Scaffold-Free Bone Regeneration Through Collaboration Between Type IV Collagen and FBXL14
Mari Akiyama

TL;DR
This study explores how type IV collagen and FBXL14 work together to regenerate bone without a scaffold, using cells from bovine periosteum.
Contribution
The study reveals a novel collaboration between type IV collagen and FBXL14 in scaffold-free bone regeneration.
Findings
Type IV collagen and FBXL14 were expressed in Volkmann’s and Haversian canals in bone and periosteum.
After 5 weeks, these proteins surrounded calcified crystals containing osteocalcin and formed periosteum-derived cells.
Von Kossa and osteocalcin staining confirmed the presence of calcified substances in the crystals.
Abstract
Background: The periosteum and periosteum-derived cells have attracted considerable attention for their potential use in clinical applications for treating bone defects. Bovine periosteum-derived cells have been investigated because of their capability for scaffold-free bone regeneration. Previous mass spectrometry (MS) and immunohistochemistry studies have shown the presence of F-box/leucine-rich repeat protein 14 (FBXL14) in bovine periosteum and periosteum-derived cells. Recently, studies using ESI-Q-Orbitrap MS suggested the presence of type IV collagen in the periosteum. The aim of the present study was to clarify the relationship between type IV collagen and FBXL14 in the formation of periosteum-derived cells. Methods: Bovine periosteum-derived cells were obtained from Japanese Black Cattle’s legs in Medium 199 with ascorbic acid and 10% fetal bovine serum. Immunohistochemistry…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1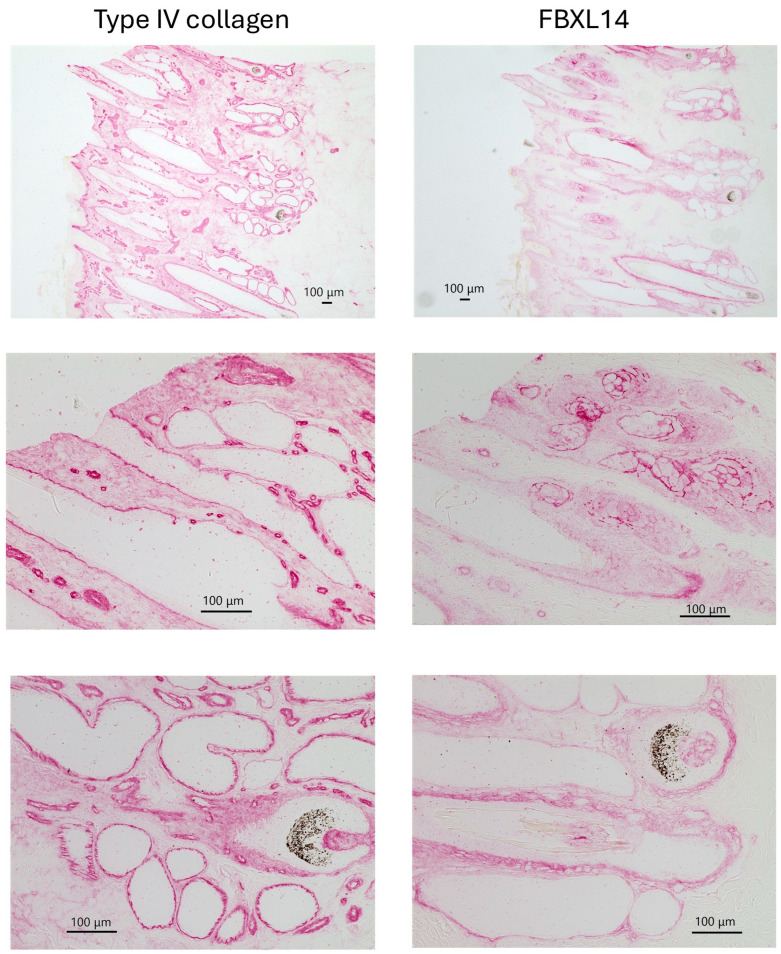 Figure 2
Figure 2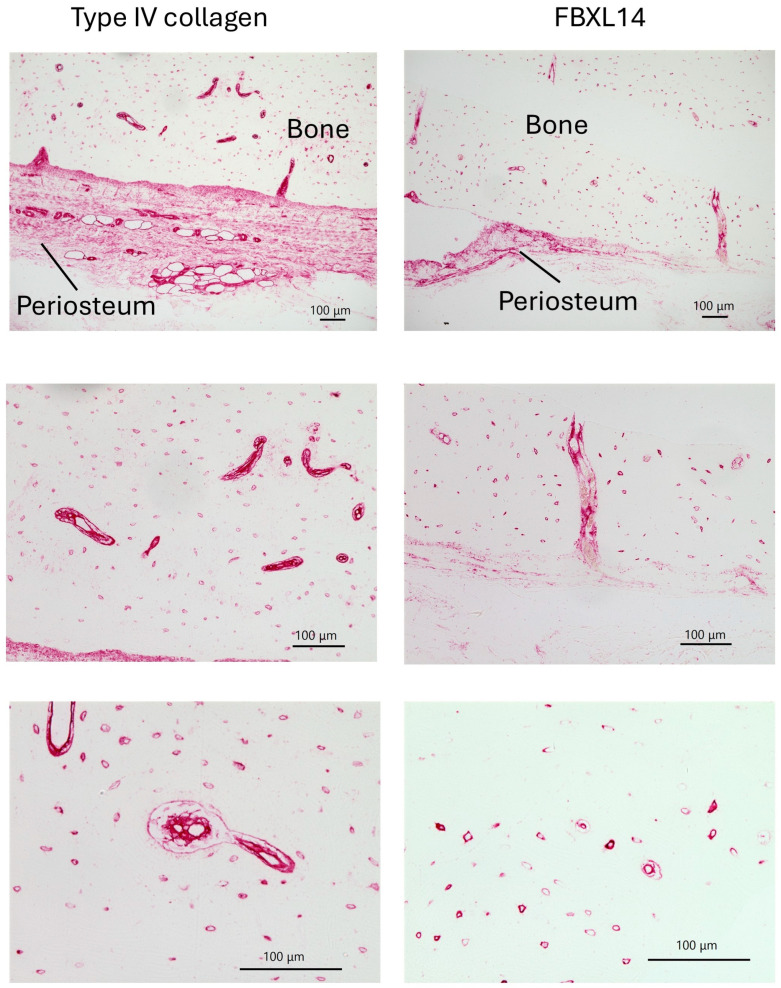 Figure 3
Figure 3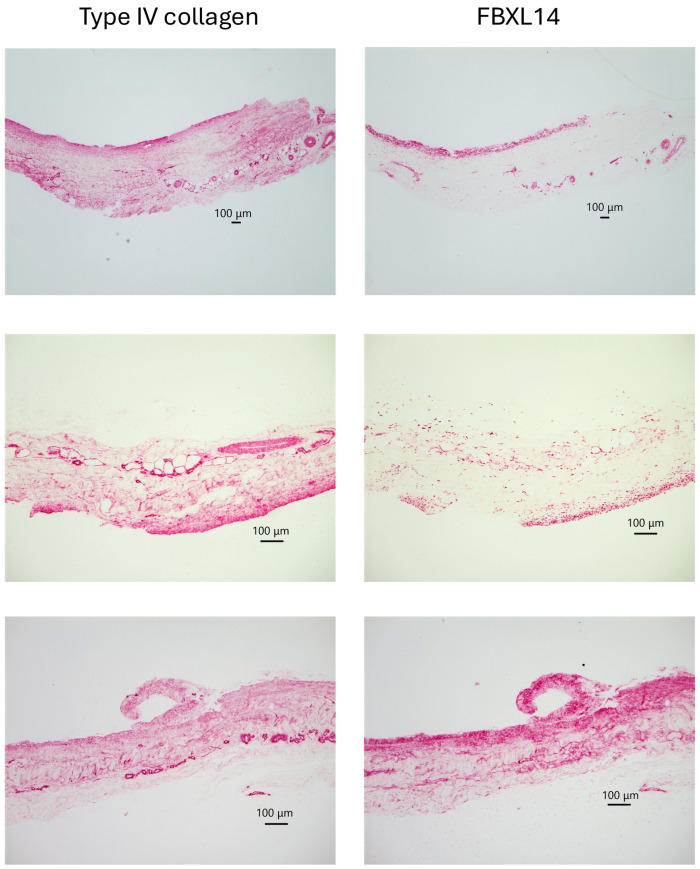 Figure 4
Figure 4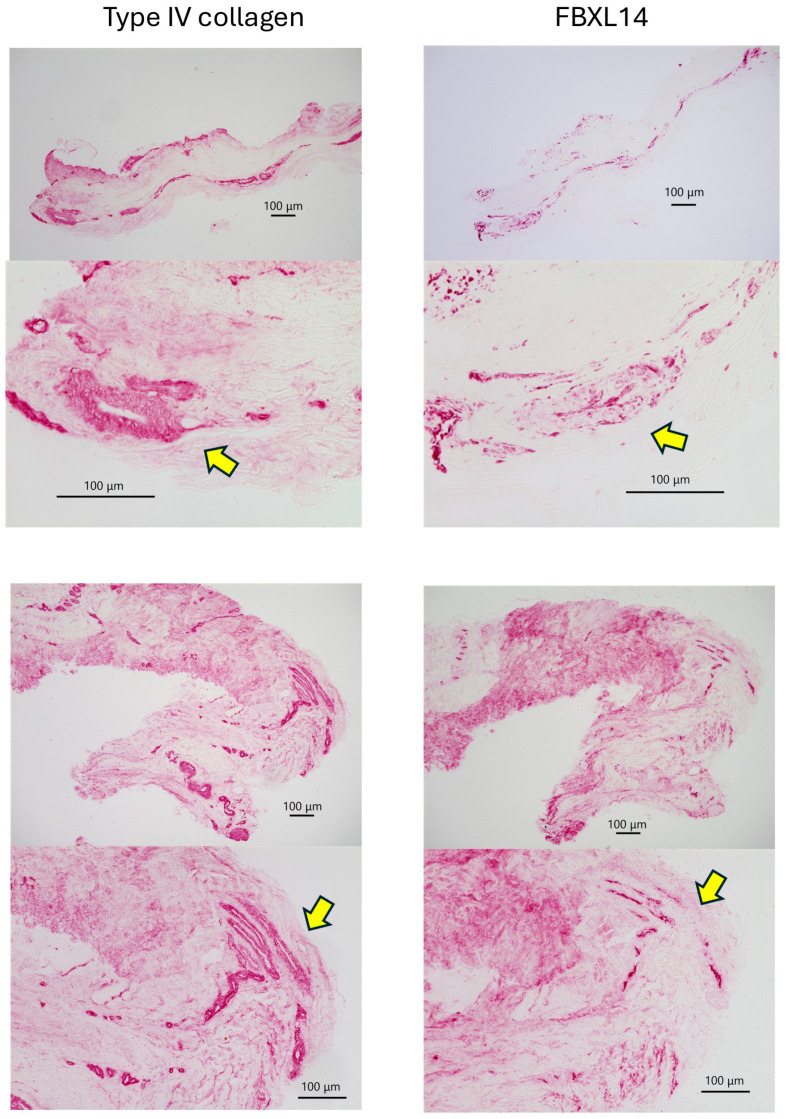 Figure 5
Figure 5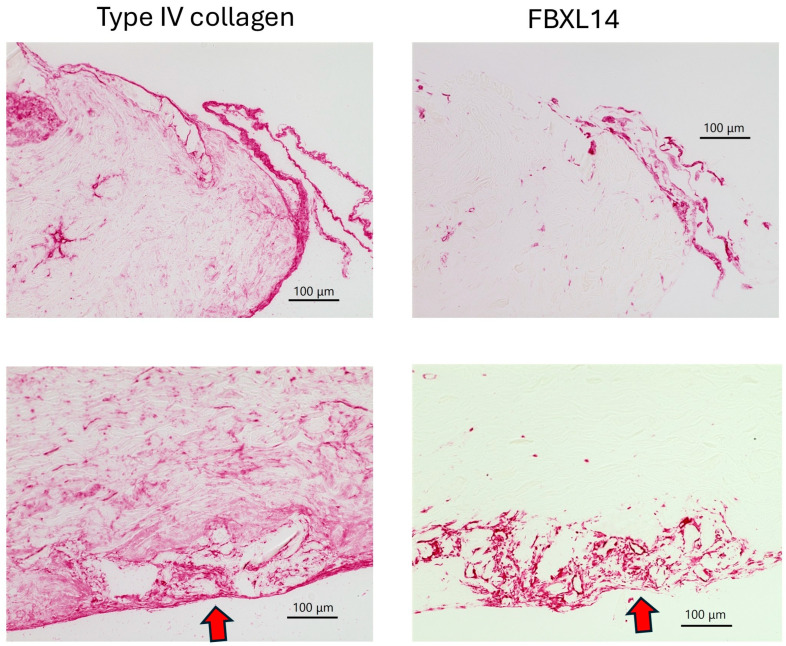 Figure 6
Figure 6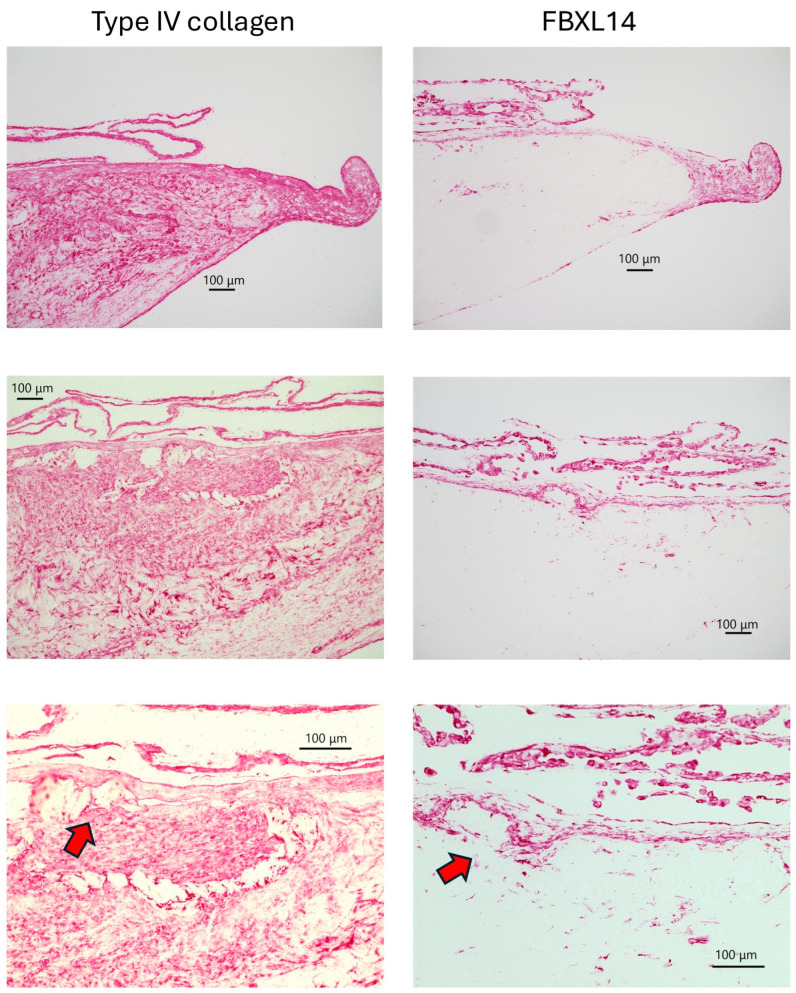 Figure 7
Figure 7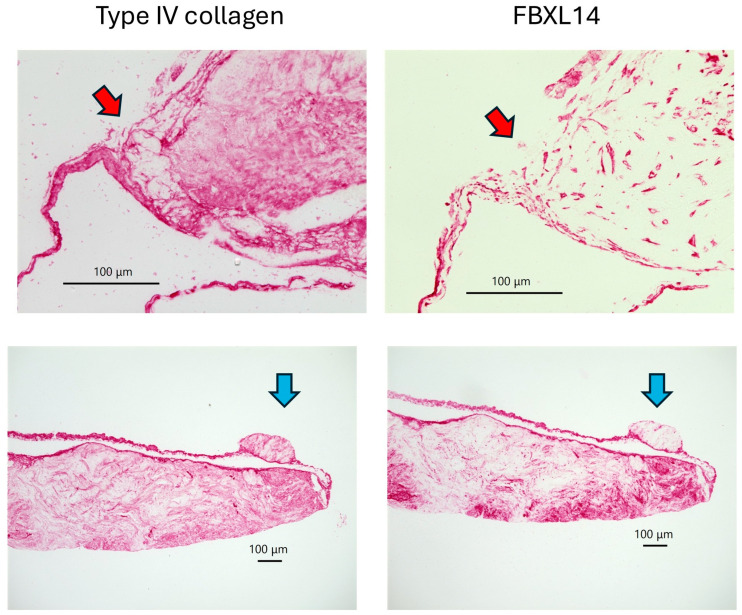 Figure 8
Figure 8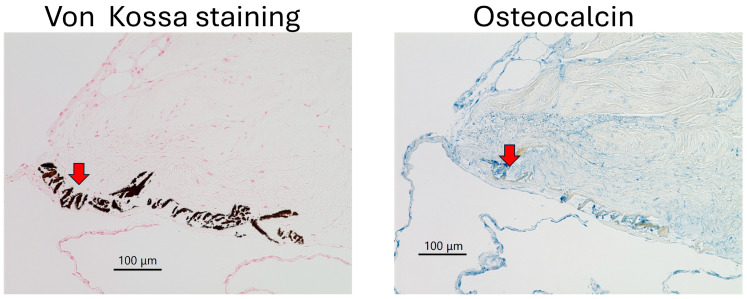 Figure 9
Figure 9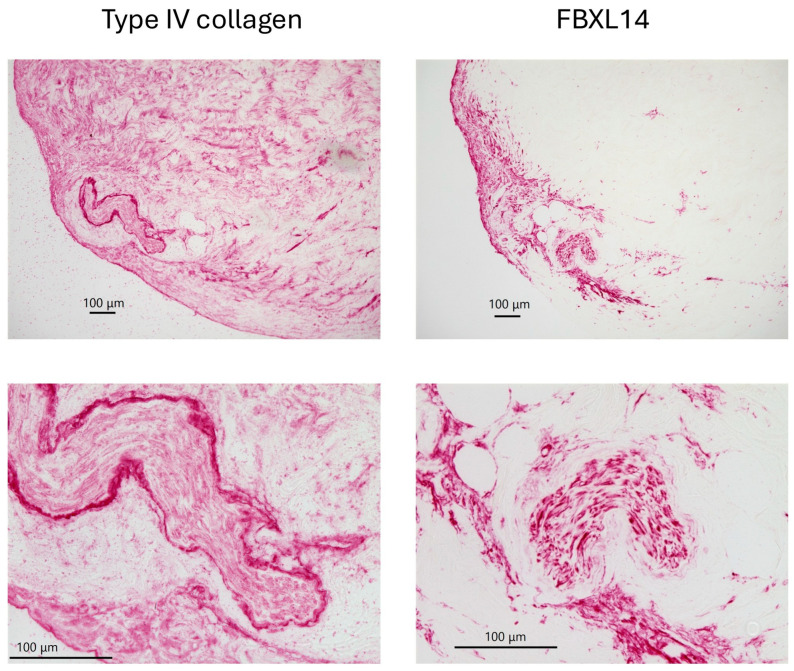 Figure 10
Figure 10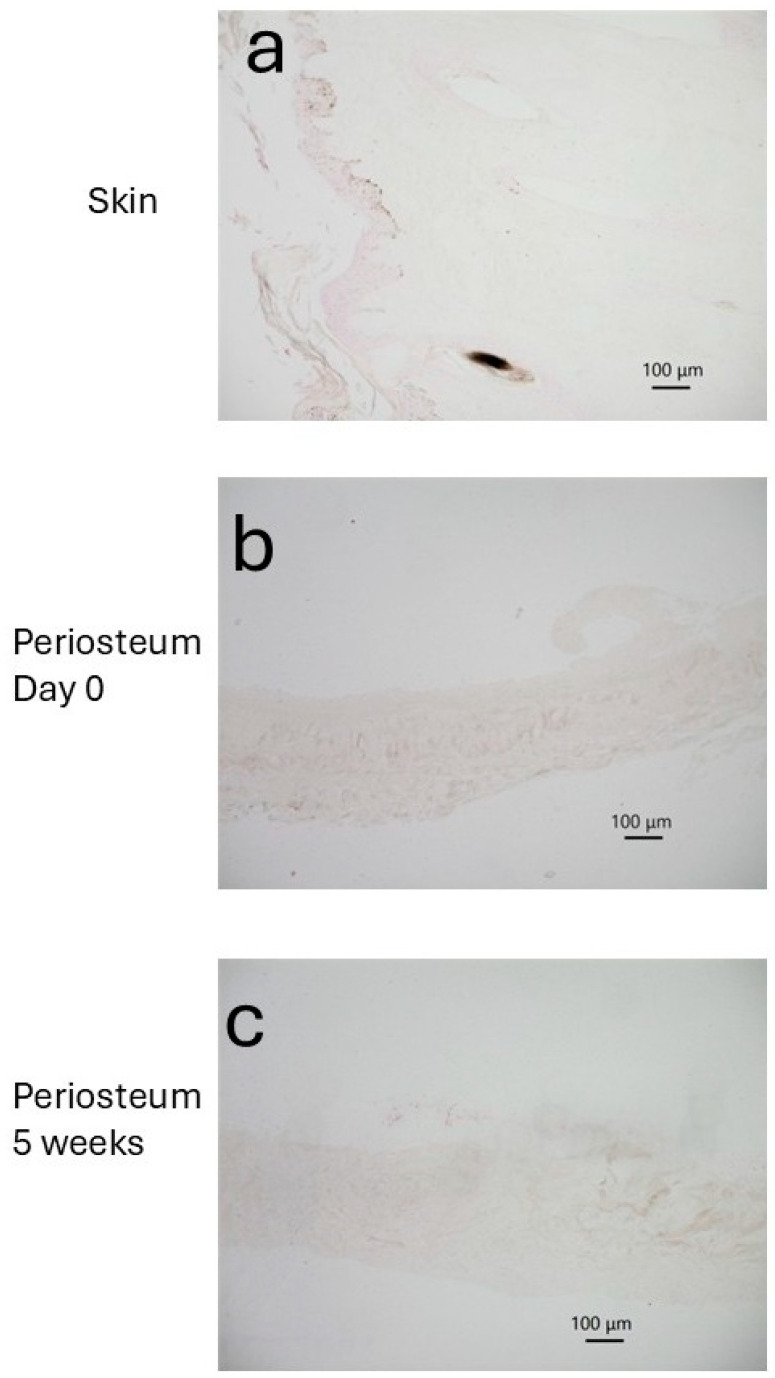 Figure 11
Figure 11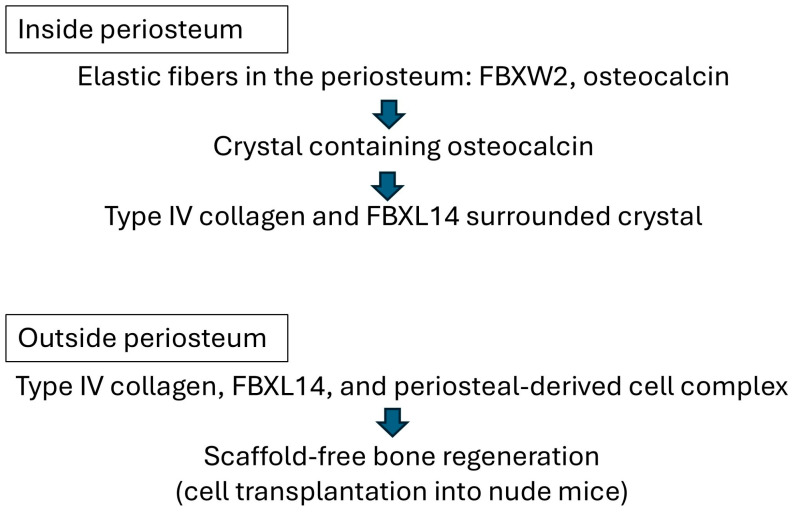 Figure 12
Figure 12- —JSPS KAKENHI
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsElectrospun Nanofibers in Biomedical Applications · Connective tissue disorders research · Periodontal Regeneration and Treatments
1. Introduction
The periosteum and periosteum-derived cells (PDCs) have attracted attention for clinical use in bone regeneration due to their osteogenic ability [1,2,3]. For tissue engineering, the periosteum and PDCs are often used with artificial materials [4,5]. There are two methods of culturing PDCs: collagenase digestion [1,6] and explant culture methods [7]. Because periosteum contains osteogenic stem cells [8], PDCs are periosteum-derived stem cells identified using stem cell markers [2]. In clinical use, PDCs are used for guided bone regeneration [3] with artificial synthetic scaffolds. However, these scaffolds have problems of rejection, shrinkage, or, quite the opposite, remain for a long time. Therefore, biomimetic scaffold-free techniques have been developed [9,10]. Bovine PDCs in the absence of any artificial material can form a multilayer structure made from extracellular matrix (ECM) in vitro (Figure 1), and scaffold-free PDCs resulted in bone regeneration in vivo when grafted into the backs of nude mice [11,12,13]. The molecular signaling of the periosteum involved in bone healing has also been investigated [1,14,15,16].
To clarify the osteogenic mechanism of protein interactions, we previously performed mass spectrometry (MS) and immunohistochemistry [17] and found that F-box/leucine-rich repeat protein 14 (FBXL14) supported the formation of multilayered PDCs [13,18]. FBXL14, an F-box protein, regulates the epithelial-to-mesenchymal transition with target proteins during development [19] and the progression of atherosclerotic plaque via the NRF2 signal pathway through ubiquitination of bispecific phosphatase-6 [20]. Osteocalcin is a vitamin K-dependent carboxylated protein that is related to osteoporosis [21,22,23,24]. Undercarboxylated osteocalcin is also related to whole-body metabolism as a hormone [25]. In 2018, Akiyama [26] reported that fibers of F-box and WD-40 domain-containing protein 2 (FBXW2) were coated with osteocalcin in the periosteum (Table S1). In the periosteum, osteocalcin was assembled into crystal form without FBXW2 fibers [26]. These osteocalcin-containing crystals were identified by using double immunostaining methods in previous studies [26,27]. Electrospray ionization (ESI)-Q-Orbitrap MS suggested the presence of a basement membrane component, type IV collagen, with over 890 proteins in the supernatant of PDCs [28] (Table S1), whereas electrospray ionization quadrupole time-of-flight (ESI-Q-TOF) MS/MS analysis was unable to detect type IV collagen [17]. Type IV collagen plays an important role in peripheral nerve myelin in regulating Schwann cells [29,30,31] and capillaries as a permeability selective barrier [32], and mutations in type IV collagen-related genes can cause disorders such as Alport syndrome [33,34,35]. Not only type IV collagen as the capillary basement membrane, but FBXL14 is involved in the capillaries of the periosteum [18] based on their presence in supernatants of periosteum using ESI-Q-TOF MS/MS analysis (Table S1). The aim of the present study was to compare and clarify the relationship between type IV collagen and FBXL14 within the ECM of PDCs for bone regeneration.
2. Materials and Methods
Bovine skin, bone, and periosteum were obtained from the legs of 30-month-old Japanese Black Cattle (female and steer, Kobe Chuo Chikusan, Kobe, Japan) within 24 h after death for beef product, according to the Osaka Dental University Regulations on Animal Care and Use (Approval No. 25-02002). Six bovine legs were chosen randomly by the staff of a slaughterhouse; the researcher could not enter the slaughterhouse. This study did not involve living animals. All tissues were fixed with 4% paraformaldehyde. Bone with periosteum from two legs was cut with an Er:Yag Laser Device Erwin Adverl MEY-1 (pulse repetition; 25 pps, energy/pulse; 70 mJ, Morita Corp., Osaka, Japan), and bone was broken with a hammer. Bone and periosteum were demineralized with 10% ethylenediaminetetraacetic-2Na for 8 days at room temperature after fixation for sectioning. The periosteum from six bovine legs was removed with aseptic methods and used for explant culture, as previously described [18]. Bovine legs were disinfected with 10% povidone-iodine (iNova Pharmaceuticals Japan, Tokyo, Japan) and 10% sodium hypochlorite solution (Antiformin, FIJIFILM Wako Pure Chemical Corporation, Osaka, Japan). Approximately 5 mm^2^ periosteal pieces were cut using a disposable scalpel (No. 10, Feather Safety Razor Co., Ltd., Osaka, Japan) and put in a 100 mm-diameter culture dish (353003, FALCON, Corning Incorporated-Life Sciences, Durham, NC, USA) with 5 mg/mL ascorbic acid (A0276-25G Sigma-Aldrich-Merck Japan, Tokyo, Japan), penicillin/streptomycin (168-23191; FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan), and 10% fetal bovine serum (Biosera, Cholet, France) in Medium 199 at 37 °C and 5% CO_2_ in an incubator and cultured for 5 weeks. The culture medium was changed once a week. The skin was prepared as a positive control for the basement membrane. All tissue samples were embedded in paraffin blocks and sectioned at a thickness of 2 μm for immunohistochemistry and von Kossa staining.
In accordance with datasets from ESI-Q-Orbitrap MS (JPST003206, JPST003258) in jPOSTrepo (Japan ProteOme STandard Repository), the presence of type IV collagen was suggested [28]. The activity of antigens in sections was retrieved using Proteinase K (Dako Cytomation, Glostrup, Denmark) for 10 min at room temperature. Anti-collagen Type IV (rabbit) antibody-600-401-1065 (1:500, 1 h; Rockland Immunochemicals, Inc., Rockland County, NY, USA) and anti-FBXL14 (rabbit) antibody (1:500 4 h; #SAB2103691; Sigma-Aldrich, Saint Louis, MO, USA) were used as the primary antibodies. Anti-EXOSC9 (1:500, 4 h; H-300; #sc-135118; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) (rabbit) antibody was used as the negative control. Alkaline phosphatase-conjugated goat anti-rabbit IgG(H+L) (1:200, 1 h; Proteintech Group, Inc., Rosemont, IL, USA) was used as the secondary antibody. All sections were visualized using PermaRed/AP (K049; Diagnostic BioSystems, Pleasanton, CA, USA). Images were obtained using a BZ-X810 microscope (Keyence Japan, Osaka, Japan) and X800 Viewer (Keyence). X800 Analyzer software (Keyence) was used to insert scale bars and increase the contrast. All images are preserved at the repository site GakuNin RDM (https://rdm.nii.ac.jp/)
To clarify the crystal components in the periosteum after 5 weeks, immunohistochemistry using antibody for osteocalcin (1:500, overnight; code no. M042, clone no. OCG2; Takara Bio Inc., Kusatsu, Japan), alkaline phosphatase-conjugated goat anti-mouse IgG(H+L) (1:200, 1 h; Proteintech Group), and PermaBlue/AP (K058; Diagnostic BioSystems) were used. From 2015 to 2017, double immunostaining of osteocalcin and other proteins was performed [26]. In previous studies, antibody for osteocalcin (1:500, 4 h; code no. M042, clone no. OCG2; Takara Bio Inc.) was used. N-Histofine Simple Stain AP (M) (#414241, Nichirei Biosciences Inc., Tokyo, Japan) was used as a secondary antibody. Rabbit uveal autoantigen with coiled-coil domains and ankyrin repeats (UACA) antibody (#bc-6308R; Bioss Inc., Woburn, MA, USA), N-Histofine Simple Stain AP (R) (#414251, Nichirei Biosciences Inc.), and PermaRed/AP were used for double immunostaining for UACA.
3. Results
3.1. Skin
Immunohistochemistry showed that type IV collagen was expressed strongly, not only in the basement membrane but also in microvessels, dermal papillae, and sweat glands (Figure 2, left). In contrast, FBXL14 was expressed strongly in sebaceous glands and the outer root sheath, but weakly in microvessels (Figure 2, right).
3.2. Bone and Periosteum
Type IV collagen and FBXL14 were expressed in the periosteum (Figure 3). In bone, type IV collagen and FBXL14 were expressed in Volkmann’s and Haversian canals. Expression of FBXL14 in Volkmann’s canals containing erythrocytes indicated the presence of FBXL14 in the capillaries (Figure 3, right, middle). However, in comparison with FBXL14, type IV collagen was expressed more extensively and strongly in capillaries because it was more abundant (Figure 3, middle, bottom).
3.3. Periosteum Explant Culture on Day 0
Figure 4 and Figure 5 show a comparison of type IV collagen and FBXL14 expressions in the periosteum on day 0. Figure 4 shows low-magnification images of the periosteum. Type IV collagen was expressed strongly in capillaries and widely throughout the periosteum. FBXL14 was mainly expressed in a single layer of the periosteum. Periosteum was divided into two layers, the cambium layer and the fibrous layer. The layer that strongly expressed type IV collagen and FBXL14 was the cambium layer, near bone (Figure 3 and Figure 4). Figure 5 shows low- and high-magnification images of the periosteum. In Figure 5, a fine mesh of type IV collagen surrounded the capillary wall, whereas FBXL14 was expressed partially within the capillary wall.
3.4. Periosteum Explant Culture at 5 Weeks
After 5 weeks, type IV collagen and FBXL14 formed a matrix in which the PDCs were supported. Type IV collagen surrounded crystals, and FBXL14-positive cells were assembled around these crystals (Figure 6, Figure 7 and Figure 8, red arrows). Type IV collagen and the assembled FBXL14-positive cells formed a membrane around crystals and the complex of PDCs, and the ECM appeared to flake off from the periosteum (Figure 6, Figure 7 and Figure 8, red arrows). As shown in the bottom images of Figure 8, part of the periosteum within the ECM had shifted. Figure 9 shows a comparison of von Kossa staining and immunostaining of osteocalcin. The crystals are calcified and contain osteocalcin. Figure 10 shows a fibrous structure in the periosteum. Type IV collagen surrounded this fibrous structure and was expressed strongly, whereas FBXL14 was expressed in the fibrous structure inside the type IV collagen membrane. Immunohistochemistry indicated the presence of a translucent membrane around FBXL14 (Figure 10, bottom, right).
Figure 11 shows negative controls using anti-EXOSC9 antibody. The skin and periosteum (on day 0 and at 5 weeks) did not exhibit any anti-EXOSC9 antibody reaction.
4. Discussion
The present study first aimed to clarify whether FBXL14 is a basement membrane component like type IV collagen, using immunohistochemistry for type IV collagen as a positive control. Skin was used for basement membrane detection. Type IV collagen is present in the basement membrane of capillaries [35] and hair follicles [36]. The expression patterns of type IV collagen and FBXL14 differ in skin (Figure 2). The periosteum contains sensory nerves [37], and type IV collagen is an important component in the myelin of peripheral nerves [29,30,31]. As shown in Figure 10, type IV collagen and FBXL14 were clearly expressed in different regions of the fibrous structure in the periosteum. S-100 protein, a nerve marker, has also been shown to be present in different regions of nerve tissue from type IV collagen [38]. However, without nerve markers, such as S-100 protein, it cannot be determined, but the fibrous structure in Figure 10 thus could be a sensory nerve.
In bone, capillaries, and the periosteum, type IV collagen and FBXL14 were expressed in similar regions, although FBXL14 expression was weaker. There is an inner cambium layer between bone and the periosteum [39]. As shown in Figure 3, type IV collagen and FBXL14 were expressed strongly in the cambium layer near bone. The periosteum is divided into two layers: the cambium layer (with bone) and the fibrous layer (detached from bone) [1]. The cambium layer could be the region in which type IV collagen and FBXL14 are strongly expressed in Figure 4, which shows the periosteum without bone. As shown in Figure 5, FBXL14 expression in capillaries was much weaker than that of type IV collagen, suggesting that type IV collagen was located outside areas in which FBXL14 was expressed. In this study, two rabbit primary antibodies, anti-collagen type IV antibody (1:500, 1 h) and anti-FBXL14 antibody (1:500 4 h), were used. Anti-EXOSC9 (1:500 4 h) antibody was used for negative controls because anti-EXOSC9 antibody is also a rabbit primary antibody and proves the absence of non-specific staining.
Type IV collagen and FBXL14 were involved in the formation of PDCs over the course of the 5-week explant culture in the present study. First, type IV collagen surrounded crystals. Next, FBXL14 formed a multilayered complex of ECM and PDCs with type IV collagen inside the periosteum. Finally, PDCs appeared to flake off outside the periosteum. In this study, crystal structures were observed in four of six cows (Table 1). In cow 2, no crystal structure was observed because the sections were thin, and these thin sections did not include crystal structures. In cow 6, PDCs were not observed. These differences were observed because the bovine legs were chosen randomly without any standard conditions (such as temperature) in the slaughterhouse. In this case, periosteal cells inside tissue became weak. According to a report by Akiyama [26], the crystal in the periosteum contains osteocalcin (Figure 9, right), which regulates bone mineralization [40]. Figure 9 and Supplementary Figure S1 show the crystal is possibly a complex of calcium salt and osteocalcin. Osteocalcin binds to calcium phosphate [41]. During the 5 weeks of explant culture, osteocalcin likely bound to calcified crystals in the periosteum. In a previous study, treatment with anti-osteocalcin antibody was for 4 h [26], whereas in the present study, it was overnight, because the secondary antibody and the lot number of the primary antibody were different. The results of the present study suggest that osteocalcin is also related to the synthesis of the ECM. In the periosteum, Wnt signaling [14], Dickkopf-related protein 3 [15], the E3 ubiquitin ligase, and the Cbl–phosphatidylinositol-3 kinase interaction [16] are important for bone regeneration. Type I collagen and osteocalcin, a non-collagenous protein, reportedly interact during bone metabolism [42]. Types I and III collagens were abundant in the ECM of PDCs [17]. The supporting results of the previous and present studies indicate that collagen types I, III, and IV and FBXL14 interact during the formation of multilayered PDCs (Figure 12).
Previous studies using ESI-Q-TOF MS/MS [17] and ESI-Q-Orbitrap MS [25] investigated the supernatants of PDCs. ESI-Q-TOF MS/MS was able to detect a few proteins associated with type I and III collagens, but not type IV collagen [17]. ESI-Q-Orbitrap MS detected over 890 proteins associated with type IV collagen [28]. However, the numerous proteins identified by ESI-Q-Orbitrap MS made it difficult to understand which proteins actually affect bone regeneration. The combination of immunohistochemistry and MS methods has revealed interactions between collagen and non-collagenous F-box proteins, but the sensitivity and precision of the antibodies used for the immunohistochemistry method were limited. In the future, more detailed investigations using mass spectrometry imaging will be needed to further clarify the interactions between collagen and non-collagenous proteins.
5. Conclusions
FBXL14 cannot be considered a basement membrane protein. However, type IV collagen and FBXL14 interact with osteocalcin during the formation of PDCs. Clinically, understanding osteocalcin-interacting proteins will help clarify osteoporosis. Biomimetics using native ECM including collagen types IV and FBXL14 may be useful for novel scaffold-free methods in tissue engineering.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lin Z. Fateh A. Salem D.M. Intini G. Periosteum: Biology and applications in craniofacial bone regeneration J. Dent. Res.20149310911610.1177/002203451350644524088412 PMC 3895334 · doi ↗ · pubmed ↗
- 2Zhang X. Deng C. Qi S. Periosteum Containing Implicit Stem Cells: A Progressive Source of Inspiration for Bone Tissue Regeneration Int. J. Mol. Sci.202425216210.3390/ijms 2504216238396834 PMC 10889827 · doi ↗ · pubmed ↗
- 3Zhang W. Wang N. Yang M. Sun T. Zhang J. Zhao Y. Huo N. Li Z. Periosteum and development of the tissue-engineered periosteum for guided bone regeneration J. Orthop. Transl.202233415410.1016/j.jot.2022.01.00235228996 PMC 8858911 · doi ↗ · pubmed ↗
- 4Zhong J. Li W. Li H. Zhang J. Hou Z. Wang X. Zhou E. Lu K. Zhuang W. Sang H. A self-forming bone membrane generated by periosteum-derived stem cell spheroids enhances the repair of bone defects Acta Biomater.202519318520110.1016/j.actbio.2024.12.05839742905 · doi ↗ · pubmed ↗
- 5Ringe J. Leinhase I. Stich S. Loch A. Neumann K. Haisch A. Häupl T. Manz R. Kaps C. Sittinger M. Human mastoid periosteum-derived stem cells: Promising candidates for skeletal tissue engineering J. Tissue Eng. Regen. Med.2008213614610.1002/term.7518383554 · doi ↗ · pubmed ↗
- 6Danalache M. Kliesch S.M. Munz M. Naros A. Reinert S. Alexander D. Quality Analysis of Minerals Formed by Jaw Periosteal Cells under Different Culture Conditions Int. J. Mol. Sci.201920419310.3390/ijms 2017419331461878 PMC 6747376 · doi ↗ · pubmed ↗
- 7Uematsu K. Ushiki T. Ishiguro H. Ohashi R. Tamura S. Watanabe M. Fujimoto Y. Nagata M. Ajioka Y. Kawase T. Osteoclastogenic Potential of Tissue-Engineered Periosteal Sheet: Effects of Culture Media on the Ability to Recruit Osteoclast Precursors Int. J. Mol. Sci.202122216910.3390/ijms 2204216933671612 PMC 7926432 · doi ↗ · pubmed ↗
- 8Hsiao H.Y. Yang C.Y. Liu J.W. Brey E.M. Cheng M.H. Periosteal Osteogenic Capacity Depends on Tissue Source Tissue Eng. Part A 2018241733174110.1089/ten.tea.2018.000929901423 · doi ↗ · pubmed ↗