AoMbp1 Governs Conidiation and Trap Morphogenesis in Arthrobotrys oligospora Via Direct Transcriptional Activation of the MAPK Sensor AoSho1
Ruobing Li, Lixiang Wei, Yanseng Sun, Chengzhi Zhang, Yuhang Nie, Qinglong Meng, Shuang Chen, Ming Wu, Xuepeng Cai, Jie Li, Qingling Meng, Jun Qiao

TL;DR
This study identifies AoMbp1 as a key regulator in a fungus that traps nematodes, controlling its growth, stress tolerance, and trap formation.
Contribution
The discovery of AoMbp1's direct activation of AoSho1 establishes a new regulatory mechanism in nematode-trapping fungi.
Findings
AoMbp1 deletion impairs growth, conidiation, trap formation, and stress tolerance in A. oligospora.
AoMbp1 directly activates AoSho1, a key MAPK sensor, linking transcriptional regulation to trap morphogenesis.
Transcriptome analysis shows widespread gene downregulation in the MAPK pathway upon AoMbp1 deletion.
Abstract
The nematode-trapping fungus (NTF) Arthrobotrys oligospora (A. oligospora) is a promising biocontrol agent, but the transcriptional regulators governing its predation remain poorly understood. Here, we demonstrated that the APSES transcription factor AoMbp1 is a master regulator of its development and stress adaptation. Deletion of AoMbp1 severely impaired mycelial growth, conidiation, trap formation, and tolerance to oxidative and osmotic stresses. Transcriptome analysis revealed that these defects were associated with the widespread downregulation of genes, including those within the MAPK signaling pathway. Crucially, we showed that AoMbp1 directly binds to the promoter of AoSho1, a key upstream sensor of the MAPK cascade, and activates its expression. This finding establishes a direct AoMbp1-AoSho1 regulatory axis controlling trap morphogenesis and environmental adaptation. Our study…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
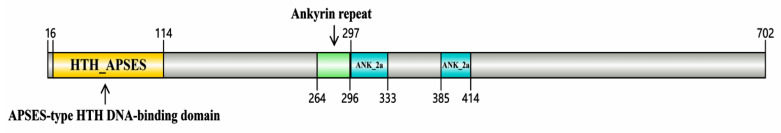 Figure 1
Figure 1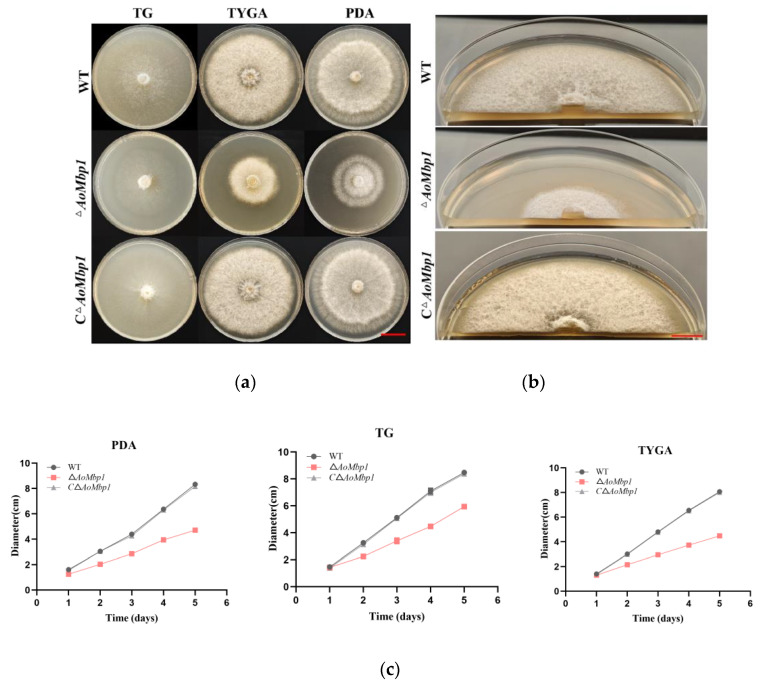 Figure 2
Figure 2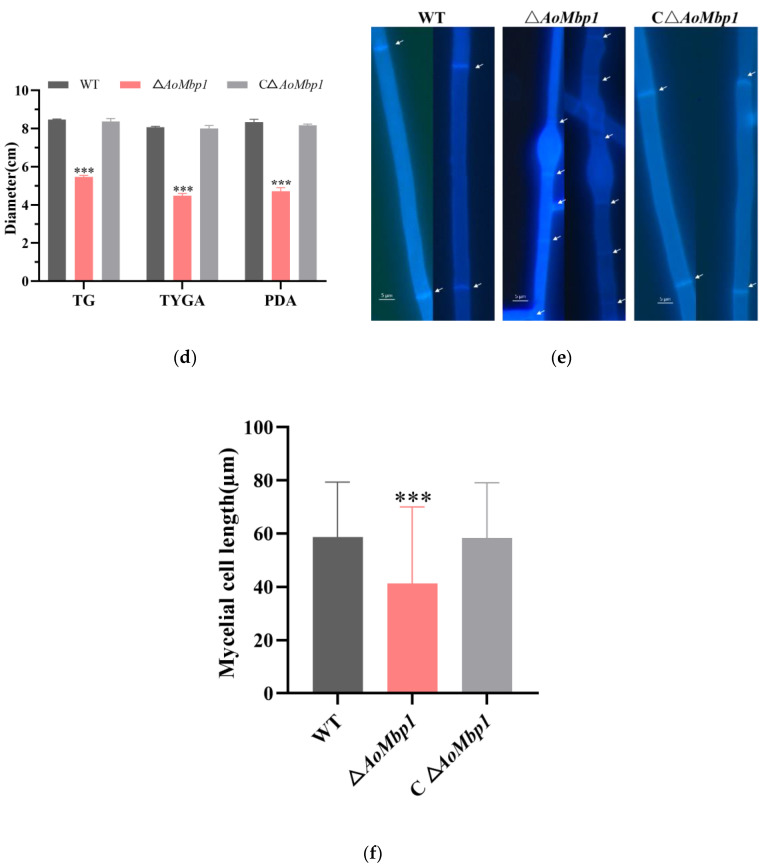 Figure 3
Figure 3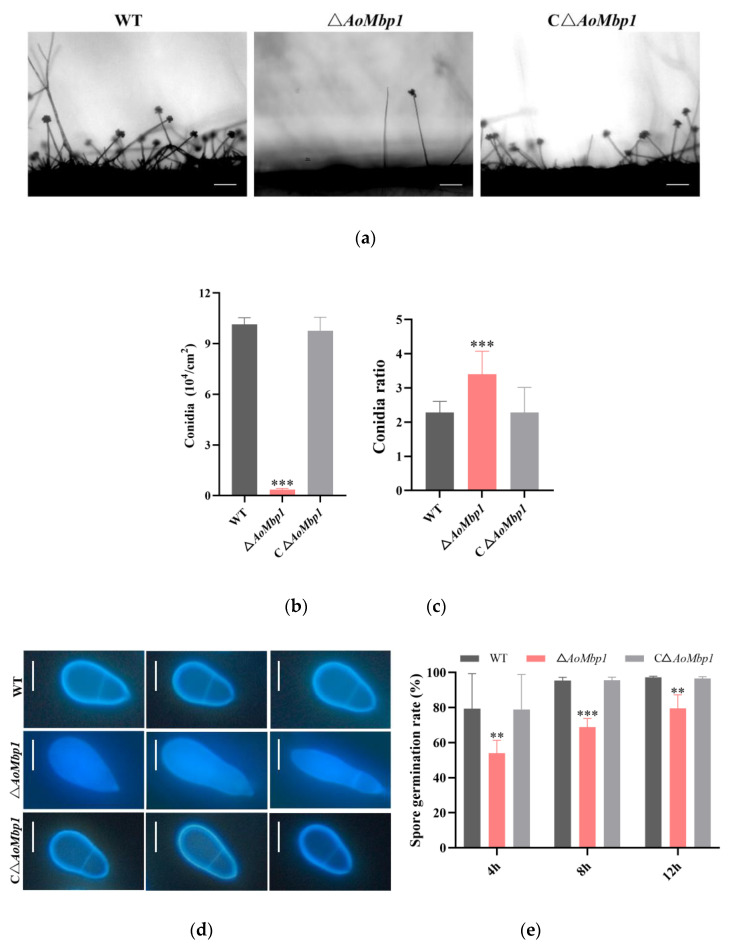 Figure 4
Figure 4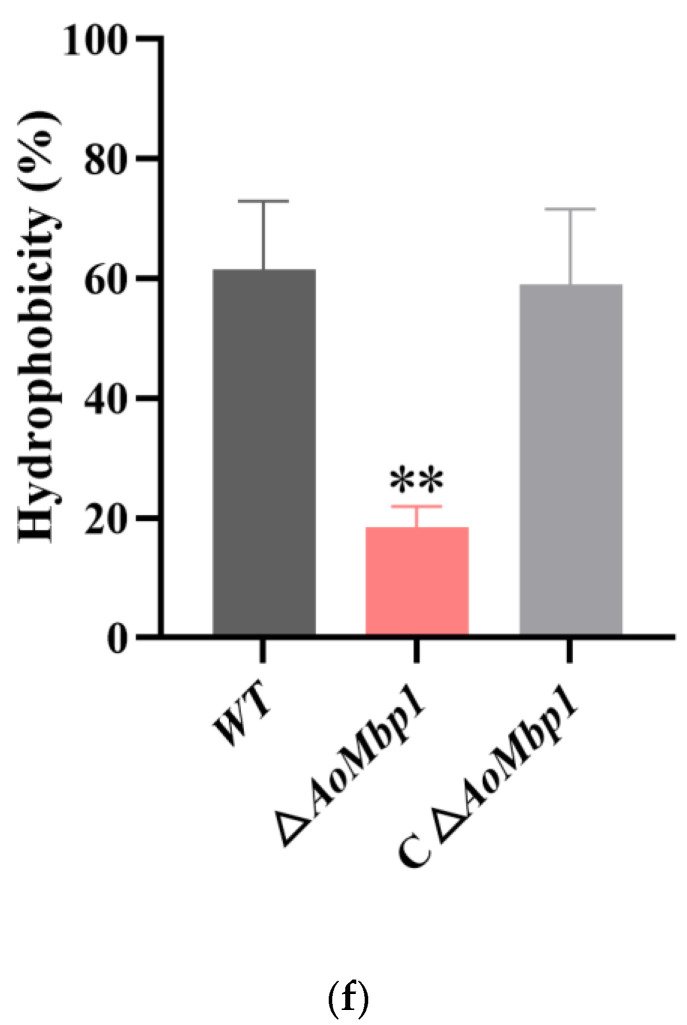 Figure 5
Figure 5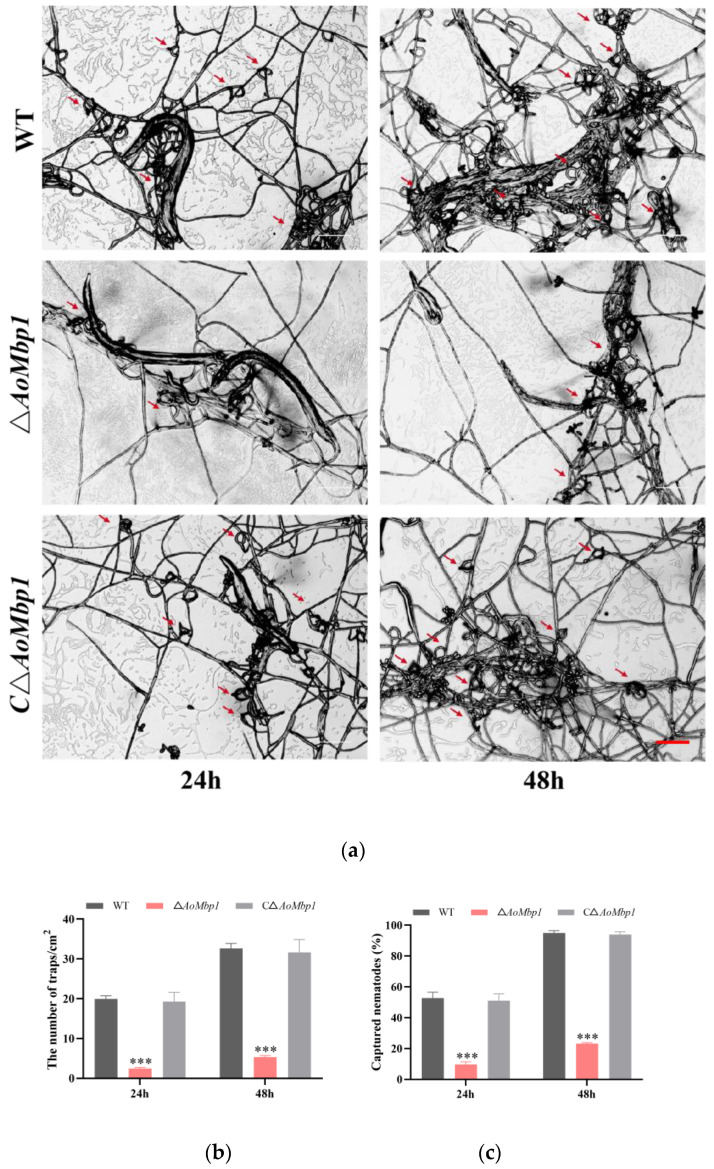 Figure 6
Figure 6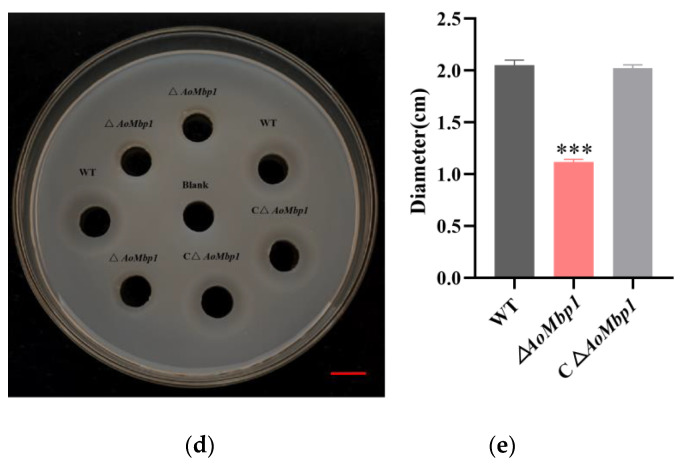 Figure 7
Figure 7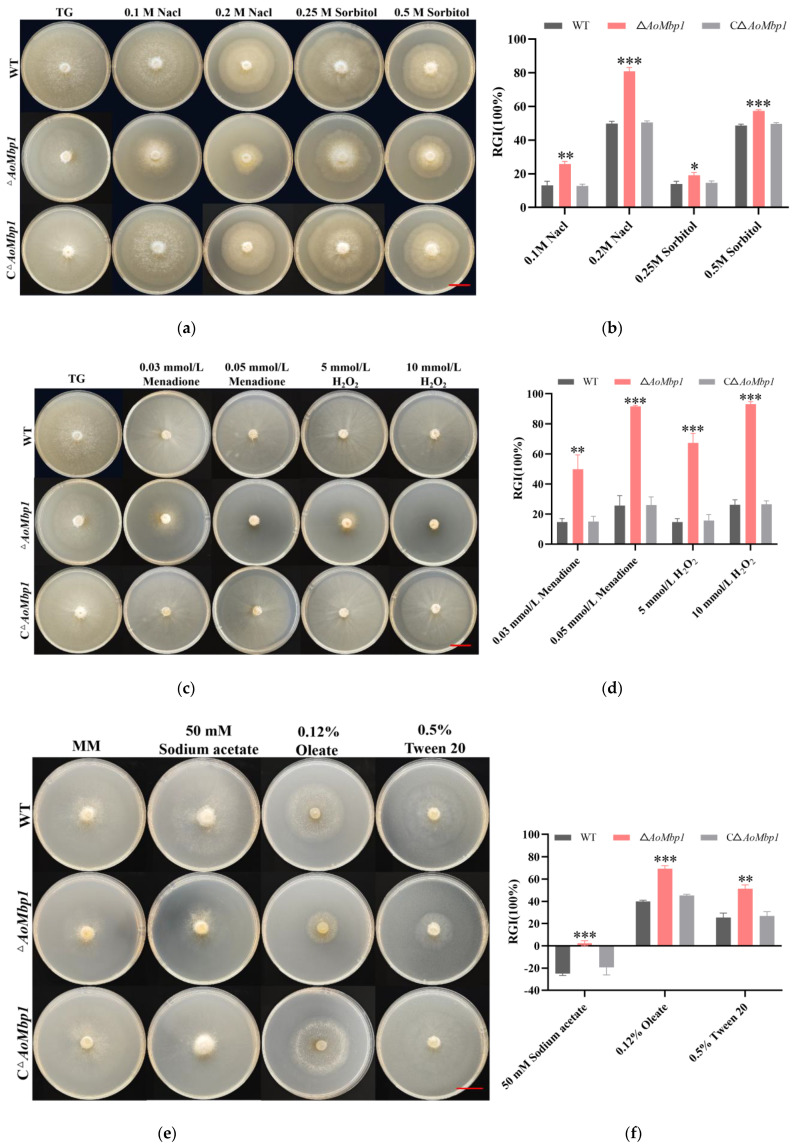 Figure 8
Figure 8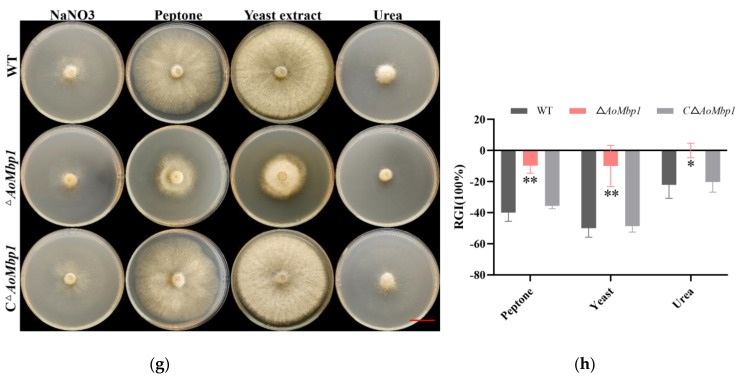 Figure 9
Figure 9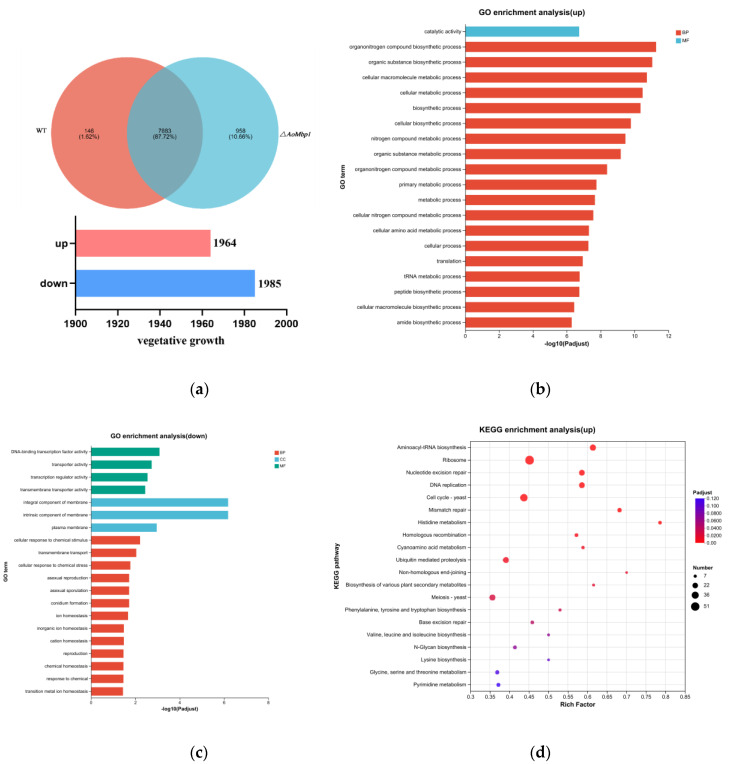 Figure 10
Figure 10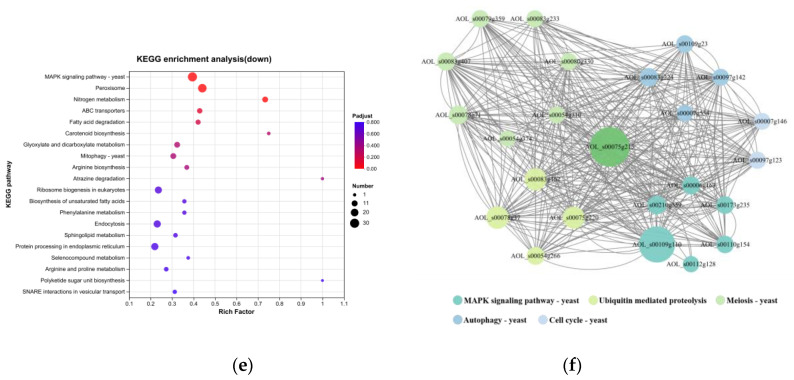 Figure 11
Figure 11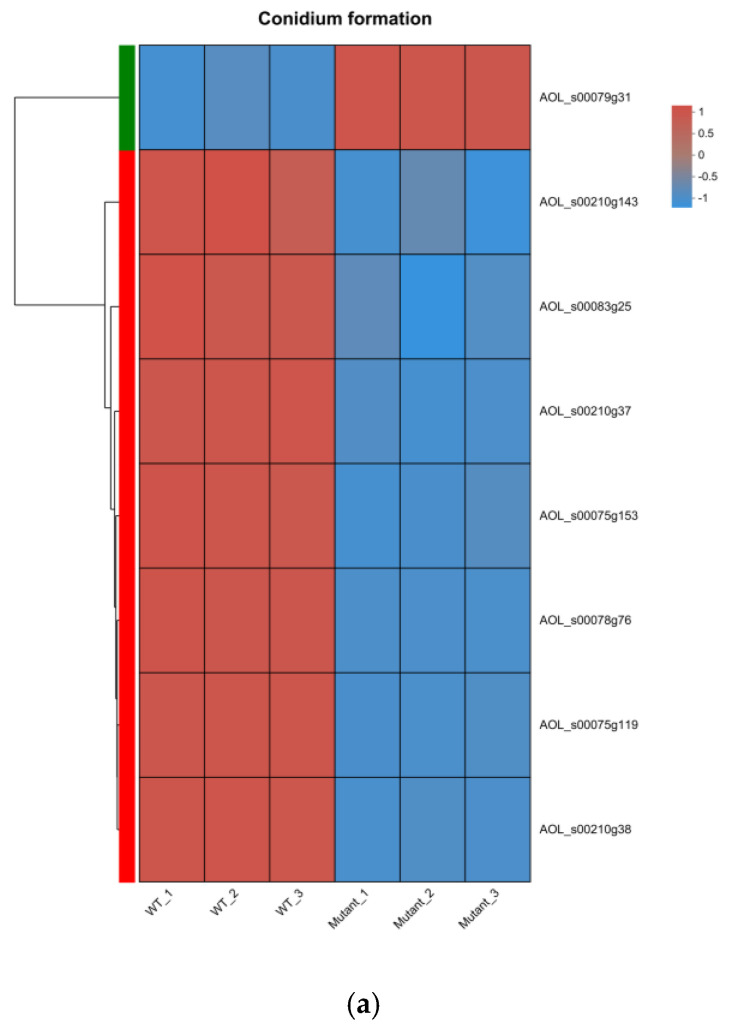 Figure 12
Figure 12 Figure 13
Figure 13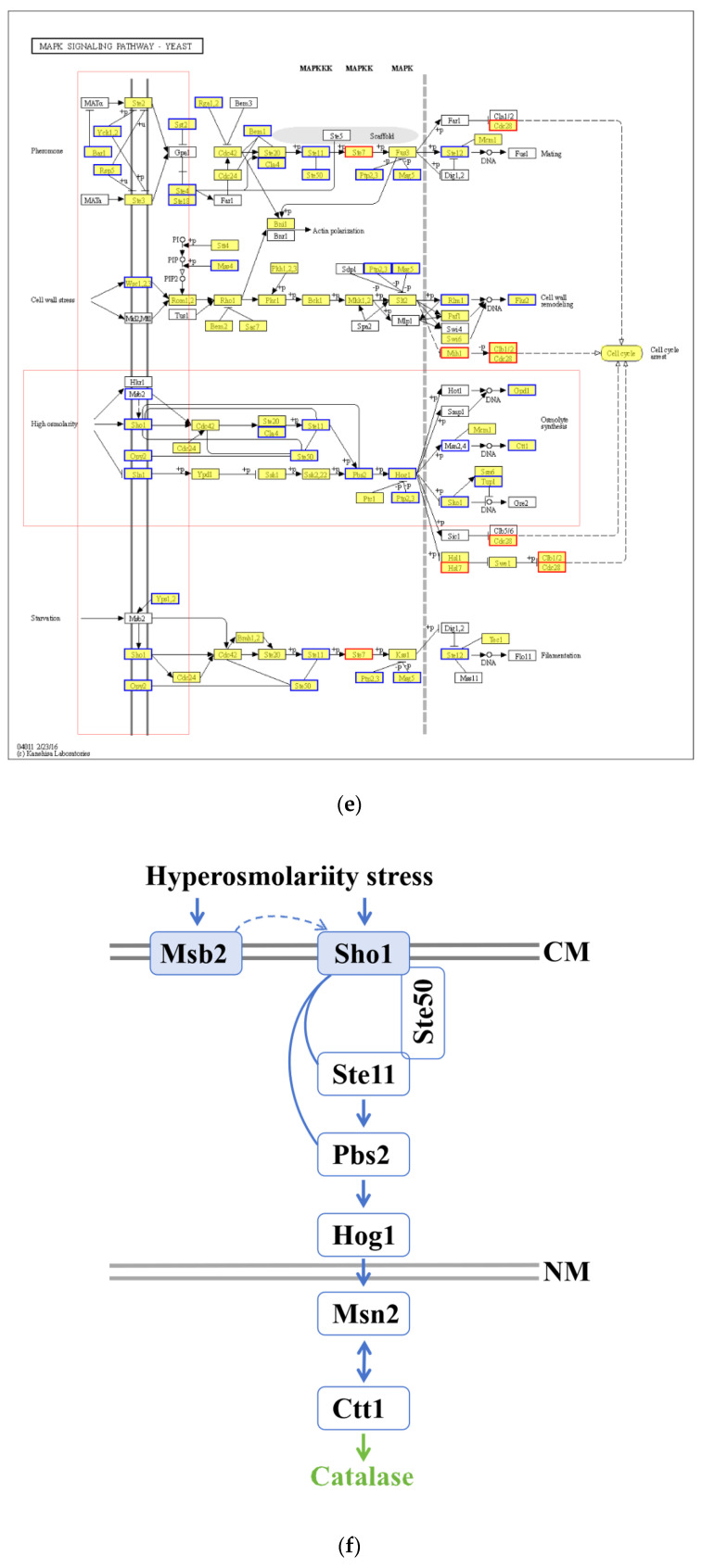 Figure 14
Figure 14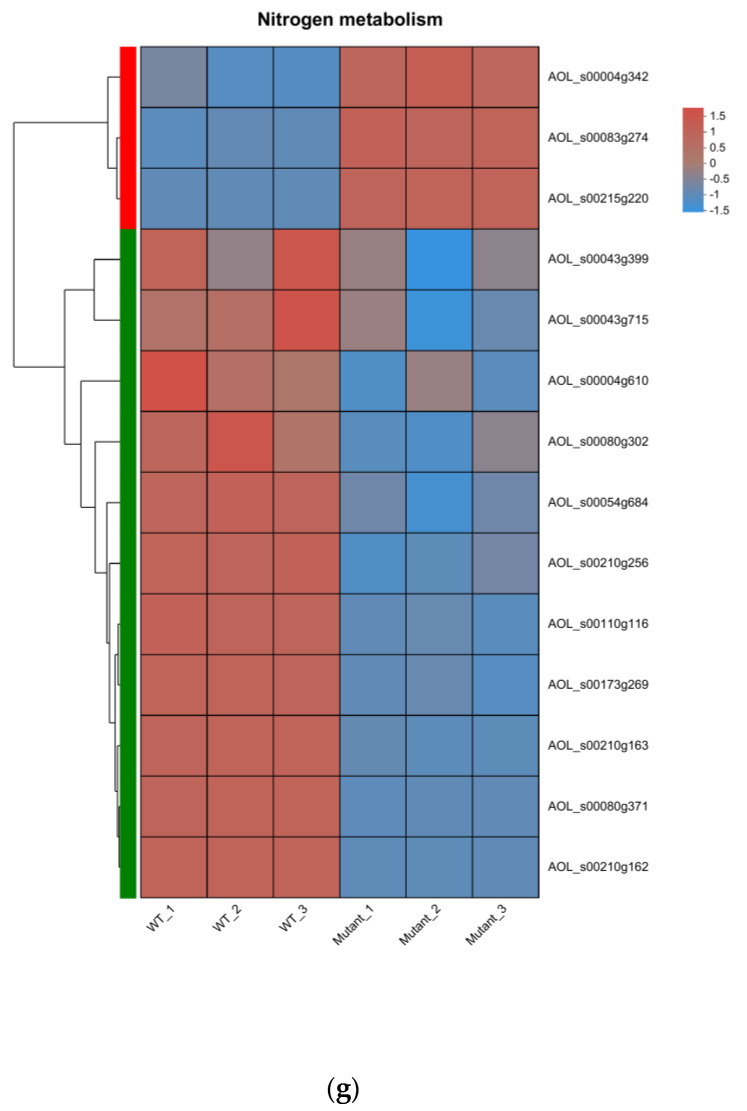 Figure 15
Figure 15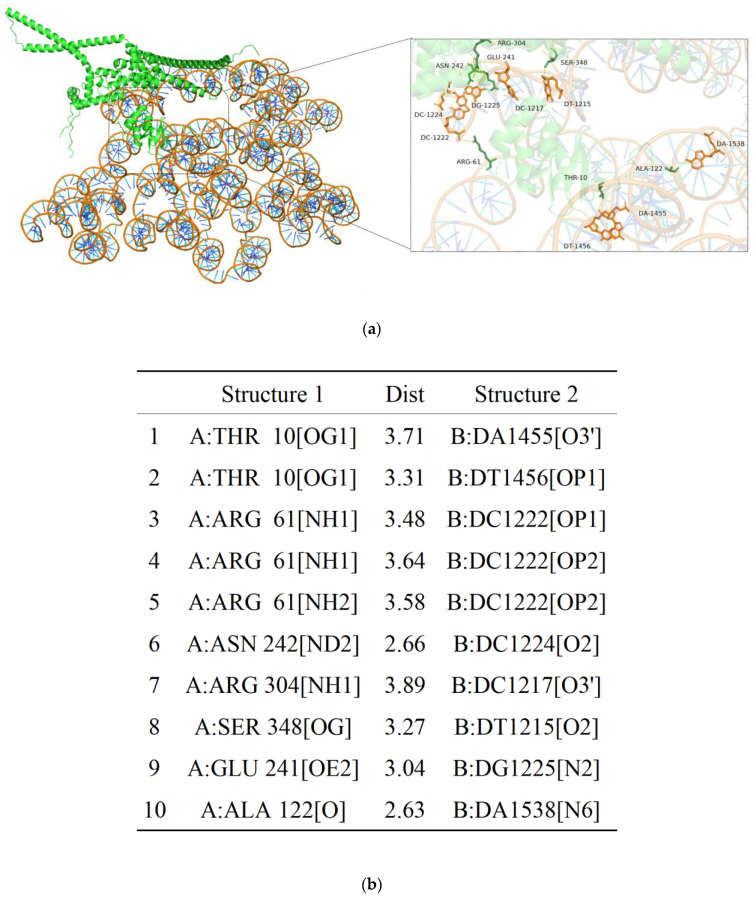 Figure 16
Figure 16 Figure 17
Figure 17 Figure 18
Figure 18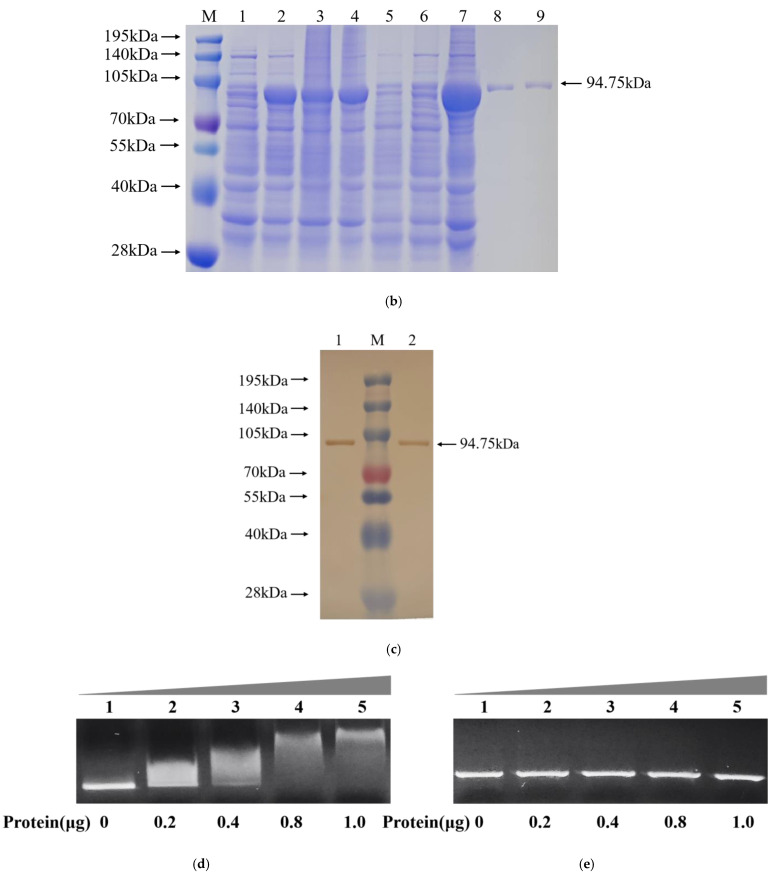 Figure 19
Figure 19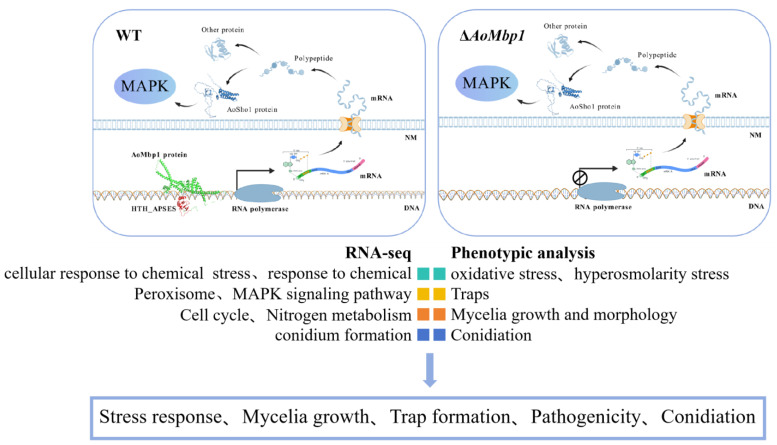 Figure 20
Figure 20- —National Natural Science Foundation of China
- —National Key Research and Development Program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal and yeast genetics research · Plant-Microbe Interactions and Immunity · Entomopathogenic Microorganisms in Pest Control
1. Introduction
Gastrointestinal nematodes (GINs) are widespread parasites in herbivorous species, posing a substantial challenge to the sustainable development of the global herbivore industry [1]. Infections caused by nematodes from genera such as Ostertagia, Trichostrongylus, Haemonchus and Nematodirus have been shown to result in increased feed costs, growth retardation, weakened immunity, and even mortality in young herbivores [2,3]. These consequences lead to significant economic losses in the herbivore industry [4].
Currently, the application of chemical dewormers during the breeding process is a common strategy for preventing and managing gastrointestinal nematode infections in livestock. However, the widespread use of these chemical treatments has led to severe challenges, including the development of drug-resistant worm strains, the accumulation of drug residues in animal products, and environmental contamination [5]. Therefore, there is an urgent need to develop innovative methods for the prevention and control of gastrointestinal nematode diseases in herbivores to ensure the sustainable and healthy growth of the animal husbandry sector.
Arthrobotrys oligospora (A. oligospora), a filamentous fungus, has been shown to produce specialized structures such as mycorrhizal rings and adhesive mycorrhizal webs [6,7], which function as snares to capture nematode larvae [8,9]. Owing to its predatory behavior, this fungus has demonstrated significant potential in the biocontrol of nematode diseases. Nevertheless, the mechanisms governing the spore-producing capabilities and predatory activity of A. oligospora, one of the most promising microorganisms for biocontrol applications, remain incompletely understood. Among the transcription factors identified in filamentous fungi, the APSES family constitutes a distinct class of fungal regulatory proteins. These transcription factors feature helix-loop-helix DNA-binding domains that play key roles in essential biological processes, including fungal growth, spore production, cell differentiation, secondary metabolite biosynthesis, and pathogenicity [10,11]. In Saccharomyces cerevisiae, the APSES family member Mbp1 regulates the expression of genes involved in the G1/S phase transition by binding to the Mlu1 cell cycle box (MCB box) in promoter regions. Deletion of Mbp1 has been associated with impaired pseudohyphal differentiation and disruptions in respiratory metabolism [12,13]. Similarly, Mbp1 in Histoplasma capsulatum influences morphological transitions between yeast and mycelial forms while also modulating pathogenicity and oxidative stress tolerance [14]. In Pleurotus ostreatus, Mbp1 plays a pivotal role in regulating β-glucan and chitin synthesis [15,16]. In Beauveria bassiana, Mbp1 is essential for mycelial differentiation and virulence expression [17,18]. Similarly, in Fusarium verticillioides, Mbp1 is critical for nutrient uptake, conidial development, stress response, and pathogenicity [19]. Despite substantial research into the role of Mbp1 in other fungal systems, its biological function in A. oligospora remains elusive. In a previous study conducted by our group, we discovered that the expression levels of AoMbp1 significantly increase during both the spore-producing and trap-forming stages of A. oligospora (Figure S1). This observation suggests that AoMbp1 is intricately linked to the development of spore-producing and trap-forming structures in this fungus.
This study aimed to bridge this knowledge gap by systematically investigating the function of AoMbp1 in the nematode-trapping fungus A. oligospora, with a focus on its potential role in connecting developmental processes with stress adaptation pathways. We demonstrated that AoMbp1 coordinates growth, conidiation, trap formation, and stress responses by directly regulating AoSho1, a key upstream component of the MAPK signaling pathway. This work provides new insights into the transcriptional mechanisms governing fungal predation.
2. Materials and Methods
2.1. Plasmids, Strains, Nematodes and Culture Conditions
The pCSN44 plasmid, which carries the hygromycin resistance gene (hph), and the pPk2-Bar plasmid, which contains the glufosinate resistance gene (bar), were obtained from the Key Laboratory of Preventive Veterinary Medicine at Shihezi University. The recombinant plasmids were constructed using the E. coli strain DH5α, which was cultured in LB media at 37 °C. The A. oligospora XJ-2 wild-type (WT) strain was also obtained from the same laboratory and cultured on YPSSA media at 28 °C. Additionally, Caenorhabditis elegans was cultured on NGM at 26 °C for 6–7 days prior to the bioassays.
2.2. Analysis of the Molecular Characteristics of the AoMbp1 Protein
The homologous protein AOL_s00075g215 (AoMbp1) was identified in the A. oligospora genome through BLAST analysis, and the protein sequence of Saccharomyces cerevisiae Mbp1 (CAA98618) was used as a query. Primers P1/P2 were designed on the basis of the AoMbp1 gene sequence, and the gene was subsequently amplified and sequenced via PCR (Table S1). The molecular characterization of the protein encoded by the AoMbp1 gene was conducted via DNAMAN software (https://www.lynnon.com/dnaman.html, accessed on 5 May 2025), the ExPASy online tool (https://web.expasy.org/translate/, accessed on 5 May 2025), and ScanProsite (https://prosite.expasy.org/scanprosite/, accessed on 5 May 2025). Additionally, MEGA 11 software was used to construct a phylogenetic tree of the proteins belonging to the APSES family. The nucleotide sequence of the AoMbp1 gene has been deposited in the NCBI GenBank database under accession number PV872033 (https://www.ncbi.nlm.nih.gov/nuccore, accessed on 5 May 2025).
2.3. Construction of AoMbp1 Gene Deletion and Complementation Strains
The deletion strain ΔAoMbp1 and the complementation strain CΔAoMbp1 were constructed through homologous recombination. Briefly, the primers P3/P4, P5/P6, and P7/P8 were specifically designed to amplify the homology arm sequences flanking the 5′ and 3′ regions of the AoMbp1 gene, along with the PtrpC-Hph expression cassette (Table S1). Gene splicing by overlap extension PCR (SOE-PCR) was employed to construct the knockout vectors, which were subsequently transformed into protoplasts of the wild-type (WT) strain. To generate the complementation strain CΔAoMbp1, the plasmid pUC19-CΔAoMbp1 was assembled via seamless cloning. Using this plasmid as a template, the primers PM1/PM2 were designed to amplify the full-length complement fragment (Table S1). The complement fragment was then introduced into the protoplasts of the confirmed ΔAoMbp1 deletion strain via protoplast transformation. PCR amplification screening was performed via primers P1/P2 to identify the ΔAoMbp1 gene deletion mutant and the CΔAoMbp1 complementation strain.
2.4. Phenotypic Analysis of Different Strains
The WT, ΔAoMbp1, and CΔAoMbp1 strains were cultured on TYGA, PDA, and TG solid media, respectively, at 28 °C for 5 days, with three biological replicates per strain. Colony morphology and aerial mycelial growth were assessed, and colony diameters were measured daily. To evaluate conidiation, germination rates, and surface hydrophobicity, the strains were incubated on CMY medium at 28 °C for 8 days in three independent experiments. Fresh conidia and hyphae from each strain were stained with calcofluor white (CFW) to observe septa in both hyphae and conidia. For trap formation assays, each strain was grown on CMA medium. Three holes were punched along the periphery of the colony, and after mycelia grew into the holes, approximately 200 nematodes were added to each well to induce trap formation. The numbers of traps and captured nematodes were recorded for each strain at 24 h and 48 h post-induction, with three biological replicates. Furthermore, each strain was incubated in PL-4 liquid medium at 28 °C for 6 days. The culture broth was then filtered, and extracellular protease activity was assessed qualitatively by measuring the size of transparent zones on skim milk powder plates in three independent experiments.
2.5. Analysis of Stress Tolerance
Mycelial plugs of each strain were inoculated onto TG agar plates supplemented with specific chemical stressors and incubated at 28 °C for 5 days. For hyperosmotic stress, the medium was supplemented with sodium chloride (0.1 or 0.2 mol/L) or sorbitol (0.25 or 0.5 mol/L). For oxidative stress, hydrogen peroxide (5 or 10 mmol/L) or menadione (0.03 or 0.05 mmol/L) was added to the medium. After incubation, colony diameters were measured, and the relative growth inhibition (RGI) was calculated as previously described. The ability of each strain to utilize different carbon and nitrogen sources was evaluated on minimal medium (MM) supplemented with various fatty acids (50 mM sodium acetate, 0.12% oleic acid, or 0.5% Tween 20) or nitrogen sources (urea, peptone, or yeast extract). Colony diameters were recorded after 5 days of incubation at 28 °C, and the RGI was calculated. All experiments were performed with at least three biological replicates [20,21].
2.6. Transcriptome Sequencing and Analysis of Differentially Expressed Genes
Briefly, the WT and ΔAoMbp1 strains were cultured on YPSSA media at 28 °C for 6 days. Mycelial samples, each with three biological replicates, were collected and sent to Shanghai Meiji Biomedical Science and Technology Co., Ltd. (Shanghai, China), for transcriptome sequencing. The raw RNA-seq data have been deposited in the NCBI Sequence Read Archive under BioProject accession PRJNA1292889. These resulting transcriptome data were subsequently analyzed via a cloud platform. The differentially expressed genes (DEGs) were subjected to Gene Ontology (GO) functional enrichment analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis. Moreover, five DEGs were randomly selected, and their transcriptomic data were validated through reverse transcription–quantitative polymerase chain reaction (RT-qPCR). Additionally, potential proteins that interact with AoMbp1 were examined via the STRING protein-protein interaction network platform. Protein-protein interaction (PPI) networks were visualized and analyzed with Cytoscape (v.3.10.0). In addition, JASPAR software (https://jaspar.elixir.no/, 5 May 2025) was used to screen and identify potential target genes regulated by AoMbp1.
2.7. Molecular Docking Analysis of AoMbp1 with the Promoter of a Downstream Target Gene
To screen candidate downstream target genes identified from transcriptome data, molecular docking was performed between AoMbp1 and the promoter DNA. The three-dimensional structures of the AoMbp1 protein and the promoter DNA fragment were modeled using AlphaFold 3 [22]. Protein-DNA docking was carried out with the HDOCK server. The complex with the highest confidence score was selected for further analysis of its binding interface using PDBePISA (https://www.ebi.ac.uk/pdbe/pisa/ accessed on 5 May 2025) and visualized with PyMOL, v.3. As molecular docking provides predictive insights, the top-ranked candidate interaction was subsequently validated experimentally using yeast one-hybrid (Y1H) and electrophoretic mobility shift assays (EMSA).
2.8. Yeast One-Hybrid (Y1H) Assay
The promoter fragment of the target gene (1043 bp) was amplified via the primers proAoSho1-F/proAoSho1-R (Table S1). It was seamlessly ligated into the pAbAi vector, generating the recombinant bait vector proAoSho1-AbAi. This recombinant vector was subsequently transformed into Y1HGold yeast strains, and the transformants were cultivated on SD/-Ura media to develop the yeast bait strain. The appropriate AbA screening concentration was then determined. Simultaneously, the full-length cDNA sequence of AoMbp1 was amplified via the primers AoMbp1-F/AoMbp1-R (Table S1). It was ligated into the pGADT7 vector to construct the pGADT7-AoMbp1 vector. The pGADT7-AoMbp1 plasmid, along with the pGADT7 empty plasmid (serving as the negative control), was transformed into yeast bait strain receptor cells. The transformation products were plated on SD/-Leu/-Ura media and incubated at a constant temperature of 30 °C for 3–5 days. After that, the positive clones identified on the primary screening plates were cultured overnight in liquid SD/-Leu media, and the resulting cultures were diluted to OD_600nm_ values of 0.2, 0.02, and 0.002. These dilutions were subsequently spotted onto SD/-Leu/AbA screening media containing the same AbA concentration. The plates were subsequently sealed and incubated at 30 °C for 3–5 days, after which growth was observed.
2.9. Electrophoretic Mobility Shift Assay (EMSA)
Briefly, the recombinant protein AoMbp1 was successfully produced via a prokaryotic expression system. The coding sequence of AoMbp1 was amplified from cDNA via F1/R1 primers, and the recombinant vector pMD19-AoMbp1 was constructed (Table S1). Both the pMD19-AoMbp1 and pET-32a plasmids were enzymatically digested with KpnI and HindIII, respectively. Following cleavage, the fragments of the pET-32a vector and AoMbp1 were recovered. Under the action of T4 DNA ligase, the target fragment was ligated, resulting in the formation of the recombinant plasmid pET-AoMbp1, which was subsequently transformed into BL21 (DE3). The expression of the recombinant product was induced with IPTG for six hours, after which the AoMbp1 protein was purified via a His-Ni affinity purification column. The efficiency of protein expression and purification was analyzed through SDS-PAGE and Western blot techniques. Additionally, the promoter DNA fragment of the AoSho1 gene (1043 bp) was amplified via the primer pair F2/R2 (Table S1). The AoSho1 promoter fragment was then incubated with varying concentrations of purified AoMbp1 protein in gel shift buffer for 30 min at room temperature. Bovine serum albumin (BSA) was included as a nonspecific control protein. The resulting mixtures were subjected to 1% agarose gel electrophoresis for analysis, and the results were documented through imaging [23].
2.10. Statistical Analysis
Statistical analyses were performed using GraphPad Prism 5.0 software. Data are presented as means ± standard deviation (SD). Unpaired two-tailed Student’s t-tests were used for comparisons between two groups. For comparisons among multiple groups, one-way analysis of variance (ANOVA) was applied, followed by Tukey’s Post Hoc test. Differences were considered statistically significant at * p < 0.05, ** p < 0.01, and *** p < 0.001.
3. Results
3.1. Molecular Characterization of the Transcription Factor AoMbp1 of A. oligospora
Phylogenetic analysis of the APSES family of transcription factors revealed the presence of five APSES family members in the A. oligospora genome: AoMbp1 (AOL_s00075g215), AoSwi6 (AOL_s00109g110), AoStuA (AOL_s00083g25), AoXbp1 (AOL_s00006g32), and AoBfp4 (AOL_s00109g183) (Figure S2). On the basis of the complete genome sequence of A. oligospora in NCBI, a 357 bp sequence between AOL_s00075g214 and AOL_s00075g215, which can drive the expression of these two genes and is a bidirectional promoter, was identified (Figure S3). The AoMbp1 gene (GenBank accession: PV872033) from the A. oligospora XJ-2 wild-type (WT) strain comprises 2330 bp, containing three introns and an open reading frame (ORF) of 2109 bp. The ORF encodes a protein comprising 702 amino acids. Moreover, the AoMbp1 protein includes a conserved APSES-type helix-turn-helix (HTH) DNA-binding domain along with two ankyrin repeat domains (Figure 1). Comparison of amino acid sequences revealed homology rates of 51.28%, 49.66%, and 50.59% between AoMbp1 and the APSES domain proteins of Tuber magnatum, Ascodesmis nigricans, and Wilcoxina mikolae, respectively (Figure S4). Furthermore, the phylogenetic tree revealed that AoMbp1 is highly conserved with the APSES domain proteins of Saitoella complicata, Dacryopinax primogenitus, Fomitiporia mediterranea MF3/22, and Guyanagaster necrorhizus MCA 3950 (Figure S5).
3.2. AoMbp1 Is Involved in the Regulation of Mycelial Growth
The ΔAoMbp1 deletion strain and CΔAoMbp1 complementation strain were constructed via homologous recombination (Figure S6a–d). Compared with the WT strain, the CΔAoMbp1 strain did not significantly differ. In contrast, the mycelia of the ΔAoMbp1 deletion strain presented a distinct trophic growth defect (Figure 2a), characterized by a flattened, compact colony morphology and a notable reduction in aerial mycelia (Figure 2b), relative to both the WT strain and the CΔAoMbp1 strain. Furthermore, the growth rates of the ΔAoMbp1 deletion strains were significantly lower on TG, TYGA, and PDA media (Figure 2c,d). Additionally, some hyphae were swollen and presented an increased number of mycelial septa (Figure 2e), resulting in shorter mycelial cell lengths (Figure 2f).
3.3. Deletion of AoMbp1 Impairs Conidial Production and Morphology
No significant differences were detected between the CΔAoMbp1 and WT strains. In contrast, the ΔAoMbp1 deletion strain produced fewer conidiophores (Figure 3a) and exhibited a marked reduction in conidial yield compared to both the WT and CΔAoMbp1 strains (Figure 3b). Approximately 84.28% of ΔAoMbp1 conidia displayed abnormal morphology. The length of WT conidia ranged from 18.24 to 27.55 μm (mean 21.31 μm), with a width of 7.38 to 12.07 μm (mean 9.44 μm). In comparison, ΔAoMbp1 conidia were 15.40–63.29 μm in length (mean 33.89 μm) and 4.74–20.29 μm in width (mean 10.39 μm). The length-to-width ratio of ΔAoMbp1 conidia was approximately 1.5 times greater than that of WT conidia, indicating an elongated and slender morphology; some conidia also exhibited incomplete septation (Figure 3c,d). The conidial germination rate of the ΔAoMbp1 strain was significantly lower than those of the WT and CΔAoMbp1 strains (Figure 3e). Furthermore, the hydrophobicity of the conidial surface was significantly reduced in the mutant (Figure 3f).
3.4. AoMbp1 Contributes to Trap Formation, Predatory Capability and Extracellular Protein Hydrolase Expression
Phenotypic analysis revealed significantly reduced trap formation in the ΔAoMbp1 strain compared to the WT and CΔAoMbp1 strains (Figure 4a,b). At 24 h post-nematode induction, the WT strain produced approximately 19.90 traps per cm^2^, while the ΔAoMbp1 mutant formed only 2.48 traps per cm^2^. After 48 h, trap numbers reached 32.64 per cm^2^ in the WT strain compared to only 5.31 per cm^2^ in the mutant strain (Figure 4b). The nematode predation rate of the WT strain was 52.70% at 24 h and increased to 94.92% at 48 h, whereas the ΔAoMbp1 strain exhibited significantly reduced predation rates of only 9.60% and 23.18%, respectively (Figure 4c). In contrast, the CΔAoMbp1 strain showed no significant differences from the WT strain in either trap formation or predatory capacity. Furthermore, the ΔAoMbp1 strain displayed a significantly smaller hydrolysis zone than both the WT and CΔAoMbp1 strains in extracellular protease assays (Figure 4d,e), indicating that AoMbp1 also contributes to regulating the production or secretion of extracellular proteolytic enzymes.
3.5. AoMbp1 Is Required for Stress Resistance and Nutrient Utilization
Under both hyperosmotic and oxidative stress conditions, all tested strains exhibited varying degrees of growth inhibition. The ΔAoMbp1 strain demonstrated significantly enhanced sensitivity to these stresses compared to the WT and CΔAoMbp1 strains. Under hyperosmotic stress conditions, the ΔAoMbp1 mutant showed markedly enhanced growth inhibition (Figure 5a,b) with significantly higher Relative Growth Inhibition (RGI) values (Figure 5b). For instance, the ΔAoMbp1 strain exhibited a 31.02% increase in RGI value under 0.2 M NaCl stress compared to the WT strain. Similarly, the mutant strain displayed pronounced sensitivity to oxidative stress agents, demonstrating substantially elevated RGI values relative to control strains (Figure 5c,d). The ΔAoMbp1 strain showed RGI value increases of 65.87% with 0.05 mmol/L menadione and 66.80% with 10 mmol/L H_2_O_2_ compared to the WT strain. When cultured on media containing sodium acetate, Tween, or oleic acid as carbon sources, the mutant exhibited stronger growth suppression and increased RGI values compared to both WT and complemented strains (Figure 5e,f). The ΔAoMbp1 strain demonstrated a 29.51% increase in RGI value with 0.12% oleate. Under different nitrogen source conditions, the ΔAoMbp1 strain showed significantly elevated RGI values (Figure 5g,h), including a 40.04% increase with yeast extract. These findings demonstrate that deletion of AoMbp1 substantially impairs the ability of A. oligospora to tolerate both hyperosmotic and oxidative stresses while reducing its capacity to utilize various fatty acids and nitrogen sources.
3.6. Transcriptome Analysis of the WT and ΔAoMbp1 Strains
The quality of the transcriptome sequencing data was evaluated (Figure S7a,b). The analysis revealed that 87.23–90.11% of the sequenced reads were successfully aligned to the A. oligospora reference genome. The error rate across all six samples was less than 0.02%, while the Q20 and Q30 quality scores ranged from 95.00% to 95.35%, and the GC content varied between 46.99% and 47.56%. These metrics indicate that the sequencing data were of high quality (Figure S7c). RT–qPCR validation of five DEGs confirmed the reliability of the RNA-seq data. The expression levels of four randomly selected genes (AOL_s00006g570, AOL_s00210g122, AOL_s00054g525, AOL_s00054g29) and AoSho1 (AOL_s00078g396) were all consistent with the transcriptomic results (Figure S7d). Compared with the WT strain, the ΔAoMbp1 mutant exhibited 1964 significantly upregulated and 1985 downregulated genes (Figure 6a and Table S2).
Gene Ontology (GO) enrichment analysis showed that upregulated genes were primarily associated with biosynthetic processes, including those for organonitrogen compound biosynthetic process, organic substance biosynthetic process, cellular macromolecule metabolic process, cellular metabolic process, catalytic activity, among others (Figure 6b). In contrast, downregulated genes were significantly enriched in processes such as cellular responses to chemical stimulus, transmembrane transport, cellular response to chemical stress, asexual reproduction, asexual spore sporulation, conidium formation, integral component of membrane, intrinsic component of membrane, plasma membrane, DNA-binding transcription factor activity, transport activity, transcriptional regulator activity, transmembrane transporter activity, among others (Figure 6c).
KEGG pathway enrichment analysis revealed that the upregulated DEGs were significantly enriched in pathways such as aminoacyl-tRNA biosynthesis, ribosome, nucleotide excision repair, DNA replication, the cell cycle (yeast), mismatch repair, and histidine metabolism (Figure 6d). In comparison, the downregulated DEGs were significantly enriched in pathways such as the mitogen-activated protein kinase (MAPK) signaling pathway, peroxisome pathway, and nitrogen metabolism pathway (Figure 6e).
Using the STRING database, a protein interaction network was constructed to identify proteins potentially functionally related to or directly interacting with AoMbp1. These results suggest that AoMbp1 may play a regulatory role in several key cellular processes, such as the MAPK signaling pathway, ubiquitin-mediated protein hydrolysis, meiosis, autophagy, and cell cycle regulation (Figure 6f). Notably, combined transcriptome analysis revealed several specific proteins, including the mitogen-activated protein kinase Hog1 (AOL_s00109g23), which is linked to the MAPK pathway and mitophagy; the cyclin-dependent kinase Cdc28 (AOL_s00210g359), associated with meiosis, the cell cycle, and the MAPK pathway; and the serine/threonine protein kinase Cdc5 (AOL_s00078g71), involved in cell cycle regulation and meiosis. Additionally, the regulatory subunit B of protein phosphatase PP2A (AOL_s00007g146) was notable for its role in the cell cycle.
3.7. Functional Analysis of DEGs
On the basis of Gene Ontology (GO) analysis, we performed a clustered heatmap analysis of DEGs related to conidium formation (GO: 0048315). The only significantly upregulated gene related to conidium formation in ΔAoMbp1 was Chitin synthase (Chs, AOL_s00079g31), whereas the significantly downregulated genes included six Chs (AOL_s00210g38, AOL_s00078g76, AOL_s00210g143, AOL_s00210g37, AOL_s00075g119, AOL_s00075g153) and StuA (AOL_s00083g25) (Figure 7a). These results suggest that AoMbp1 may play a crucial regulatory role in the sporulation process of A. oligospora by coordinately regulating multiple chitin synthases (Chs).
On the basis of KEGG analysis, we performed a clustered heatmap analysis of DEGs related to peroxisomes (pathway ID: map04146), the MAPK signaling pathway (pathway ID: map04011), and nitrogen metabolism (pathway ID: map00910). Among the DEGs involved in peroxisome synthesis (e.g., Pex and Pxmp4), those genes were significantly lower in the ΔAoMbp1 strain than in the WT strain (Figure 7b). In addition, the expression of antioxidant enzyme-encoding genes involved in the response to oxidative stress (e.g., Cat,* Prdx5*) and genes involved in fatty acid β-oxidation (e.g., AMP-dependent synthase/ligase) were generally downregulated, whereas the expression of the superoxide dismutase gene Sod2 was upregulated (Figure 7b,c), suggesting that AoMbp1 is essential for maintaining normal peroxisome biogenesis, cellular redox homeostasis and fatty acid metabolism.
The Hog1-MAPK pathway, from the upstream sensor Sho1 to the core kinase cascade (Ste11, Pbs2, Hog1) and then to the downstream transcription factor (Msn2) and effector gene (Ctt1), was significantly downregulated (Figure 7d–f). In addition, other key genes in the MAPK pathway, such as Ste12 and Ime2, were also significantly downregulated. Notably, on the basis of the previous GO analysis results, genes whose expression was downregulated in the ΔAoMbp1 deletion strain were significantly enriched in membrane components. Further analysis revealed that multiple genes encoding membrane proteins related to the MAPK pathway (such as Sho1, Msb2, and Wsc) were also significantly downregulated (Figure 7d,e), suggesting that AoMbp1 may regulate the membrane protein-mediated MAPK signaling pathway. AoSho1 (AOL_s00078g396), encoding a membrane protein, was significantly downregulated. Moreover, AoSho1 serves as an upstream sensor of the Hog1-MAPK pathway, and its homologs in yeast may be regulated by Mbp1 [24]. Therefore, AoSho1 was selected as a representative target gene for subsequent verification of AoMbp1 transcriptional regulation.
Furthermore, the expression of those DEGs related to nitrogen metabolism was significantly downregulated in the ΔAoMbp1 deletion strain (Figure 7g). These downregulated genes encoded key enzymes involved in nitrogen assimilation, transport, and catabolism, such as carbonic anhydrase (AOL_s00210g256), nitrate reductase (AOL_s00210g162), glutamate dehydrogenase (AOL_s00080g371), nitrite transporter (AOL_s00004g610), formamidase (AOL_s00043g399), and glutamate synthase (AOL_s00173g269).
3.8. AoMbp1 Directly Binds to the Promoter of the AoSho1 Gene
To experimentally verify the interaction between AoMbp1 and its target promoter, we performed molecular docking, Y1H, and EMSA. Although no typical MluI box (5′-ACGCGT-3′) was detected in the AoSho1 promoter region, a similar sequence (5′-CGCG-3′) was identified (Figure S8a,b). Molecular docking revealed strong binding affinity between AoMbp1 and the 1000bp AoSho1 promoter region, with a confidence score of 0.9456. Visualization using PDBePISA and PyMOL indicated the presence of 10 potential hydrogen bonds at the binding interface (Figure 8a,b). In the Y1H assay, yeast strains co-transformed with pGADT7-AoMbp1 and the proAoSho1-AbAi bait vector grew vigorously on SD/-Leu medium containing 150 or 200 ng/mL aureobasidin A (AbA), whereas negative controls showed no growth (Figure 8c), indicating a specific interaction. EMSA results demonstrated a dose-dependent mobility shift in the AoSho1 promoter fragment upon incubation with increasing concentrations of purified AoMbp1 protein (Figure 9a–d). No shift was observed with bovine serum albumin (BSA) as a nonspecific control (Figure 9e), confirming in vitro binding specificity.
4. Discussions
Taken together, our analyses demonstrate that deletion of AoMbp1 leads to severe phenotypic defects, including restricted mycelial growth, abnormal hyphal morphology, significantly impaired conidiation with aberrant conidial morphology, markedly reduced trap formation, and enhanced sensitivity to osmotic and oxidative stresses. Transcriptome profiling further validated these phenotypic observations, collectively confirming the essential role of AoMbp1 in regulating hyphal development, conidiation, stress adaptation, and pathogenicity in A. oligospora (Figure 10).
Building on the conserved role of Mbp1 as a key regulator of the G1/S transition in yeast, we investigated its function in hyphal morphology and cell cycle regulation in A. oligospora. The ΔAoMbp1 mutant exhibited swollen hyphae, increased septation, and a reduced growth rate, suggesting dysregulation of cell cycle progression, a process known to be essential for septation and hyphal elongation [25]. Transcriptomic analysis revealed significant upregulation of genes involved in fundamental cell cycle-related pathways, including aminoacyl-tRNA biosynthesis, ribosome biogenesis, nucleotide excision repair, DNA replication, cell cycle, and mismatch repair (Figure 6d). Notably, expression of the DNA damage response gene Rnr1 (AOL_s00110g137) increased by 2.48-fold. In yeast, Mbp1 directly binds the Rnr1 promoter to regulate its transcription. These results suggest that AoMbp1 deletion disrupts the G1/S transition and induces DNA replication stress, leading to genomic instability and activation of DNA damage response mechanisms. This diversion of cellular resources toward repair processes likely impairs normal hyphal elongation and coordinated cell division, resulting in aberrant morphology and growth delay. Additionally, these disruptions may indirectly affect conidiation, which requires precise cell cycle control.
In contrast, downregulated genes were enriched in pathways associated with cellular stress responses, such as the MAPK signaling pathway (Figure 6e). The ΔAoMbp1 strain exhibited heightened sensitivity to hyperosmotic stress, correlated with broad downregulation of the Hog1-MAPK signaling pathway (Figure 7f). In A. oligospora, Hog1 acts as a core kinase of the HOG-MAPK pathway, regulating hyperosmotic stress responses, trap morphogenesis, and hyphal development [21,26]. Impaired Hog1 signaling, a central coordinator of osmotic stress responses in fungi, likely underlies this severe sensitivity. Sho1, a conserved membrane sensor, activates the MAPK cascade via the Sho1-Ste11-Pbs2-Hog1 pathway and is critical for fungal development, pathogenicity, and responses to diverse stresses [27,28,29,30]. Our Y1H and EMSA experiments confirmed direct binding of AoMbp1 to the AoSho1 promoter in vitro and in yeast systems. However, it must be noted that the absence of in vivo validation through chromatin immunoprecipitation (ChIP) represents a key limitation of this study. While our data provide strong evidence for this interaction and support a model where AoMbp1 modulates the osmotic stress response by transcriptionally regulating AoSho1, definitive confirmation under native chromatin conditions awaits future investigation.
AoMbp1 is also implicated in oxidative stress response and metabolic regulation, processes crucial for environmental adaptation and trap development. The ΔAoMbp1 mutant showed increased sensitivity to oxidants and impaired growth under nutrient stress. Transcriptome analysis revealed significant downregulation of genes related to Peroxisome and nitrogen metabolism. Expression of the downstream transcription factor AoMsn2, which regulates oxidative stress responses and lipid metabolism, was also reduced [31]. In yeast, Pex34 interacts with Pex11, Pex25, and Pex2 to regulate peroxisome proliferation [32]. Similarly, in A. oligospora, Pex1 and Pex6 are associated with hyphal growth and trap formation [20], while Pex14/17 influences development and secondary metabolism [33]. Given the well-established roles of peroxisomes in β-oxidation and ROS detoxification, and their notable enrichment in trap structures, we hypothesize that the observed downregulation of peroxisome-related genes in ΔAoMbp1 impairs peroxisomal function. This dysfunction likely reduces cellular ROS scavenging capacity and disrupts fatty acid metabolism, which collectively contributes to the growth deficiencies and increased oxidative stress sensitivity observed in the mutant.
Deletion of AoMbp1 also markedly reduced conidial surface hydrophobicity, induced morphological abnormalities including defective septa, and significantly decreased spore production. Transcriptome analysis revealed that AoMbp1 deletion significantly downregulated the expression of chitin synthase (Chs) genes and RodA (AOL_s00006g570). In Aspergillus fumigatus, RodA is a key hydrophobin critical for spore formation, surface permeability, hydrophobicity, and immune evasion [34]. Coordinated expression of Chs family members is essential for fungal morphology [35,36]. For instance, in Aspergillus nidulans, specific chitin synthases are required for septal chitin synthesis, a major structural component of fungal cell walls and septa [37]. Therefore, AoMbp1 likely coordinates chitin synthesis and distribution through regulating Chs genes and RodA, thereby influencing conidial septum formation, morphology, and environmental adaptation.
The pleiotropic effects observed in the AoMbp1 deletion mutant originate from the integrated impact of direct transcriptional regulation and indirect secondary consequences. Molecular characterization identified AoSho1 as a direct transcriptional target, establishing a definitive mechanistic link for AoMbp1-mediated regulation of stress sensitivity and trap formation through the MAPK pathway. In contrast, the broader developmental and growth impairments appear to stem from secondary effects that manifest as functional disruptions across three distinct levels. Developmental processes including asexual reproduction, sporulation, and conidial formation show significant defects, while metabolic functions exhibit systematic dysregulation in organonitrogen compound biosynthesis, macromolecular metabolism, and transmembrane transport. Concurrently, stress response capabilities demonstrate substantially diminished capacity to counter chemical and environmental challenges. These functional deficiencies find molecular correlation through coordinated downregulation of core pathways including MAPK signaling, peroxisome function, and nitrogen metabolism. This transcriptional reprogramming is accompanied by broad suppression of key molecular functions, particularly transmembrane transporter activity and DNA-binding transcription factor activity. Furthermore, fundamental cellular processes involving aminoacyl-tRNA biosynthesis, ribosomal function, DNA replication, and repair mechanisms are significantly compromised.
Building upon these comprehensive findings, we propose an integrated “Offense-Defense-Metabolism” model wherein AoMbp1 serves as a master transcriptional coordinator. During the ecological transition from saprophytism to predation, elevated AoMbp1 expression orchestrates the coordinated regulation of genes involved in membrane composition, MAPK signaling, peroxisome biogenesis, fatty acid degradation, and nitrogen metabolism. Within the offense module, A. oligospora detects nematode-derived signals through membrane-localized GPCRs, transducing these signals to activate the intracellular MAPK pathway, with AoMbp1 facilitating trap morphogenesis and promoting active infection through regulation of key signaling components. The defense module features AoMbp1-mediated enhancement of fungal tolerance to nematode-derived oxidative compounds via modulation of peroxisome biogenesis and associated functional genes, ensuring predation success through improved oxidative stress resistance. In the metabolism module, post-capture nutrient utilization is optimized through AoMbp1 coordination of fatty acid β-oxidation and nitrogen metabolic pathways, enabling efficient energy allocation during predation. Collectively, our results demonstrate that the APSES transcription factor AoMbp1 in A. oligospora has functionally diverged from its canonical cell cycle regulatory role in S. cerevisiae, representing an evolutionary adaptation to complex soil environments that enables the fungal transition from saprophytic to predatory existence through sophisticated integration of developmental, stress adaptation, and metabolic processes.
The pleiotropic role of AoMbp1 in regulating conidiation, trap formation, and stress tolerance highlights its potential as a target for genetic strain improvement. This underscores the ability of master transcriptional regulators to reprogram fungal adaptation, as exemplified by the Zn_2_Cys_6_ transcription factor CtBOT6, whose overexpression activates the ABA-BOT gene cluster and converts the beneficial endophyte Colletotrichum tofieldiae into a potent pathogen [38]. Consequently, targeted modulation of AoMbp1 expression, for example, through controlled overexpression, could generate A. oligospora strains with enhanced fitness under field conditions. Such strains are anticipated to exhibit improved conidial robustness, accelerated trap formation, and greater tolerance to environmental stresses common in agricultural soils. The identification of the AoMbp1–AoSho1 regulatory axis provides a genetic framework and potential molecular markers, such as AoSho1 expression levels, for tracking the performance of engineered strains. Thus, this work not only establishes a foundation for developing more effective biocontrol agents but also advances our understanding of how transcriptional circuits govern ecological strategies in fungi.
5. Conclusions
This study demonstrated that the APSES family protein AoMbp1 is a global transcription factor that plays critical roles in regulating oxidative stress, hyperosmotic stress, pathogenicity, trap formation, mycelial growth, and conidial formation by modulating downstream target genes, such as AoSho1. These findings not only provide new insights into the regulatory mechanisms of the APSES family of transcription factors in A. oligospora but also lay a theoretical foundation for the development of genetically modified A. oligospora.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Freitas L.A. Savegnago R.P. Menegatto L.S. Bem R.D.D. Stafuzza N.B. Paz A. Pires B.V. Costa R. Paz C.C.P. Cluster analysis to explore additive-genetic patterns for the identification of sheep resistant, resilient and susceptible to gastrointestinal nematodes Vet. Parasitol.202230110964010.1016/j.vetpar.2021.10964034973595 · doi ↗ · pubmed ↗
- 2Besier R.B. Kahn L.P. Sargison N.D. Van Wyk J.A. The Pathophysiology, Ecology and Epidemiology of Haemonchus contortus Infection in Small Ruminants Adv. Parasitol.2016939514310.1016/bs.apar.2016.02.02227238004 · doi ↗ · pubmed ↗
- 3Li S. Wang D. Gong J. Zhang Y. Individual and Combined Application of Nematophagous Fungi as Biological Control Agents against Gastrointestinal Nematodes in Domestic Animals Pathogens 20221117210.3390/pathogens 1102017235215117 PMC 8879429 · doi ↗ · pubmed ↗
- 4Hou B. Yong R. Wuen J. Zhang Y. Buyin B. Subu D. Zha H. Li H. Hasi S. Positivity Rate Investigation and Anthelmintic Resistance Analysis of Gastrointestinal Nematodes in Sheep and Cattle in Ordos, China Animals 20221289110.3390/ani 1207089135405881 PMC 8997026 · doi ↗ · pubmed ↗
- 5Kaplan R.M. Biology, Epidemiology, Diagnosis, and Management of Anthelmintic Resistance in Gastrointestinal Nematodes of Livestock Vet. Clin. N. Am. Food Anim. Pract.202036173010.1016/j.cvfa.2019.12.00132029182 · doi ↗ · pubmed ↗
- 6Zhou D. Xu J. Dong J. Li H. Wang D. Gu J. Zhang K.Q. Zhang Y. Historical Differentiation and Recent Hybridization in Natural Populations of the Nematode-Trapping Fungus Arthrobotrys oligospora in China Microorganisms 2021991910.3390/microorganisms 909191934576814 PMC 8465350 · doi ↗ · pubmed ↗
- 7Wang D. Ma N. Rao W. Zhang Y. Recent Advances in Life History Transition with Nematode-Trapping Fungus Arthrobotrys oligospora and Its Application in Sustainable Agriculture Pathogens 20231236710.3390/pathogens 1203036736986289 PMC 10056792 · doi ↗ · pubmed ↗
- 8Szewc M. De Waal T. Zintl A. Biological methods for the control of gastrointestinal nematodes Vet. J.202126810560210.1016/j.tvjl.2020.10560233468301 · doi ↗ · pubmed ↗