three samples could be acquired each. To some extent, this might be indicative of various threats like changes in land use, loss of habitat, increased nutrient deposition, or more rapid desiccation (especially of bogs) due to climatic change, entailing higher temperatures and more frequent and severe droughts impacting the larvae of these species. Two of these species, C. hylas and C. lunulatum, are known to be very rare in Austria. Coenagrion hylas is a glacial relict species with, apart from some location in northwestern Russia, vital populations in Europe known only from a very limited area in the northern Calcareous Alps of Tyrol, Austria, and might hence be considered the rarest European zygopteran species [56,77]. Coenagrion lunulatum is a Euro-Siberian species, with very few Austrian records over the last decades [56,78]. The other two species, L. dryas and S. flaveolum, have an allegedly wide distribution, also in Austria [56]. Both species, however, rely on ephemeral habitats, such as flooded meadows and swampy depressions, types of habitat that have become increasingly rare in Austria. These threats may also change over time [79] and can lead to re-arrangements of regional diversity, i.e., species composition [80,81]. Some species have already been reported to expand their altitudinal and latitudinal range [80,82,83,84] as a consequence of changing environments and climate change, but since this is not possible for every species, for example, cold-adapted higher-altitude species, they could be threatened by extinction. However, predictions and evaluations of these dynamics are often difficult to make as population trends of endangered and protected species are still unclear in many parts of Europe. Furthermore, several endangered species are not even listed in the annexes of the Habitats Directive of the European Union [85]. Therefore, increased monitoring efforts are needed to track changing population trends and regional Odonata community assemblages and provide data for informed directions of conservation priorities. Approaches like the genetic identification of larvae, the non-invasive DNA barcoding of exuviae, species delimitation by taxonomic lay people, or large-scale monitoring via eDNA [30,31,32] all rely on reliable and comprehensive reference data which we have provided in this study to make them available for both scientists and conservationists alike.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sahlén G. Kalkman V. Boudot J. Bernard R. Conze K. De Knijf G. Dyatlova E. Ferreira S. Jovic M. Ott J. European Red List of Dragonflies Publications Office of the European Union Luxemburg 201010.2779/84650 · doi ↗
- 2Kalkman V. Boudot J. Bernard R. De Knijf G. Suhling F. Termaat T. Diversity and conservation of European dragonflies and damselflies (Odonata)Hydrobiologia 201881126928210.1007/s 10750-017-3495-6 · doi ↗
- 3Geiger M. Koblmüller S. Assandri G. Chovanec A. Ekrem T. Fischer I. Galimberti A. Grabowski M. Haring E. Hausmann A. Coverage and quality of DNA barcode references for Central and Northern European Odonata Peer J 20219 e 1119210.7717/peerj.1119233986985 PMC 8101477 · doi ↗ · pubmed ↗
- 4De Marchi G. Precopulatory reproductive isolation and wing colour dimorphism in Calopteryx splendens females in southern Italy (Zygoptera: Calopterygidae)Odonatologica 199019243250
- 5Reinhardt K. Gerighausen U. Oviposition site preference and egg parasitism in Sympecma paedisca (Odonata: Lestidae)Int. J. Odonatol.2001422123010.1080/13887890.2001.9748169 · doi ↗
- 6Rivera A. Andrés J. Córdoba-Aguilar A. Utzeri C. Postmating sexual selection: Allopatric evolution of sperm competition mechanisms and genital morphology in calopterygid damselflies (Insecta: Odonata)Evolution 20045834935910.1111/j.0014-3820.2004.tb 01650.x 15068351 · doi ↗ · pubmed ↗
- 7Fincke O. Jödicke R. Paulson D. Schultz T. The evolution and frequency of female color morphs in Holarctic Odonata: Why are male-like females typically the minority?Int. J. Odonatol.2005818321210.1080/13887890.2005.9748252 · doi ↗
- 8Dijkstra K.D.B. Kalkman V.J. Phylogeny, classification and taxonomy of European dragonflies and damselflies (Odonata): A review Org. Divers. Evol.20121220922710.1007/s 13127-012-0080-8 · doi ↗