PDE4-Selective Inhibition in Chronic Obstructive Pulmonary Disease and Pulmonary Fibrosis: Different Agents or Different Targets?
Graeme B. Bolger

TL;DR
This review explores how PDE4 inhibitors like roflumilast and nerandomilast work in different lung diseases, suggesting their effects depend on disease mechanisms rather than drug differences.
Contribution
The paper proposes that differences in therapeutic outcomes of PDE4 inhibitors are due to disease-specific pathogenesis, not drug-specific effects.
Findings
PDE4 inhibitors show clinical benefits in both obstructive and restrictive lung diseases.
The drugs affect multiple pulmonary cell types, with only a subset dysregulated in COPD or IPF.
Observed differences in drug efficacy likely reflect disease pathogenesis rather than drug mechanism differences.
Abstract
Highly selective inhibitors of the members of the cAMP-selective cyclic nucleotide phosphodiesterases, or PDE4 family, have shown clinically meaningful activity in two different classes of lung disease: roflumilast in obstructive lung disease, specifically chronic obstructive pulmonary disease (COPD), and nerandomilast in restrictive lung diseases characterized by inflammation/fibrosis of the alveolar interstitium, including idiopathic pulmonary fibrosis (IPF) and progressive pulmonary fibrosis (PPF). The beneficial therapeutic benefit of these agents in both of these disorders suggests that they share a common mechanism that underlies their effects on different pulmonary cells and tissues. This review outlines the biochemical, pharmacologic and cellular effects of PDE4-selective inhibitors, emphasizing their role in signal transduction pathways common to many pulmonary cell types. It…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
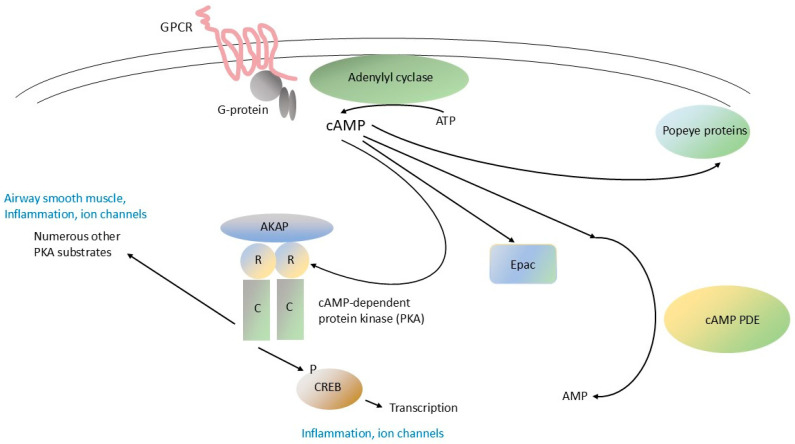 Figure 1
Figure 1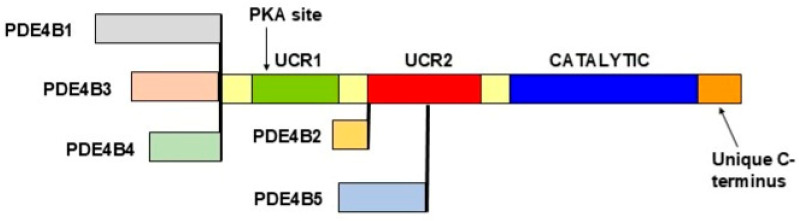 Figure 2
Figure 2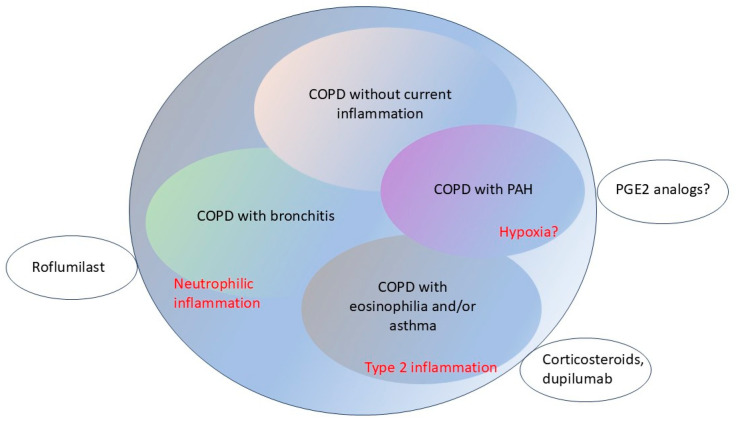 Figure 3
Figure 3- —BZI Pharma LLC
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhosphodiesterase function and regulation · Interstitial Lung Diseases and Idiopathic Pulmonary Fibrosis · Asthma and respiratory diseases
1. Introduction
Highly selective inhibitors of the cAMP-specific phosphodiesterases, or PDE4 enzymes, have been studied intensively in numerous, rigorous clinical trials and are currently available for clinical use in a wide variety of human diseases, where they have potent immunomodulatory and anti-inflammatory effects (Table 1; refs. [1,2]). PDE4-selective inhibitors have been conspicuously tested in a range of pulmonary disorders, including chronic obstructive pulmonary disease (COPD) and pulmonary fibrosis, where much of their clinical efficacy reflects their anti-inflammatory actions [3,4,5,6,7]. PDE4-selective inhibitors also have numerous other effects in the lung, including the modulation of ion channel activity ([8], see ref. [9] for a review) and potential effects on the modulation of airway smooth muscle tone.
The expansion of the PDE4-selective toolbox in human diseases, especially in the lung, currently reveals an interesting paradox. Many of the currently available PDE4-selective inhibitors have overlapping, indeed sometimes indistinguishable, enzyme-inhibitory profiles, as described in more detail below. However, review of the current clinical indications for these drugs shows that each is currently indicated for a narrow range of disorders that typically has minimal, if any, overlap with the indications for the other available drugs in this class (Table 1).
This review will discuss to what extent these therapeutic differences reflect differences in the mechanistic effects of these drugs, with special emphasis on pulmonary cells and tissues. It will start with a review of the clinical and biologic aspects of COPD and pulmonary fibrosis, including recent attempts to define sub-classes of these disorders that would guide therapeutic management. It will then discuss pre-clinical models of each of the disorders in an attempt to characterize their pathophysiology. With this foundation in place, it will review the biochemical and cellular mechanisms of action of PDE4 inhibitors in these disorders. It will then turn directly to clinical issues, including completed and ongoing clinical trials of these agents. Finally, it will offer a perspective on biomarker development in relation to both clinical practice and the next generation of clinical trials.
2. COPD and Pulmonary Fibrosis: Similarities and Differences in Clinical Presentations and Therapy
Although PDE4 inhibition has therapeutic value in both COPD and fibrotic lung disease, these two classes of lung disorders differ profoundly in their pathophysiology and clinical manifestations.
2.1. COPD—Definition and Sub-Classes
According to the 2025 report from the Global Initiative for Chronic Obstructive Lung Disease (“GOLD”), COPD is defined by chronic respiratory symptoms caused by airway abnormalities that produce persistent and frequently progressive airflow obstruction [21]. The diagnosis is confirmed by the demonstration of non-fully reversible airflow obstruction on spirometry [21]. Patients typically complain of dyspnea and/or cough. Examination of lung tissue from patients with COPD shows small-airway narrowing (bronchiolar obstruction), destruction of alveolae and their replacement with large air-filled cavities (emphysema), increased mucus production, inflammation and fibrosis [22]. These pathologic findings are consistent with inflammation in COPD leading to tissue destruction; in contrast, in pulmonary fibrosis, inflammation leads to abnormal cellular proliferation.
The heterogeneity of COPD, even for patients whose disease clearly conforms to the GOLD criteria, has stimulated attempts to divide the disease into sub-types. The 2025 GOLD report proposes a taxonomy based on differences in etiology (“etiotypes”), including, for example, genetically determined causes, or environmental causes (smoking, pollution), or prior infections or asthma [21]. Alternatively, patients can be subdivided into groups on the basis of clinical findings. Such a clinically based classification could identify patients with a predominantly bronchitic presentation (i.e., the regular production of sputum for several months each year [23]), or, alternatively, those presenting predominantly with emphysema (airway obstruction and structural lung lesions on imaging). An increased level of circulating eosinophils has been used to define a subtype of COPD that has a strong allergic/inflammatory component and whose pathophysiology that overlaps that of asthma; such patients do not necessarily have the reversible airway obstruction characteristic of asthma, but typically benefit from some of the anti-inflammatory strategies that help patients with asthma, such as inhaled corticosteroids [24,25,26,27,28,29].
Extensive clinical study has shown the value of roflumilast and several other PDE4-selective inhibitors, such as cilomilast, in COPD, even though they do not have significant acute bronchodilator activity (reviewed in Refs. [30,31]). Roflumilast has shown to reduce the frequency of COPD exacerbations, especially in patients with severe COPD who have chronic bronchitis and/or are repeatedly hospitalized for COPD [5,10,11,12,32]. Clinical trials have shown roflumilast to be of value in patients who cannot tolerate inhaled corticosteroids or who are not well-controlled on inhaled LABA/LAMA/steroid combinations [12,31]. Nausea, diarrhea, and changes in mood and behavior, all typical class side effects of PDE4 inhibitors, lead to its discontinuation in some patients [5,30].
The development of ensifentrine, the first PDE inhibitor that can be administered by nebulization, is an important advance in the therapy of COPD, especially for patients presenting with COPD exacerbations [17,33,34,35,36,37]. Ensifentrine has activity against both PDE3 and PDE4 [38]; see ref. [9] for a review. A slightly older inhaled PDE4 inhibitor, tanimilast, has also shown activity in several clinical trials (see ref. [9] for a review) but has not been approved for clinical use. Unlike roflumilast or nerandomilast, ensifentrine has significant acute bronchodilator activity, which contributes substantially to its therapeutic action.
Patients with COPD and eosinophilia have been shown to benefit from monoclonal antibodies that target elements of the immune system, such as dupilumab [39,40,41], or, very recently, mepolizumab [42,43]; see refs. [44,45] for reviews. Dupilumab blocks a shared component of the receptors for interleukin-4 and interleukin-13 (IL-4 and IL-13; refs [46,47]), while mepolizumab targets interleukin-5 (IL-5; refs [43,48]). IL-4, IL-5 and IL-13 are key drivers of Type 2 inflammation, key to the pathogenesis of many disorders associated with eosinophilia ([49,50]; see ref. [51] for a review).
As the number of potential therapeutic options expands in COPD, there has been a pressing need to develop better classification systems that would inform clinical practice and the design of the next generation of clinical trials (see ref. [9] for a review). For example, patients with a bronchitic phenotype would seem to benefit from inhaled β2-adrenergic agonists and muscarinic antagonists. They will also generally benefit from roflumilast and/or ensifentrine. Patients with eosinophilia will benefit from the addition of dupilumab or mepolizumab to their ongoing inhaled corticosteroids. There remains a large number of COPD patients, typically those predominantly with emphysema, who lack therapeutic options beyond inhaled muscarinic antagonists. Further research on the pathobiology of COPD may yield additional prognostic/classification criteria, especially additional biomarkers, as discussed in greater detail below.
2.2. Pulmonary Fibrosis—Definition and Sub-Classes
Pulmonary fibrosis can be defined as the development of pulmonary lesions characterized by fibroblast proliferation in the alveolar interstitium that is associated with progressive pulmonary dysfunction [52]. Idiopathic pulmonary fibrosis (IPF), i.e., fibrosis that is not associated with an underlying medical disorder, is a subset of interstitial lung disease (ILD), a broader class of pulmonary disorders that is characterized by inflammation and/or fibrosis of the lung parenchyma [53]. Progressive pulmonary fibrosis (PPF) describes a fibrosing ILD that arises in pre-existing pulmonary disease, such as auto-immune ILD, hypersensitivity pneumonitis, or other forms of ILD [18]. Typical auto-immune disorders leading to PPF are scleroderma, rheumatoid arthritis with systemic manifestations, and other rheumatologic conditions [54]. PPF can be produced by exposure to numerous drugs, including amiodarone and several anti-neoplastic agents, especially bleomycin [55]. Finally, there is a well-validated association between germline variants (single nucleotide polymorphisms, or SNPs) in the telomerase genes TERT and RTEL1, as well as weaker associations with some other loci, and the development of IPF and other fibrosing diseases [56,57,58,59]. Closely related to IPF and PPF are pulmonary diseases associated with exposure to particulates, such as asbestosis [60] and silicosis [61,62]. Although there is yet no formal classification of pulmonary fibrosis that includes all of the above conditions, current clinical practice includes many of the above factors in decision-making, and, more recently, in the design of clinical trials [18,19,52].
Two agents are currently available for the treatment of pulmonary fibrosis; on the basis of recently completed phase 3 clinical trials, another two are likely to be approved very shortly (Table 2).
Pirfenidone, the first agent to be developed specifically for fibrotic lung disease, has been shown on the basis of multiple phase 3 trials to be safe and effective in both IPF and PPF [68,69,70,71]. The biochemical target of pirfenidone has been the subject of considerable research; it is clearly not a PDE4-selective inhibitor (see references in the pre-clinical sections below). Nintedanib, which targets several protein-tyrosine kinases, most convincingly the receptors for vascular endothelial growth factor (VEGF), platelet-derived growth factor (PDGF), and fibroblast growth factor (FGF), has also been shown on the basis of multiple phase 3 trials to be safe and effective in both IPF and PPF [63,64,65,66]. Combination therapy with both of these agents appears to have additive benefit [72,73,74].
The PDE4B-selective inhibitor nerandomilast has been shown on the basis of two phase 3 trials to be safe and effective in both IPF and PPF [19,67]. Head-to-head comparisons of nerandomilast with older agents in fibrotic lung disease have yet to be reported, but the magnitude of benefit seen with nerandomilast in these two phase 3 trials appears to be as good as, or possibly better than, that of the older drugs [75,76,77].
Several recent phase 3 clinical trials, some of which have yet to be published in full, have demonstrated activity of the prostaglandin analog treprostinil in patients with IPF [78,79,80,81,82]. These trials have tested treprostinil in at least two different formulations as a dry powder (i.e., as a metered-dose inhaler) and, more recently, in an oral form. This drug has been available for some time for the treatment of pulmonary arterial hypertension (PAH); the newer study studies its effects in patients with both PAH and IPF. Treprostinil acts primarily as a vasodilator; its benefits in IPF may be due to its vasodilatory properties. However, given the key role of prostaglandins in the pathogenesis of IPF (see below), it may also have beneficial effects on the underlying disease process.
Given the considerable differences between COPD and the fibrotic lung diseases, it is perhaps surprising that both these classes of disorders can benefit from treatment with PDE4-selective inhibitors. To determine what aspects of PDE4 inhibition might be responsible for this common effect, it is useful to review the enzymology, pharmacology and cellular regulation of the PDE4s and recent advances in PDE4 inhibitor pharmacology.
3. Molecular Pharmacology of the PDE4 Family: Genes, mRNAs, and Proteins
3.1. cAMP Signaling
Signal transduction pathways modulated by the prototypical small molecule cyclic nucleotide “second messenger” 3′, 5′ cyclic adenosine monophosphate (cAMP) are ubiquitous in cells and tissues. cAMP signaling plays a critical role in the function of diverse organs, including the lung, the CNS, inflammation and immunity, and the endocrine system. cAMP is synthesized by membrane-associated adenylyl cyclase in response to extracellular stimuli, and then diffuses throughout the cell, where it interacts with specific downstream effector proteins (Figure 1, ref. [3]). G-protein-coupled receptors (GPCRs), which regulate membrane-associated adenylyl cyclase through trimeric GTP-binding proteins (G-proteins) are among the most important regulators of adenylyl cyclase and thereby cAMP signaling generally. Also essential to the regulation of cAMP signaling are cyclic nucleotide phosphodiesterases (PDEs), which are enzymes that hydrolyze (degrade) cAMP (and/or 3′, 5′ cyclic guanosine monophosphate, or cGMP) and thereby modulate its levels in cells [3,4,7,83]. Downstream effectors of cAMP action in cells include cAMP-specific protein kinase (protein kinase A, PKA), cyclic nucleotide-gated ion channels, exchange proteins activated by cAMP (EPACs), and Popeye proteins [84,85,86,87,88]. These various pathway components also mediate “cross-talk” between cAMP signaling and other cellular pathways, particularly the MAPK pathway. Both synthesis and breakdown of cAMP can be highly localized in cells, producing “compartments”, “pools” or “gradients” where its concentration is tightly regulated in space and time [89,90,91]. The overall process is highly dynamic, with short- and long-term feedback loops that adjust the “gain” of various components of the pathways and increase their versatility and range of response.
3.2. The PDE Superfamily
The ability of the cAMP-specific phosphodiesterases, or PDE4 enzymes, to hydrolyze cAMP is key to their role in regulating cAMP signaling pathways in cells. The PDE4s are members of the larger cyclic nucleotide phosphodiesterase superfamily, whose members consist of a total of 11 families, encoded by a total of 21 genes [1,3,83]. The various members of the PDE superfamily are defined on the basis of their substrate specificity (cAMP, cGMP, or dual-specificity for both cAMP and cGMP) and their ability to be inhibited by pharmacologic inhibitors specific to each family. PDE3, PDE4, PDE7 and PDE8 are cAMP-selective; while PDE1, PDE2, PDE10 and PDE11 are dual-specific [3,4,7,83]. The primary amino acid sequence and fold of the catalytic regions of each family determine, at least in part, the substrate specificity of each family member and its ability to be selectively inhibited by family-specific inhibitors [83,92,93]. Consistent with this observation, PDE-selective inhibitors act at the catalytic sites of the PDE enzymes and therefore act, at least in part, as competitive inhibitors of cAMP and/or cGMP hydrolysis. To date, PDE4 and possibly PDE3 have been shown to be pharmacologically relevant targets in COPD (see references above and refs. [1,9] for reviews) and fibrotic lung disease (see references above). PDE5 inhibitors modulate nitric oxide—cGMP signaling and have been shown to be safe and effective in the treatment of PAH. The pharmacologic potential of other PDE families in pulmonary disorders has yet to be determined.
3.3. PDE4 Multiplicity: Genes, mRNAs and Proteins
Key to understanding the effects of PDE4 inhibitors in tissues and organs is the incredible diversity of PDE4 isoforms. There are over 20 PDE4 isoforms, which are encoded by 4 genes in mammals (PDE4A, PDE4B, PDE4C and PDE4D in humans), with additional diversity being produced by alternative mRNA splicing and the use of alternative promoters for each isoform [94,95]. Specifically, the human PDE4B gene encodes 5 different isoforms (Figure 2; see ref. [96] for a review) and the human PDE4D gene encodes 9 different isoforms (see ref. [97] for a review). The mRNA, and corresponding protein, for each isoform has a distinct pattern of expression in tissues, suggesting that each has a distinct tissue or organismal function; these cellular and regional differences in expression has been explored most carefully in the central nervous system (see refs. [96,97] for a review), but is highly likely to be the case in other organs, including the lung.
PDE4 proteins can consist of up to three domains, each of which has a distinct primary sequence and fold. The catalytic region contains all elements necessary for cAMP hydrolysis, while the UCR1 and UCR2 regions are essential for dimerization and serve to regulate the activity of the catalytic domain (Figure 2, refs. [94,98]). “Long” PDE4 isoforms contain UCR1, UCR2 and the catalytic domain, while “short” isoforms contain UCR2 and the catalytic domain, and “super-short” isoforms contain a portion of UCR2 and the catalytic domain. The catalytic sites of the various PDE4 isoforms are extremely similar (approximately 90% primary amino acid sequence homology, refs. [94,95]); until relatively recently, this close homology complicated the development of inhibitors selective for the isoform(s) encoded by any specific PDE4 gene.
3.4. Regulation of the PDE4 Proteins
The various PDE4 isoforms are regulated by a complex and interacting network of phosphorylation events and interactions with other signaling proteins (see refs. [1,4,95]). Numerous proteins have been shown to interact with specific PDE4 isoforms: for example, the DISC1 protein interacts specifically with long PDE4B isoforms, most notably PDE4B1 [99], while the RACK1 protein and β-arrestin2 both interact specifically with PDE4D5 [100,101]. The subcellular localization of numerous PDE4 isoforms is determined by their interaction with scaffold and anchoring proteins, including A-kinase anchoring proteins, or AKAPs [102]. These and other protein–protein interactions can determine the subcellular localization of various PDE4 isoforms, their ability to undergo phosphorylation (see next paragraph), and their interactions with other signal transduction components [1,4,7].
Among the most important mechanisms for the regulation of PDE4 isoforms is their ability to be phosphorylated by several physiologically important kinases. Long PDE4 isoforms are phosphorylated by PKA at a specific serine located at the N-terminal end of UCR1 (Figure 2); PKA phosphorylation serves to inhibit the activity (velocity) of the enzyme [103,104]. PKA phosphorylation also is essential to the dimerization of long PDE4 isoforms, which may be essential for enzyme inhibition [98]. Through dimerization or potentially other mechanisms, PKA phosphorylation changes the conformation of the catalytic region, as manifested by changes in the susceptibility of long PDE4 isoforms to PDE4-selective inhibitors [95,103]. Germline mutations in the human PDE4D gene that localize to UCR1 and UCR2 and which affect dimerization and PKA regulation of long PDE4D isoforms cause acrodysostosis, a debilitating disorder that affects bone formation and the brain (ref. [105]; see ref. [83] for a review). PDE4 isoforms are also phosphorylated by ERK1/2 and undergo numerous other forms of secondary modification; determining the physiological significance of many of these events remains an active area of investigation [106,107].
3.5. Structure and Regulation of the PDE4 Proteins: Relevance to Roflumilast and Nerandomilast
Given our basic knowledge of the structure and regulation of the PDE4 enzymes, what insights can this knowledge provide on the actions of roflumilast and nerandomilast? The available pre-clinical data does suggest that there is a difference in the selectivity of these two drugs. Roflumilast and its active metabolite, roflumilast N-oxide, are highly specific for PDE4 (as opposed to other PDE families), but inhibit all PDE4 isoforms relatively equally (i.e., IC_50_ values that vary from 0.2 to 7.8 nM, depending on the PDE4 isoform, ref. [8]). In contrast, the values provided for nerandomilast show it to be relatively specific for PDE4B (IC_50_ values of 10 nM for PDE4B, 91 nM for PDE4D, 248 nM for PDE4A, and 8700 nM for PDE4C, ref. [108]. However, it should be noted that the enzyme preparations used in the reported nerandomilast assays consisted of PDE4B and PDE4D active site fragments (i.e., containing just the core catalytic domain (Figure 2)), while those for PDE4A appeared to include UCR1, UCR2 and the catalytic region, and those for PDE4C apparently included full-length protein (PDE4C2; ref. [108]). Given that UCR1 and UCR2 have important effects on the activity of the PDE4s, including their ability to dimerize and be phosphorylated by PKA and other kinases, it is quite possible that different values for specificity would have been obtained if full-length isoforms had been compared [109]. More generally, in the future, it will be of considerable value if comparative IC_50_ values are obtained on correspondingly similar enzymes (e.g., by comparing the long isoforms encoded by each of the 4 different PDE4 genes, OR by comparing just the isolated catalytic domain for isoforms encoded by the 4 different PDE4 genes). Finally, given the important effects of phosphorylation on the activity of PDE4s, and on their ability to be inhibited by PDE4-selective inhibitors, it would be essential to examine/compare phosphorylated and non-phosphorylated long enzyme preparations.
Given these insights into how roflumilast and nerandomilast act on the enzymatic aspects of the PDE4 enzymes, with special emphasis on the PDE4B enzymes, it is of particular interest how they might influence the cellular and tissue effects of these drugs. This will be explored in the next two sections.
4. Defining the Cellular Targets of PDE4 Inhibitors: Insights from Single-Cell Sequencing
Advances in single-cell RNA sequencing have greatly enlarged knowledge of the various cell types present in the lung, and in turn have increased the precision in determining the site of action of PDE4-selective inhibitors. Single-cell sequencing of human and murine tracheal epithelium has demonstrated the presence of many new cell types (58 in all), many of which were previously not previously identified [110]. Among the new cell types identified by these studies are pulmonary ionocytes [111,112], which are highly enriched in the cystic fibrosis transmembrane regulator (CFTR) protein. CFTR is a Cl^−^ ion channel that is regulated, at least in part, by cAMP-signaling pathways, which modulate the phosphorylation of the CFTR regulatory domain by PKA. Several laboratories, including our own, have shown that PKA-mediated phosphorylation of CFTR is in turn regulated by PDE4 (see ref. [9] for a review). More generally, the expanded knowledge of the cell types, and the gene-expression pattern in each of these cell types, in the lung has the potential to greatly increase our ability to determine which cell types in the lung are the targets of pharmacological intervention. We apply these new concepts to PDE4 action in the lung in the next sections.
5. Pre-Clinical Models of COPD and Fibrotic Lung Disease: Clues to the Mechanisms of PDE4 Action in Lung Diseases
5.1. Pre-Clinical Models of COPD Demonstrate Multiple Beneficial Effects of PDE4 Inhibition in COPD
Extensive clinical study has shown the value of roflumilast and several other PDE4-selective inhibitors, such as cilomilast, in COPD. PDE4 inhibitors do not have significant acute bronchodilator activity in COPD ([113], see ref. [8] for a review). However, they have myriad effects on pulmonary cells and tissues that collectively account for their therapeutic benefit.
5.2. Role of PDE4 in Pulmonary Inflammation
Decades of pre-clinical studies have demonstrated that PDE4-selective inhibitors have activity in various immune/inflammatory pathways in lung tissue. The extensive data for first-generation PDE4-selective inhibitors has been summarized in older reviews [95]. Many of the older studies focused on models pertinent to allergy/asthma that are of uncertain relevance to COPD. Roflumilast and its active metabolite, roflumilast N-oxide, affects the functions of many cell types, including CD4+ and CD8+ T-cells, monocytes/macrophages and neutrophils [8]. In particular, the action of roflumilast against neutrophils probably explains much of its activity in COPD, as neutrophilic airway inflammation is a hallmark of COPD, as described below. In contrast, the monoclonal antibodies dupilumab and mepolizumab appear to act on inflammatory processes mediated, at least in part, by eosinophils [43,46,47,48,49,50,51]. The importance of neutrophil inflammation in COPD also appears to explain much of the pathogenesis of the disease, specifically the role of neutrophil-derived proteases, especially matrix metallopeptidases, and reactive oxygen species, in the destruction of lung tissue typically of COPD.
5.3. Other Actions of Roflumilast in the Lung
Roflumilast has actions on numerous non-immune/inflammatory cells in the lung, including airway and vascular smooth muscle, endothelial cells, and respiratory epithelium. Its actions on vascular smooth muscle cells probably account for its ability to prevent or ameliorate pulmonary hypertension in several rodent models [8] and potentially in humans. Roflumilast also has potent effects on fibroblasts, as discussed in detail below. Roflumilast is a potent stimulator of the CFTR Cl^−^ ion channel, which is expressed in pulmonary epithelial cells and ionocytes and which is essential for mucociliary clearance and the maintenance of airway surface liquid (see ref. [9] for a review). Collectively, these immune and non-immune effects of roflumilast are likely to account for its beneficial action in mouse models of COPD, and in clinical use.
5.4. Actions of Roflumilast in Animal Models of COPD
Extensive study of the effects of roflumilast in rodent models of COPD have provided insights into its cellular, tissue and whole-organ benefits in human COPD. The rodent models used in these experiments have a number of limitations and certainly do not replicate all aspects of human COPD. The most-commonly used model is the treatment of rodents, typically mice, with cigarette smoke for a few hours or days; this model clearly differs substantially from COPD in humans, which typically progresses over many years. A variant animal model, incorporating smoke exposure for several months, would appear to be more appropriate, but even the mice in this model develop only the early stages of COPD (GOLD stages I and II; see ref. [8] for a review). However, these models show clearly that smoke exposure produces primarily a neutrophilic inflammatory response [8]; this may account for the beneficial effects of roflumilast and other PDE4-selective inhibitors in these models. In contrast, corticosteroids, which have beneficial effects in many forms of airway inflammation, especially in asthma [51], are ineffective in these models. Although additional refinement of these models would be useful, they provide a useful platform for the testing of compounds that can be moved into clinical trials. Further study of these models also has the potential to accelerate biomarker development, which is a key unmet clinical need in COPD.
5.5. Pre-Clinical Models of Fibrotic Lung Disease and the Role of PDE4 Inhibition
Key to understanding fibrotic lung disease, including IPF and PPF, is the proliferation of fibroblasts, which form the cellular backbone of the lesions seen in these disorders. Fibroblasts also synthesize and secrete extracellular molecules, such as collagen and other components of the extra-cellular matrix, which are essential components of these lesions. Although fibroblast proliferation in disease can be the end-result of various stimuli, it is generally agreed that inflammation is a key contributing factor and that fibroblast proliferation mediated by the production of various growth factors and cytokines by inflammatory cells, especially neutrophils, is a hallmark of PPF and IPF. PDE4 inhibitors can intervene at several steps in this process.
5.6. PDE4s and Fibroblast Proliferation in the Human Lung
Roflumilast, nerandomilast, and other PDE4-selective inhibitors have anti-fibrotic activity in several cell-based models of inflammation-mediated fibrosis. A commonly used model assesses the deformation of collagen gels in in-vitro culture, where fibroblast proliferation produces complex three-dimensional structures. Roflumilast can slow fibroblast growth and attenuate the development of these structures in these cultures. In these models, roflumilast acts synergistically with prostaglandin E_2_ (PGE_2_), which binds to its cell-surface receptor, a GPCR, on fibroblasts, activates adenylyl cyclase, and increases intracellular cAMP (Refs. [8,54,114,115,116,117,118,119,120], Figure 1). PGE_2_ can be derived from activated neutrophils or macrophages, or be produced by proliferating fibroblasts and act in an autocrine fashion. Fibroblast proliferation mediated by the production of VDGF, PDGF and FGF by inflammatory cells is also an important contributor to this process. Another key regulator of fibroblast proliferation is transforming growth factor beta 1 (TGF-β1), which is secreted by macrophages and other inflammatory cells and which acts through SMAD pathway signaling to stimulate fibroblast proliferation and production of extra-cellular matrix proteins [108,121,122,123,124]. Tumor necrosis factor alpha (TNF-α), which is produced primarily by neutrophils and activated macrophages [108,120,125,126,127] and which has numerous effects on immune effector cells, also stimulates fibroblast growth and the production of extracellular matrix. It is likely, therefore, that roflumilast, nerandomilast, and other PDE4-selective inhibitors have a dual mechanism of action in these models: (1) they have direct effects on fibroblasts and (2) they down-regulate immune/inflammatory cells that produce factors that stimulate fibroblast growth.
5.7. Actions of Roflumilast and Nerandomilast in Animal Models of Fibrotic Lung Disease
Several well-validated rodent models have provided key insights into the pathogenesis of human fibrotic lung disease and also serve as test platforms for assessing PDE4 inhibitors (and other treatments) in these disorders. The best-described model is bleomycin-induced lung fibrosis in mice. Bleomycin is an anti-neoplastic agent with potent activity in several human cancers; it is a potent generator of free radicals, which accounts for both its antitumor activity and its toxicities. Intra-tracheal treatment of mice with bleomycin produces an intense inflammatory response that is rich in macrophages, neutrophils and lymphocytes and then, over several weeks, progresses to severe fibrosis. TNF-α, along with other cytokines, is felt to be a major contributor to bleomycin-induced lung inflammation and fibrosis. Bleomycin-induced lung inflammation is attenuated by corticosteroids, which also prevent or ameliorate the resulting fibrosis [128]. In contrast, as described above, tobacco smoke typically produces lung inflammation that does not progress to fibrosis and which does not respond to glucocorticoids [8].
All three oral drugs (pirfenidone, nintedanib, and nerandomilast) currently available for the treatment of IPF and PPF have significant activity in bleomycin-induced mouse lung injury models [108,129,130,131,132,133,134,135], and pirfenidone and nintedanib have been used successfully off-label for the treatment of bleomycin lung toxicity in humans [55]; because nerandomilast has only recently been approved, clinical use in human bleomycin toxicity has yet to be reported. Roflumilast also has significant activity in the same mouse model [128,136,137] and has also been used in humans therapeutically for this purpose [55]; head-to-head comparisons between roflumilast and nerandomilast (or other agents) in this setting has yet to be reported.
Apart from bleomycin, exposure to several other agents, including the minerals silica and asbestos, can also produce fibrotic lung disease in mouse models (and also in humans); there is less data on the value of pharmacological interventions in these models [60,61,108].
5.8. PDE4 Expression and Function in Key Pulmonary Cell Types: Relationship to COPD and Pulmonary Fibrosis
Although there are extensive data from pre-clinical models for the efficacy of both roflumilast and nerandomilast in both COPD and pulmonary fibrosis, important questions remain. What specific PDE4 isoforms (e.g., those encoded by PDE4A vs. PDE4B vs. PDE4D, and the various isoforms encoded by each of these genes) are responsible for the effects of both of these drugs in these models? There is surprisingly little reported data on which PDE4 isoforms are expressed and functionally active in each of the pre-clinical models; such knowledge would allow the generation of hypotheses about how they modulate specific PDE4 signaling pathways. Further characterization of these models, especially using the newer technology of single-cell mRNA sequencing, would be very valuable. The current state of knowledge is summarized in Table 3.
In respiratory epithelium and pulmonary ionocytes, PDE4 isoforms are essential modulators of CFTR Cl^−^ channel activity and thereby play an important role in the maintenance of airway surface liquid. PDE4 isoforms are also important modulators of mucus secretion and ciliary function, both of which, along with airway surface liquid, contribute to pulmonary epithelial mucociliary clearance. The release of reactive oxygen species (ROS) and various inflammatory cytokines, such as TNF-α, IL-1 and IL-8, by pulmonary neutrophils is also regulated, at least in part, by various PDE4 isoforms. The production of TNF and other cytokines, and the release of inflammatory mediators and growth factors, by macrophages, neutrophils and fibroblasts in lung tissue also reflects regulation of these functions, at least in part, by PDE4 isoforms. Finally, PDE4 isoforms are present and have important function roles in lymphocytes that have migrated into lung tissue.
Given our current state of knowledge, it is clear that further pre-clinical research is necessary to determine the exact role(s) of individual PDE4 isoforms (e.g., those encoded by PDE4A vs. PDE4B vs. PDE4D, and the various isoforms encoded by each of these genes) in critical aspects of the pathogenesis of COPD, such as small airway inflammation, mucus hypersecretion, the activation and tissue effects of neutrophils and macrophages, and the production of various cytokines. Similarly, much more pre-clinical research is necessary for determining the roles of individual PDE4 isoforms in critical aspects of the pathophysiology of pulmonary fibrosis, such as the migration, proliferation and differentiation of fibroblasts, the activation of the TGF-β pathway in these cells, and the role of fibroblasts in production and remodeling of the extracellular matrix. Data obtained from these studies will provide better pre-clinical rationale for the use of roflumilast in COPD and nerandomilast in pulmonary fibrosis, respectively.
6. Insights from Clinical Investigation, Clinical Trials, and Translational Research
6.1. Lessons from Clinical Trials of Roflumilast in COPD; Role of Inflammation
The clinical data on roflumilast clearly confirms the pre-clinical data that a major target of the drug is PDE4 isoforms in pulmonary inflammatory cells, as opposed to PDE4s in vascular or airway smooth muscle. Early-stage clinical trial data with roflumilast showed that it did not have significant acute bronchodilator activity in COPD ([113], see ref. [8] for a review). Consequently, phase 3 clinical trials of roflumilast have focused on patients with COPD and severe airflow limitation, symptoms of bronchitis, and frequent exacerbations [5,10,11,12], where its beneficial effects on lung inflammation would be more apparent. Other effects of roflumilast, such as its augmentation of the activity of the Cl^−^ ion channel CFTR in lung epithelium, may also contribute to it benefits in these patients (see ref. [9] for a review).
There is an interesting paradox between the therapeutic actions of PDE4-selective inhibitors, such as roflumilast, in obstructive pulmonary disease, compared to the actions of β-adrenergic agonists. β-adrenergic agonists have acute bronchodilator activity [16,140,141], while roflumilast does not. Yet it is clear that both classes of agents act by increasing intracellular cAMP levels (Figure 1). Potential reasons for the differences in therapeutic effect of these two classes of agents may reflect differences in target tissues (i.e., airway smooth muscle vs. polarized pulmonary epithelium and inflammatory cells), or underlying disease (i.e., COPD with an asthmatic component vs. other sub-types of COPD), or delivery (i.e., inhaled vs. oral). Intriguingly, ensifentrine does have acute bronchodilator activity (see references above); whether this therapeutic effect is mediated through PDE4 inhibition, or instead by inhibition of PDE3, or to some other factor (e.g., inhaled vs. oral delivery) remains to be determined.
6.2. Lessons from Human Inflammatory Disorders That Lead to Fibrotic Lung Disease
Patients with several well-defined systemic human immune/inflammatory diseases, including scleroderma and rheumatoid arthritis, can develop interstitial lung disease (ILD), which can progress to IPF or PPF [18,53,142]. Scleroderma (synonym: systemic sclerosis) is characterized by fibrosis of the skin, musculoskeletal system, and other tissues and the development of several different antibodies. Patients with scleroderma can develop ILD that progressed to PPF; nintedanib has been shown to have benefit in pre-clinical models of scleroderma and has been shown in clinical trials to slow the progression to PPF in these patients [54,65,143]. Patients with rheumatoid arthritis, who typically present with symptoms related to the musculoskeletal system, and who develop antibodies against citrullinated proteins, can develop ILD that can progress to PPF [144]; pirfenidone has been shown to have clinical benefit in this specific patient population [145]. Germline variants in a number of genes, most notably MUC5B and SETPC, are major predisposing factors to the development of IPF in many of these disorders [142,146,147]. Patients with these disorders have shown in clinical trials to benefit from nerandomilast [18] and also from pirfenidone and nintedanib (see references above); head-to-head comparisons of these three drugs have yet to be reported. The pre-clinical models of these disorders, the results obtained from many areas of clinical investigation, and the clinical trial data are all consistent with concept that ongoing inflammation is key to fibrotic lung disease and that targeting inflammation is key to the beneficial effects of the three drugs currently available for these disorders.
6.3. PDE4 Inhibitors in COPD and Pulmonary Fibrosis: Different Diseases, Common Mechanisms of Action
Given that inflammation plays an essential role in the pathogenesis of both COPD and pulmonary fibrosis, and that the major therapeutic effects of both roflumilast and nerandomilast are achieved through the targeting of PDE4 isoforms in lung inflammatory cells, what exactly are the differences between these two drugs? In many of the pre-clinical models, the effects of the two drugs appear to be similar. Even their non-inflammatory effects, such as their actions on CFTR (in COPD) and their direct effects on fibroblasts (in pulmonary fibrosis), appear to be similar; indeed, these effects may help patients with both disorders. There are many residual questions: Does the broader action of roflumilast (i.e., activity against all PDE4 isoforms and therefore against an extensive range of pulmonary pathways) contribute to its therapeutic benefit? Alternatively, does the narrower action of nerandomilast (specific for PDE4B and, to some extent, PDE4D isoforms, and the pathways mediated specifically by these isoforms) a major contributor to its therapeutic benefit?
6.4. The Comparative “Druggability” of Roflumilast and Nerandomilast
In the design of small-molecule therapies, many factors influence the “druggability” of any individual compound, which in turn can be pivotal to its eventual success in humans. For example, solubility, protein-binding, stability in vivo, bioavailability, dosing, tissue availability, and pharmacokinetics are key factors. Such factors may have played a central role in the development of nerandomilast and therefore account for its potentially greater benefit over older drugs, such as roflumilast, in pulmonary fibrosis and potentially in other indications [148].
Related, but technically separate, from druggability is the question of tolerability (side effects) in humans. Useful tolerability data can be obtained from early-stage clinical trials, but important data may not emerge until later in the drug-testing process. The extensive phase 3 trials on both roflumilast and nerandomilast provide extensive data on the tolerability of both drugs (see references cited in the first section, above). Both agents are well-tolerated, with discontinuation percentages in the low-single digits. Diarrhea, mild nausea and fatigue are the most common side effects; alterations of mood and affect also occur, but are rarer. All these side effects are typical of PDE4-selective inhibitors as a class. We have developed preliminary data suggesting that the diarrhea seen with PDE4 inhibitors reflects their augmentation of CFTR Cl^−^ channel activity in the GI tract [138], consistent with a class effect. Current data on the relative toxicity/tolerability of roflumilast and nerandomilast needs to be interpreted with caution because of differences in patient populations in the various clinical trials reported to date. Ideally, data comparing the relatively tolerability of these two agents should be obtained by head-to-head, randomized, blinded, comparisons of both tolerability and efficacy. Therefore, a review of comparative head-to-head clinical trials of various PDE4 inhibitors, including these two drugs, in lung and other indications, would certainly be of interest.
7. Ongoing and Possible Future Clinical Trials of PDE4 Inhibitors and Other Agents in COPD and Pulmonary Fibrosis
7.1. Currently Recruiting Clinical Trials of Roflumilast, Nerandomilast, Pirfenidone and Nintedanib
To obtain a better perspective on the future roles of PDE4 inhibitors in COPD and pulmonary fibrosis, we searched www.ClinicalTrials.gov for all actively recruiting clinical trials for roflumilast, nerandomilast, pirfenidone and nintedanib in pulmonary disorders as of 28 August 2025 (see Supplemental File). This search yielded 19 clinical trials; in contrast, a search for ongoing clinical trials of all agents in COPD, performed 30 October 2023, yielded a total of 480 trials [9]. Among the most important trials identified in the current search is a phase 3 multicenter trial comparing roflumilast to the antimicrobial azithromycin as prevention of COPD exacerbations (NCT04069312) and a comparison of roflumilast to the inhaled PDE4 inhibitor tanimilast as maintenance therapy in COPD (NCT04636814; for further discussion of the pre-clinical and early-stage development of tanimilast and ensifentrine, see a prior review [9]). Also noteworthy is the PROGRESSION trial (NCT03939520), a randomized phase 3 trial comparing the efficacy and tolerance of the combination of pirfenidone and nintedanib, compared to either as monotherapy, in previously treated patients with IPF.
7.2. Considerations for the Design of Future Clinical Trials
Review of completed and ongoing clinical trials of PDE4-selective inhibitors in COPD and pulmonary fibrosis has shown that much progress has been made, but also that there is room for improvement in trial design. One of the key deficiencies is the lack of head-to-head comparisons of many of the available, or soon-to-be-available agents, as discussed in detail in several sections above. Several other potential areas of improvement can be identified: At what point should intervention be initiated (e.g., pre-clinical vs. symptomatic)? What biomarkers (e.g., information obtained from blood and/or tissue samples) or other procedures, such as imaging or spirometry [149,150]), can be used to determine eligibility and to stratify patients on these trials, and in clinical practice? Several recent attempts to identify and validate biomarkers in both COPD and pulmonary fibrosis are discussed in the next section. What end-points should be chosen in the design of the newer clinical trials? Greater use of biomarkers and other intermediate end-points should facilitate more rapid, and potentially smaller, clinical trials and is an important goal. Conversely, trials that focus on clinically meaningful endpoints (reductions in hospitalizations, exacerbations, and improvements in quality of life) would seem to be essential for assessing the ultimate value of newer therapies [151,152]. A related question is that of duration: given the chronicity of these diseases, how long should patients be treated on a trial before determining whether the intervention(s) are effective? In the ongoing interactions between drug-development teams, regulators, clinical trialists and patients, careful attention to these issues would seem to be essential.
8. Future Directions: Subclassifications, Biomarkers, and Pathways
Several new features are dominating the evolving landscape of novel therapies in both COPD and pulmonary fibrosis. The first, clearly, is the novel and important division of both these disorders into broad sub-classes that guide therapy and that are also useful in stratifying patients on clinical trials. In COPD, the use of eosinophilia as a marker for Type 2 inflammation has identified a subclass of patients that respond to cytokine-modulating agents, such as dupilumab and mepolizumab. In pulmonary fibrosis, the division of patients into two broad categories—IPF and PPF—has been useful in stratifying patients into appropriate clinical trials and, in the case of PPF, has the potential to base therapies on the pathobiology of the underlying disease. Current national and international guidelines are beginning to incorporate these concepts into routine clinical practice [16,21,29,31]. These stratification criteria lead to a tentative COPD classification system that reflects the underlying pathophysiology and which directly affects clinical decision-making (Figure 3, refs. [9,153,154]).
The second feature, related to the first, is the development of biomarkers that (1) reflect underlying disease mechanisms; (2) can be used in decision-making in individual patients; (3) can be used to stratify patients on clinical trials; (4) can serve as end-points in early-stage clinical trials. An emerging biomarker in COPD is an elevated fractional exhaled nitric oxide (FeNO) or alveolar NO level, which reflects ongoing inflammation, is frequently correlated with eosinophilia, and helps to identify patients with a COPD and asthma overlap phenotype [155,156,157,158,159,160,161]. Several recent publications have shown grounds for promise in identifying additional blood and tissue biomarkers, such as IL-1β, CXCL10, Eotaxin-3, PARC and IgE, in both COPD [25,39,41,162] and pulmonary fibrosis [53,61,163]. New, emerging technologies, such as single-cell sequencing on patient-derived samples (e.g., blood, sputum, BAL fluid and biopsies; ref. [164]), should contribute substantially to biomarker efforts. Additionally, there is emerging data on the use of germline genetic markers predictive of response; among the most interesting of these is the identification of polymorphisms (SNPs) in the PDE4 genes that correlate with responsiveness to the PDE4-selective inhibitor apremilast in psoriasis [165]; there is a need for similar studies in the pulmonary area.
It is apparent that study of the mechanisms of action of PDE4 drugs in a variety of human diseases, including lung disease, has enabled progress on understanding fundamental disease mechanisms in COPD and pulmonary fibrosis. Further study of these mechanisms is essential for the development of newer agents in these diseases, which would include not only PDE4 inhibitors, but agents targeting novel targets. One example is the possible use of CFTR potentiators in the treatment of COPD, which has a compelling pre-clinical rationale (reviewed in ref. [9]) and has been the subject of early-phase clinical trials [166].
Further effort, both pre-clinical and clinical, will be required to determine whether the differences postulated between various drugs in different pulmonary diseases reflects differences in the action(s) of the drugs (i.e., selectivity for certain targets), or difference in the underlying diseases (i.e., the presence of specific drug targets in different disorders), or other factors. We are in the midst of an exciting time for pulmonary therapeutics and additional insights and additional agents are emerging in a rapid fashion.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kelly M.P. Nikolaev V.O. Gobejishvili L. Lugnier C. Hesslinger C. Nickolaus P. Kass D.A. Pereira de Vasconcelos W. Fischmeister R. Brocke S. Cyclic nucleotide phosphodiesterases as drug targets Pharmacol. Rev.20257710004210.1016/j.pharmr.2025.10004240081105 · doi ↗ · pubmed ↗
- 2Hanania N.A. Celli B.R. Phosphodiesterase Inhibition as a Therapeutic Strategy for Chronic Obstructive Pulmonary Disease: Where We Have Been and What Lies Ahead Chronic Obstr. Pulm. Dis.202512829210.15326/jcopdf.2024.055939688360 PMC 11925072 · doi ↗ · pubmed ↗
- 3Conti M. Beavo J. Biochemistry and physiology of cyclic nucleotide phosphodiesterases: Essential components in cyclic nucleotide signaling Annu. Rev. Biochem.20077648151110.1146/annurev.biochem.76.060305.15044417376027 · doi ↗ · pubmed ↗
- 4Baillie G.S. Tejeda G.S. Kelly M.P. Therapeutic targeting of 3′,5′-cyclic nucleotide phosphodiesterases: Inhibition and beyond Nat. Rev. Drug Discov.20191877079610.1038/s 41573-019-0033-431388135 PMC 6773486 · doi ↗ · pubmed ↗
- 5Janjua S. Fortescue R. Poole P. Phosphodiesterase-4 inhibitors for chronic obstructive pulmonary disease Cochrane Database Syst. Rev.20205 CD 00230910.1002/14651858.cd 002309.pub 632356609 PMC 7193764 · doi ↗ · pubmed ↗
- 6Francis S.H. Blount M.A. Corbin J.D. Mammalian cyclic nucleotide phosphodiesterases: Molecular mechanisms and physiological functions Physiol. Rev.20119165169010.1152/physrev.00030.201021527734 · doi ↗ · pubmed ↗
- 7Maurice D.H. Ke H. Ahmad F. Wang Y. Chung J. Manganiello V.C. Advances in targeting cyclic nucleotide phosphodiesterases Nat. Rev. Drug Discov.20141329031410.1038/nrd 422824687066 PMC 4155750 · doi ↗ · pubmed ↗
- 8Hatzelmann A. Morcillo E.J. Lungarella G. Adnot S. Sanjar S. Beume R. Schudt C. Tenor H. The preclinical pharmacology of roflumilast—A selective, oral phosphodiesterase 4 inhibitor in development for chronic obstructive pulmonary disease Pulm. Pharmacol. Ther.20102323525610.1016/j.pupt.2010.03.01120381629 · doi ↗ · pubmed ↗