Postharvest Disease Management of ‘Akizuki’ Pear in China: Identification of Fungal Pathogens and Control Efficacy of Chlorine Dioxide
Haichao Jiang, Lixin Zhang, Yang Zhang, Yudou Cheng, Cunkun Chen, Yongxia Wang, Junfeng Guan

TL;DR
This study identifies fungal pathogens causing rot in stored 'Akizuki' pears in China and tests chlorine dioxide as an effective control method.
Contribution
The study identifies specific fungal pathogens and demonstrates the efficacy of chlorine dioxide as an alternative to conventional fungicides.
Findings
Alternaria alternata, Diaporthe eres, and Penicillium expansum were identified as major pathogens causing postharvest rot in 'Akizuki' pears.
Chlorine dioxide significantly inhibited the growth of these pathogens, with EC50 values comparable to conventional fungicides.
Chlorine dioxide disrupted cell membrane structures and suppressed mycelial growth of the tested fungal strains.
Abstract
The ‘Akizuki’ pear has become increasingly popular in China in recent years. However, the ‘Akizuki’ pear often suffers from severe rot diseases during the postharvest storage period. Those during storage have not been thoroughly elucidated In this study, fungal pathogens causing postharvest decay of ‘Akizuki’ pear were identified through multi-gene phylogenetic analysis, followed by assessment of the antifungal efficacy of chlorine dioxide (ClO2) at varying concentrations. A total of 18 strains were isolated and identified as pathogens by Koch postulates. The isolated pathogens were taxonomically identified by combining morphological characterization of hyphae/spores with multi-gene phylogeny (ITS, β-tub, tef1). The results revealed that isolates A1-A11 were identified as Alternaria alternata, D1-D3 as Diaporthe eres, P1 as Penicillium citrinum, and P2-P4 as Penicillium expansum. The…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
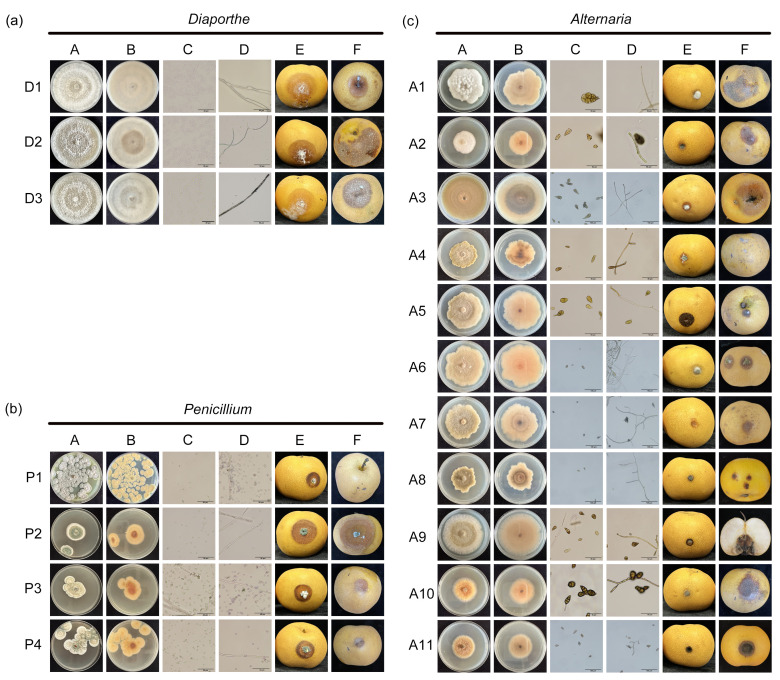 Figure 1
Figure 1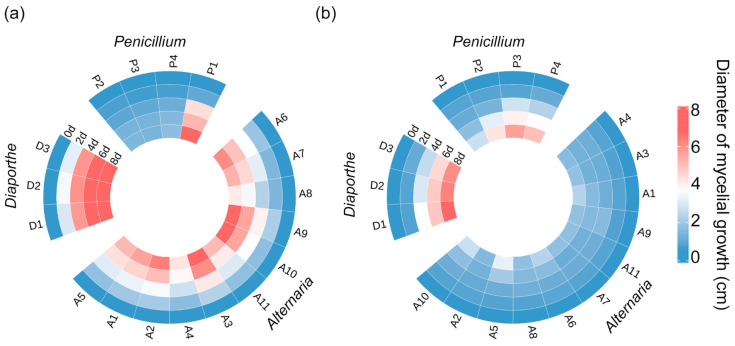 Figure 2
Figure 2 Figure 3
Figure 3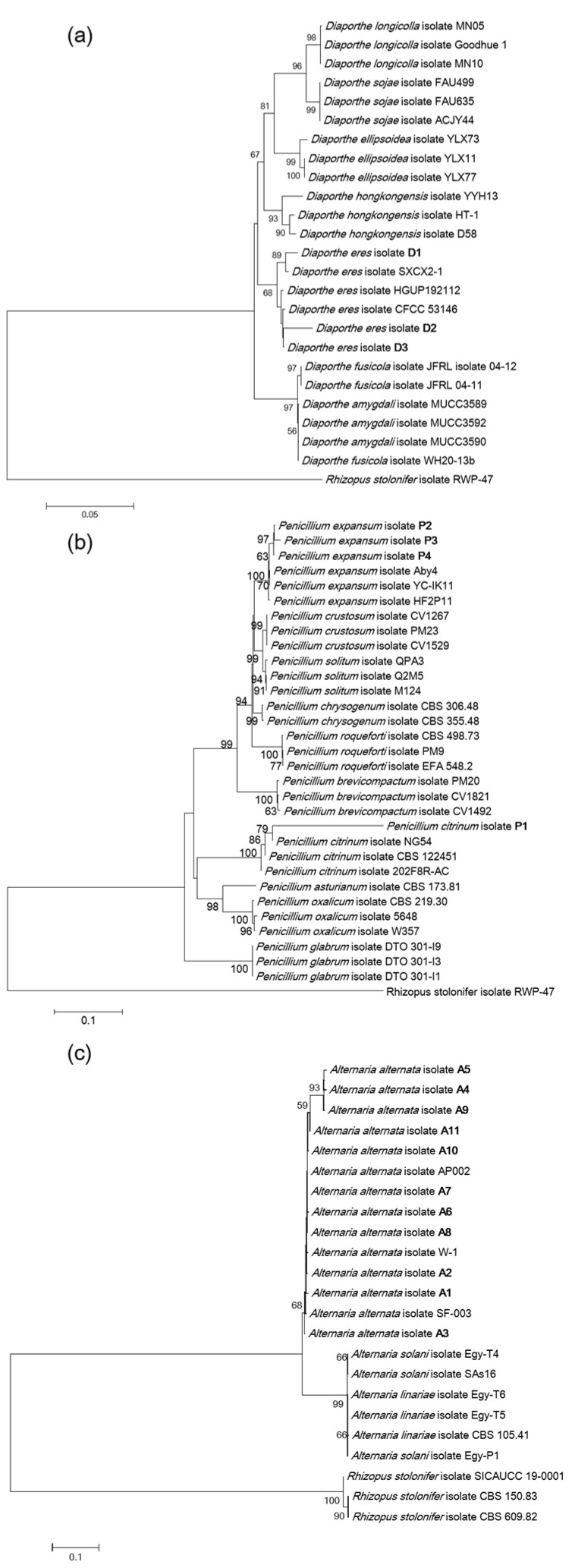 Figure 4
Figure 4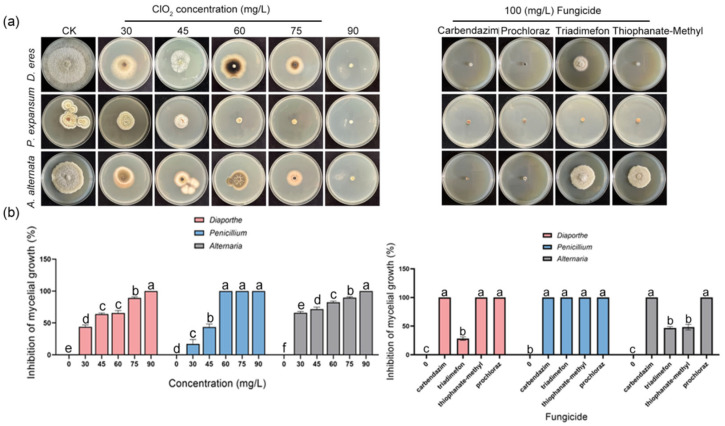 Figure 5
Figure 5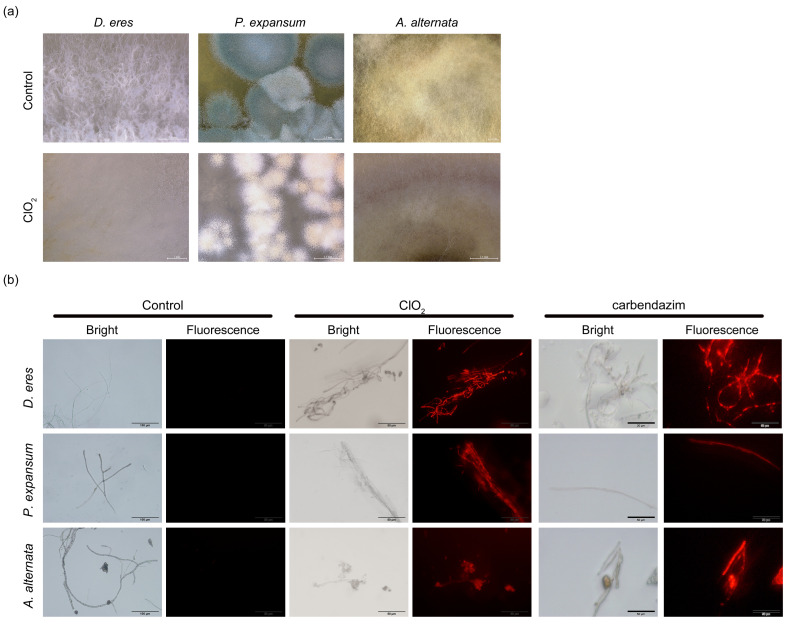 Figure 6
Figure 6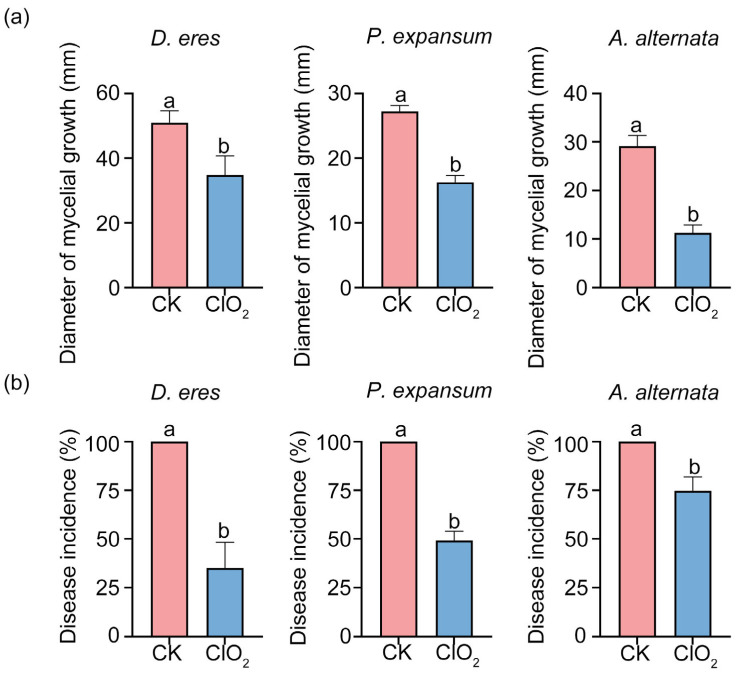 Figure 7
Figure 7- —Ministry of Finance, Ministry of Agriculture and Rural Affairs, Modern Agricultural Industry (Pear) Technology System Project
- —Agriculture Science and Technology Innovation Project of HAAFS
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Postharvest Quality and Shelf Life Management · Plant Physiology and Cultivation Studies
1. Introduction
‘Akizuki’ pear (Pyrus pyrifolia Nakai) is a mid-to-late maturing sand pear variety, rich in various organic acids, soluble sugars, and multiple phenolic compounds essential for human health [1], and is highly favored by consumers for its uniform fruit shape, tender flesh texture, and sweet flavor [2]. China is the world’s largest producer of the ‘Akizuki’ pear, with an annual production of 1,500,000 tons, covering an area of about 80,000 hectares. In recent years, it has been widely planted and is currently predominantly distributed in areas such as Wei County and Zhao County in Hebei, Laiyang city in Shandong, and Xinyi city, Suining, and Tongshan city in Jiangsu.
However, the ‘Akizuki’ pear is susceptible to pathogen infections that lead to fruit rot. These problems seriously affect the quality and commercial value of the ‘Akizuki’ pear [3]. According to a 2023 report by the USDA, postharvest diseases of pears in China cause an annual yield loss of approximately 8%, with the economic losses exceeding USD 1.5 billion. Postharvest disease is a significant global challenge [4]. As a hallmark of postharvest disease, latent infection of fruit involves pathogen colonization at field development stages, while symptomatic decay manifests exclusively during postharvest storage under conducive conditions [5]. Fungal pathogens are causal agents of postharvest diseases in pears, such as mold diseases caused by Penicillium spp. [6], black spot disease by Alternaria spp. [7], soft rot by Rhizopus spp. [8], gray mold by Botrytis cinerea [9], rot disease by Diaporthe spp. [10], ring spot disease by Botryosphaeria dothidea [11], and rot disease by Fusarium avenaceum [12]. To date, there have been no reports on the identification of postharvest diseases in the ‘Akizuki’ pear. Conventional morphological identification only provides a preliminary classification of pathogens. With the rapid advancement of molecular biology techniques, molecular identification methods have come to be widely used for pathogen classification. Phylogenetic analysis using concatenated sequences offers a potential solution to addressing the classification of fungal species within genera [13].
Current control methods mainly rely on fungicides, such as triazole, imidazole, and benzimidazole [14]. However, excessive application of chemical agents results in environmental pollution, food safety concerns, and the development of drug resistance [15]. Therefore, there is an urgent need to find a green control method for postharvest disease control. In this situation, chlorine dioxide (ClO_2_) treatment presents a promising alternative. As a strong oxidizing agent, ClO_2_ has shown safe and effective characteristics in disease control, and has been granted approval by the Food and Drug Administration (FDA) in U.S. for postharvest application in 2004 [16]. Importantly, almost all residual ClO_2_ in edible tissues degrades into chloride ions, minimizing residue concerns. As a result, ClO_2_ is widely used in the control of the postharvest diseases of fruits. ClO_2_ can inhibit foodborne pathogens such as Escherichia coli [17] and Listeria [18] and significantly inhibit fruit rot [19] and the development of gray mold [20,21]. In addition, ClO_2_ can also alleviate enzymatic browning reactions in fruits [22]. As for the antimicrobial mechanism of ClO_2_, it shows that ClO_2_ treatment inhibits the growth of P. digitatum by disrupting cell membrane integrity [23], and enhances the chitinase and glucanase activities of fruits, leading to damage to the cell walls of pathogenic fungi [24]. Despite the widespread application of ClO_2_ in suppressing the postharvest diseases of fruits and vegetables to extend shelf life, ClO_2_ fumigation for controlling postharvest diseases in the ‘Akizuki’ pear remains unexplored.
To address the previously uncharacterized pathogenesis of the postharvest decay of the ‘Akizuki’ pear, this study systematically isolated and identified the pathogens causing rot in ‘Akizuki’ pears during the storage period, mainly isolating Alternaria, Penicillium, and Diaporthe. The sensitivity of the most virulent strains in each genus to ClO_2_ was determined, and the antifungal mechanism was investigated with the aim of providing a basis for in-depth research on the occurrence and control of postharvest diseases in the ‘Akizuki’ pear.
2. Materials and Methods
2.1. Fruit Collection
‘Akizuki’ pear fruits at maturity stage were harvested from 4 regions including Zhao County (114.896689 E, 37.775025 W), Wei County (115.2666089 E, 36.9820674 W), Ningjin County (114.9145405 E, 37.6180744 W), and Jinzhou City (115.0341055 E, 38.0328936 W) in Hebei Province, China. Then the fruits were transported to the laboratory, and stored at 0–1 °C. During cold storage, fruits that developed diseases were selected for the isolation, purification, and identification of pathogens.
2.2. Pathogen Isolation and Purification
A small piece of tissue was cut from the area at the junction between diseased and healthy parts, and then it was disinfected with 1% (w/v) NaClO for 1 min, followed by three rinses with sterile water. The tissues were plated on potato dextrose agar (PDA) medium and cultured in the dark at 28 °C for 8 days. Mycelia from the edges of the colonies were transferred to fresh PDA for purification. This process was repeated three to four times to obtain pure strains, which were stored in test tubes at 4 °C.
2.3. Koch’s Postulates
Healthy pear fruits were selected and disinfected with 75% ethanol. The fruits were divided into 18 groups, each containing six fruits. By using a sterile punch, a small hole (5 mm in diameter) was made on the equatorial surface of each fruit. After the wound dried, a 5 mm mycelial plug was obtained using a sterile punch and inoculated into the puncture site, whereas treatments without fungal isolates served as controls. All samples were then kept in a constant humidity incubator (Shanghai PHC Health Medical Devices Co., Ltd., Shanghai, China), maintained at 25 °C with 90% humidity. The diameter of the lesions was measured on days 0, 2, 4, 6, and 8, respectively.
2.4. Morphological Identification
The purified Penicillium and Alternaria were cultured on PDA medium and the purified Diaporthe on oat agar medium (OA) according to Bastide et al. [25]. All cultures were incubated in the dark at 28 °C for 8 days. Conidia and mycelia were mounted in water and examined under an optical microscope (OLYMPUS BX 51, Tokyo, Japan). The experiment was repeated 3 times.
2.5. Molecular Biological Identification
The mycelium was collected from the media of the isolated strains. The total DNA of 18 strains was extracted using a fungal genomic DNA rapid extraction kit (Sangon Biotech, Shanghai, China). Primers ITS1/ITS4 and Bt2a/Bt2b were used to amplify the sequences of strains P1, P2, P3, and P4 [15]; primers ITS1/ITS4 and Bt2a/Bt2b were used to amplify the sequences of strains D1, D2, and D3 [17]; and primers ITS1/ITS4 and EF1-728F/EF1-986R were used to amplify the sequences of strains A1 through A11 [15]. The PCR amplification reaction mixture consisted of 25 μL of PCR Mix, 2 μL each of forward and reverse primers at 10 μmol/L, 1 μL of fungal DNA, and 20 μL of ddH_2_O to make a total volume of 50 μL. The amplification conditions were set as follows: initial denaturation at 95 °C for 5 min, followed by 34 cycles of 95 °C denaturation for 30 s, annealing at suitable temperatures for 30 s (53 °C for ITS, 52 °C for tef1, and 60 °C for β-tub), and extension at 72 °C for 30 s. Detect the PCR amplification products using 1% agarose gel. Perform electrophoresis at 120 V for 15 min, then send the DNA samples with correctly sized bands to Sangon Biotech (Shanghai) Co., Ltd. for bidirectional sequencing using the Sanger Sequencing method. A multi-gene phylogenetic tree was constructed using the amplified nucleotide sequences, with Rhizopus stolonifer designated as the outgroup and the number of bootstrap replicates was set as 1000. Using MEGA 11 software, a multi-gene phylogenetic tree was constructed employing the Neighbor-Joining (NJ) method.
2.6. Determination of the Antifungal Effect of ClO2
The susceptibility of pathogenic fungi to ClO_2_ was determined by the mycelial growth rate method. A ClO_2_ slow-release agent (Tianjin Baiduochun Technology Co., Ltd., Tianjin, China) was used to generate a constant supply of ClO_2_ in this study. The concentration of ClO_2_ was detected using the spectrophotometry measurement proposed by Masschelein [26] and then used immediately for experiments. Fungal plugs (Ø = 5 mm) were inoculated in the middle of the medium. The antifungal activity of ClO_2_ was determined using the double-dish counter method [27]. The culture medium was placed in a 28 °C dark incubator for cultivation. After continuous fumigation for 24 h, the diameter of the colonies was measured using the cross method. Each treatment was repeated three times, with three plates per replicate. The colony diameter was measured, and the inhibition rate of different concentrations of ClO_2_ was calculated as follows:
2.7. Determination of Fungicide Efficiency
The virulence of fungicides to pathogenic fungi was determined by the mycelial growth rate method according to Chen et al. [28]. Stock solutions of carbendazim, triadimefon, thiophanate-methyl, and prochloraz (Shanghai Macklin Biochemical Co., Ltd., Shanghai, China) were prepared with dimethyl sulfoxide (DMSO), and the stock solutions were diluted to prepare 100 mg/L medicated plates. Fungal plugs (Ø = 5 mm) were inoculated in the middle of the medium. The culture medium was placed in a 28 °C dark incubator for cultivation. The colony diameter was measured, and the inhibition rates of different fungicides were calculated as mentioned above. Each treatment was repeated three times.
2.8. Propidium Iodide Staining
After ClO_2_ treatment for 48 h, a small amount of mycelium was taken into a 2 mL centrifuge tube; 200 μL of 10 μg/mL propidium iodide (PI) solution was added for staining. The mixture was stained in the dark at 37 °C for 15 min, washed three times with PBS solution, and the mycelium staining was observed and photographed using a microscope (OLYMPUS BX51, Tokyo, Japan) equipped with a luciferin rhodonine filter set (OLYMPUSU-RFL-T, Tokyo, Japan) at various magnifications. The non-treated group was used as the negative control, and the fungicide was used as the positive control. The experiment was repeated three times.
2.9. Effect of ClO2 on the Control of Pathogenic Fungi in Fruits
Healthy pear fruits were selected. The surface was then washed with sterile water and subsequently disinfected using 75% alcohol. There were 6 fruits in each group. A small hole (diameter = 5 mm) was made on the equatorial surface of the ‘Akizuki’ pear using a sterile punch. After the pear wound had dried, a 5 mm mycelial plug of each strain was inoculated into the hole with a sterile punch. The samples were placed in a 16 L polypropylene light-shielding freshness maintaining box with a polyethylene bag containing different concentrations of ClO_2_ for the chlorine dioxide fumigation treatment. The concentration of ClO_2_ in the box was measured using spectrophotometry [26]. Fruit without ClO_2_ was used as a control. After continuous fumigation for 24 h, all samples were kept in a constant-humidity incubator (Shanghai PHC Health Medical Devices Co., Ltd., Shanghai, China) at 25 °C. The lesion diameter was measured using the cross method. Each treatment was repeated three times, with three fruits per replicate.
2.10. Statistical Analysis
All data were statistically analyzed using the SPSS software version 26.0 (IBM Corp, Armonk, NY, USA). A one-way analysis of variance was undertaken to evaluate the differences between treatments; those followed by different letters are statistically different according to Tukey’s multiple range test for multiple comparison. Significance was declared at p < 0.05.
3. Results
3.1. Isolation and Morphological Identification of Pathogens from ‘Akizuki’ Pear
By using the tissue separation method, a total of 18 pathogenic fungal isolates were obtained during storage and preliminarily classified into three morphological groups based on colony characteristics (Figure 1). Further, comprehensive microscopic examination of conidial and hyphal structures, coupled with colony morphology analysis, was then conducted for identification of the pathogens at the genus level (Figure 1).
Colonies of isolates D1, D2, and D3 were round and concentric with well-developed mycelia. The color of the colony is initially white and later turns to grayish white, with the production of gray-brown pigments on the surface. Microscopic analysis identified that the conidia of isolates D1, D2, and D3 were single-celled, aseptate, colorless, and transparent, which are morphological characteristics of the genus Diaporthe (Figure 1a).
Isolate P1 colonies were radiating, grayish green with raised centers and thick white margins at the edge, and the back of the medium was yellow. The conidia of isolate P1 were subglobose to ellipsoidal, single-celled, aseptate, and greenish. The colonies of isolates P2, P3, and P4 were velvety in texture, blue-green, circular with radial striations, and the reverse side was cinnamon-colored. The conidia of isolates P2, P3, and P4 were oval or round, blue-green, and smooth-walled. Based on these characteristics, isolates P1, P2, P3, and P4 were identified as belonging to the genus Penicillium (Figure 1b).
The colonies of isolates A1-A11 were flocculent with dense hyphae, which were white initially and then turned brown, and the back of the medium was cinnamon-colored or brown. The conidia were clavate, ovoid, pyriform, or elliptical with a bluntly rounded base, brown, and two to four septa were observed, which are morphological characteristics of the genus Alternaria (Figure 1c).
3.2. Pathogenicity Test of Pathogens
Strains of Diaporthe, Penicillium, and Alternaria were inoculated onto healthy pear fruits, inducing disease symptoms in all cases. After 3~5 days of cultivation, obvious disease symptoms appeared, consistent with those observed under natural conditions (Figure 1). Subsequently, the corresponding pathogenic fungi were reisolated from the diseased areas, preliminarily confirming them as the same pathogens. Analysis of lesion sizes from reinoculated fruits revealed that there were differences in pathogenicity among different genera and among different species within the same genus (Figure 2a). The genera Penicillium, Diaporthe, and Alternaria, Diaporthe exhibited the strongest pathogenicity. Within Penicillium, strain P3 showed strong pathogenicity towards ‘Akizuki’ pears, while strains P1, P2, and P4 exhibited weaker pathogenicity. In the genus Alternaria, strain A5 demonstrated strong pathogenicity, whereas strains A7 and A9 showed weaker pathogenicity (Figure 2b). All strains (D1, D2, and D3) in the genus Diaporthe exhibited strong pathogenicity. Therefore, strains P3, A5, and D1 were selected based on their pathogenicity for subsequent ClO_2_ concentration screening (Figure 2).
3.3. Molecular Biological Identification of Pathogens
The ITS* (internal transcribed spacer)*, *β-tub *(β-tubulin), and tef1 (translation elongation factor 1-α) regions were amplified from 18 strains by using universal fungal primers, resulting in ITS bands of approximately 500 bp, tef1 bands around 300 bp, and β-tub bands ranging from 500 to 750 bp (Figure 3). The evolutionary tree results showed that strains of the genus Diaporthe clustered into ten branches. Strain D1 and D. eres SXCX2-1 grouped on the same branch (Table 1), while strain D2 and D3 clustered together with D. eres CFCC 53,146 (Figure 4a). Strains P2, P3, P4, and P. expansum Aby4 grouped on the same branch with a support rate of 63% (Table 1). Strains P1 and P. citrinum NG54 clustered together with a support rate of 79% (Figure 4b). Strains A3, A4, A5, A9, A10, A11, and A. alternata AP002 grouped in the same branch, while strains A1, A2, A6, A7, A8, and A. alternata W-1 clustered on another branch (Figure 4c). Combining morphological and phylogenetic analyses, strain P1 was identified as P. citrinum; strains D1, D2, and D3 as D. eres; strains P4, P2, and P3 as P. expansum; and strains A1-A11 as A. alternata.
3.4. Effect of ClO2 on the Development of Pathogens
To evaluate the antifungal efficacy of ClO_2_, sensitivity assays were performed on P. expansum, A. alternata, and D. eres. As shown in Figure 5a, higher ClO_2_ concentrations significantly reduced lesion diameters and increased inhibition rates. It was demonstrated that ClO_2_ effectively suppresses the development of the postharvest pathogens in ‘Akizuki’ pears. Additionally, pathogens exhibited marked differential sensitivity to ClO_2_ treatment. ClO_2_ had the best inhibitory effect on A. alternata, followed by P. expansum and D. eres. ClO_2_ treatment had an inhibition rate of 17% for P. expansum at 30 mg/L and completely inhibited the production of lesions at 60 mg/L (Figure 5b). The dose–effect regression equation was y = 16.0517x − 26.0532, and the EC50 value was 41.98 mg/L (Table 2). ClO_2_ treatment had an inhibition rate of 65% for A. alternata at 30 mg/L and completely inhibited it at 90 mg/L (Figure 5b). The virulence regression equation was y = 4.8672x − 6.7795, and the EC50 value was 24.71 mg/L (Table 2). ClO_2_ treatment had an inhibition rate of 44% for D. eres at 30 mg/L and completely inhibited it at 90 mg/L (Figure 5b). The virulence regression equation was y = 6.0360x − 9.3617, and the EC50 value was 35.56 mg/L (Table 2).
Furthermore, the antifungal efficacy of ClO_2_ was compared with four commercial fungicides, namely carbendazim, triadimefon, thiophanate-methyl, and prochloraz. At 100 mg/L, all four of the above fungicides significantly inhibited the mycelial growth of P. expansum, A. alternata, and D. eres, whereas ClO_2_ attained complete suppression (100%) at 90 mg/L. This further indicated that ClO_2_ fumigation serves as an effective measure for controlling postharvest diseases in the ‘Akizuki’ pear.
3.5. Effect of ClO2 on the Cell Membrane of Pathogens
The impact of ClO_2_ on hyphal morphology and membrane integrity was investigated in P. expansum, A. alternata, and D. eres to elucidate its antifungal mechanism. Based on in vitro experimental results, it was observed that the antifungal activity against pathogenic fungi significantly increased when the concentration of ClO_2_ reached a certain level. A concentration of 45 mg/L ClO_2_ was selected for subsequent experiments to prevent phytotoxicity in ‘Akizuki’ pears while maintaining significant antifungal efficacy. Observation of the hyphal morphology of P. expansum, A. alternata, and D. eres by using ultra-depth-of-field microscopy showed that the color of P. expansum hyphae changed from green to white, and hyphal growth was inhibited. The color of A. alternata and D. eres hyphae changed significantly, with the inhibition of D. eres hyphae being the most obvious (Figure 6a). This suggested that ClO_2_ significantly disrupted hyphal morphology in P. expansum, A. alternata, and D. eres, causing abnormal growth and reducing infectivity. Propidium iodide (PI) is a nucleic acid dye that can pass through damaged cell membranes and bind to DNA to form a stable red fluorescent complex. PI staining of hyphae exposed to 45 mg/L ClO_2_ demonstrated red fluorescence in all pathogens, indicating nucleic acid leakage through compromised membranes. D. eres displayed the strongest fluorescence intensity (Figure 6b), confirming severe membrane damage. These results revealed that the antifungal mechanism of ClO_2_ involves the disruption of hyphal structure and membrane integrity.
3.6. Effect of ClO2 on the Disease Incidence of ‘Akizuki’ Pear Fruits
The pathogenicity experiment confirmed the efficacy of ClO_2_ in suppressing postharvest rot in ‘Akizuki’ pears in vivo. Fumigation with 45 mg/L ClO_2_ significantly inhibited disease progression in pear fruits inoculated with A. alternata, P. expansum, and D. eres. Compared to untreated fruits, the results showed that the lesion diameters of ClO_2_-treated fruits were significantly reduced. The lesion diameter of P. expansum was inhibited by 10 mm, A. alternata by 18 mm, and D. eres by 17 mm (Figure 7a). The disease incidence of ClO_2_-treated pear fruits was reduced by 54%, 73%, and 34% compared with the control group (Figure 7b). The results showed that ClO_2_ fumigation effectively inhibited the occurrence of postharvest diseases of ‘Akizuki’ pear.
4. Discussion
Currently, research on the ‘Akizuki’ pear primarily focuses on non-infectious diseases, particularly cork spot, a physiological disorder. For instance, Duan et al. investigated the physiology of cork spot in the ‘Akizuki’ pear by comparing mineral elements and physiological indicators between diseased and healthy fruits [29], and Yang et al. analyzed the mechanisms of cork spot in ‘Akizuki’ pears through combined mineral and metabolomic approaches [30]. In contrast, studies on infectious diseases in the ‘Akizuki’ pear remain limited. This study employed a combination of morphological analysis, multi-gene phylogenetic analysis, and pathogenicity assessment to identify the pathogens, thereby elucidating the primary pathogens involved in the rotting of ‘Akizuki’ pears during storage, specifically within the genera Alternaria, Penicillium, and Diaporthe. Subsequently, effective ClO_2_ concentrations against these pathogens were systematically screened.
Our study found that D. eres, A. alternata, and P. expansum are the most important pathogenic fungi causing postharvest rot diseases in ‘Akizuki’ pears. D. eres is known to cause severe fruit rot in pears [31] and diverse phytopathologies including black rot in persimmons [32], root rot in Coptis chinensis [33], and postharvest rot in peaches [34]. It is well known that A. alternata is a significant postharvest pathogen. The black spot disease, caused by A. alternata in pears, not only significantly shortens fruit shelf life but also poses serious food safety risks due to mycotoxin contamination. Furthermore, A. alternata demonstrates widely phytopathogenicity, as evidenced by its causation of black rot in cherries [35], leaf spot in passion fruits [36], and scab in kiwifruits [37]. P. expansum causes characteristic blue mold decay of fruits across multiple hosts, including kiwifruits [38], apples [39], pears [40], and citruses [41], making it one of the key postharvest diseases. All the pears in our study were collected from Hebei Province, which is one of the major production areas for ‘Akizuki’ pears. However, the identification of postharvest diseases in other major ‘Akizuki’ pear-producing regions in China still requires further research.
In this study, gaseous ClO_2_ fumigation demonstrated concentration-dependent antifungal effects against postharvest pathogens P. expansum, A. alternata, and D. eres in both culture media (in vitro) and fruits (in vivo) (Figure 5a). Complete inhibition was achieved at 60 mg/L for P. expansum and 90 mg/L for A. alternata and D. eres (Figure 5b). These concentrations align with Lee et al.’s findings where 20 mg/L gaseous ClO_2_ completely suppressed Diaporthe batatas [42], demonstrating the consistent efficacy of gas-phase treatment. Crucially, efficacy varied substantially across application methods. Semi-permeable film packaging with ClO_2_ slurry achieved complete A. alternata control at 10 mg/L in tomatoes [43]. Solid formulations required drastically higher doses than gaseous fumigation. Specifically, effective concentrations reached 400 mg/L for P. expansum [23] and 250 mg/L for P. digitatum [44]. This dosage reduction underscores the superior efficiency of gaseous fumigation, being attributable to enhanced gas diffusion and sustained biocidal contact. Notably, comparing the same concentration of ClO_2_ with four commonly used fungicides, such as carbendazim, thiophanate-methyl, prochloraz, and triadimefon, the antifungal effect in vitro was similar. The results are consistent with the study by Hatamzadeh et al. [45], who found comparable antifungal effects in vivo and in vitro when comparing ClO_2_ to four commercial fungicides (thifluzamide, imazalil, cyprodinil, and thiophanate-methyl) at 1000 mg/L. Zhang et al. found that treatment with 60 mg/L ClO_2_ can significantly enhance the antioxidant capacity of cherry fruits while effectively delaying the process of quality deterioration [46]. Since the 45 mg/L concentration adopted herein is significantly lower than the above-mentioned safe and effective 60 mg/L, ClO_2_ fumigation will not cause harm to the fruits. Most significantly, gaseous ClO_2_ treatment reduced the disease severity in pear fruits, demonstrating its practical value as an efficient antimicrobial agent (Figure 7a).
The antifungal mechanism of ClO_2_ is currently mainly divided into two categories—by increasing the activity of defense enzymes, and by directly destroying the cell membrane of pathogens to inhibit the development of the postharvest diseases. It is important to note that gaseous ClO_2_ is unstable and sensitive to light and heat, which limits its application [47]. In order to promote the development of ClO_2_ in the fruit preservation industry, studies have explored the effect of different packaging containing chlorine dioxide on the storage quality of grapes and found that ClO_2_ could effectively reduce the respiration rate and significantly increase the total phenols, flavonoids, and anthocyanins compared with the control group [48]. Sun et al. found that ClO_2_ microcapsule treatment could effectively inhibit Listeria and E. coli and delay the decline in quality during the storage period of blueberries [49]. However, few studies have explored the combination of ClO_2_ with other preservation methods through different packaging approaches to control postharvest diseases. Therefore, it is necessary to further study the combined application of ClO_2_ and other preservation technologies through different packaging strategies.
5. Conclusions
This study presents the first systematic identification of pathogenic fungi causing postharvest decay in ‘Akizuki’ pears. The 18 pathogenic fungal strains were identified as P. expansum, P. citrinum, D. eres, and A. alternata based on morphological characterization, pathogenicity testing, and multi-locus phylogenetic analysis. Subsequent evaluation of ClO_2_ efficacy demonstrated its potent antifungal activity against the key pathogens P. expansum, A. alternata, and D. eres. The antifungal mechanism involved the disruption of hyphal morphology and damage to cell membrane integrity. Importantly, ClO_2_ exhibits improved antifungal efficacy comparable to that of conventional fungicides. These findings provide crucial insights into the etiology of postharvest decay in ‘Akizuki’ pears and establish gaseous ClO_2_ fumigation as a promising, effective, and eco-friendly strategy for controlling these diseases.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Li L. Zhang H.H. Wu L. Gu S.F. Xun J. Jia B. Ye Z.F. Heng W. Jin X. An early asymptomatic diagnosis method for cork spot disorder in ‘Akizuki’ pear (Pyrus pyrifolia Nakai) using micro near infrared spectroscopy Food Chem. X 20231910085110.1016/j.fochx.2023.10085137780255 PMC 10534216 · doi ↗ · pubmed ↗
- 2Xiong J.H. Gu S.F. Rao Y. Liu L. Zhang X.D. Wu Y.T. Jin X. A multi-source feature stable learning method for rapid identification of cork spot disorder in ‘Akizuki’ pear Postharvest Biol. Technol.202521911328510.1016/j.postharvbio.2024.113285 · doi ↗
- 3Oyom W. Li Y.C. Prusky D. Zhang Z. Bi Y. Tahergorabi R. Recent advances in postharvest technology of Asia pears fungi disease control: A review Physiol. Mol. Plant Pathol.202211710177110.1016/j.pmpp.2021.101771 · doi ↗
- 4Sardella D. Muscat A. Brincat J.P. Gatt R. Decelis S. Valdramidis V. A comprehensive review of the pear fungal diseases Int. J. Fruit Sci.20161635137710.1080/15538362.2016.1178621 · doi ↗
- 5Zhang J.X. Timmer L.W. Preharvest application of fungicides for postharvest disease control on early season tangerine hybrids in Florida Crop Prot.20072688689310.1016/j.cropro.2006.08.007 · doi ↗
- 6Khokhar I. Jia Y. Mukhtar I. Wang J.H. Yan Y.C. First report of Penicillium polonicum causing blue mold on stored pear (Pyrus bretschneideri) fruits in China Plant Dis.2019103327910.1094/PDIS-05-19-1041-PDN · doi ↗
- 7De Shields J.B. Kc A.N. Morphological and molecular characterization of Alternaria spp. isolated from European pears Plant Dis.20211052531254010.1094/PDIS-10-20-2143-RE 33434041 · doi ↗ · pubmed ↗
- 8Kwon J.H. Lee C.J. Rhizopus soft rot on pear (Pyrus serotina) caused by Rhizopus stoloniferin Korea Mycobiology 20063415115310.4489/MYCO.2006.34.3.15124039490 PMC 3769565 · doi ↗ · pubmed ↗