Silencing the cyp314a1 and cyp315a1 Genes in the Aedes albopictus 20E Synthetic Pathway for Mosquito Control and Assessing Algal Blooms Induced by Recombinant RNAi Microalgae
Xiaodong Deng, Changhao He, Chunmei Xue, Dianlong Xu, Juncai Li, Xiaowen Fei

TL;DR
This study uses genetically modified algae to silence genes in Aedes albopictus mosquitoes, reducing their population and improving water quality.
Contribution
A novel RNAi-based biocide using recombinant microalgae to control Aedes albopictus and alter aquatic ecosystems.
Findings
RNAi recombinant algae caused high mortality and disrupted development in Aedes albopictus larvae.
Recombinant Chlorella reduced mosquito populations and improved water quality in simulated field experiments.
Algal blooms triggered biotic community changes, including proliferation of anaerobic bacteria.
Abstract
This study explores a novel approach to controlling Aedes albopictus, a key vector for dengue fever, by using RNA interference (RNAi) mediated by genetically modified microalgae. We constructed double-stranded RNA (dsRNA) expression vectors that target the cyp314a1 and cyp315a1 genes of Ae. albopictus, and we transformed two species of algae, Chlamydomonas reinhardtii and Chlorella vulgaris, to express these dsRNAs. The recombinant RNAi algal strains were then fed to Ae. albopictus larvae, resulting in high mortality rates, disrupted pupation and reduced emergence. The algae triggered an increase in reactive oxygen species (ROS) and enzyme activity in the larvae, indicating redox stress. In simulated field experiments, RNAi recombinant Chlorella significantly reduced mosquito populations, and improved water quality by reducing nitrogen, phosphorus and others. Furthermore, with the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
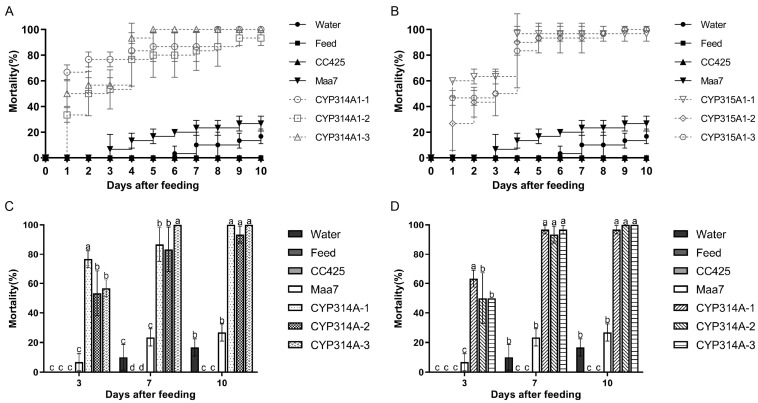 Figure 1
Figure 1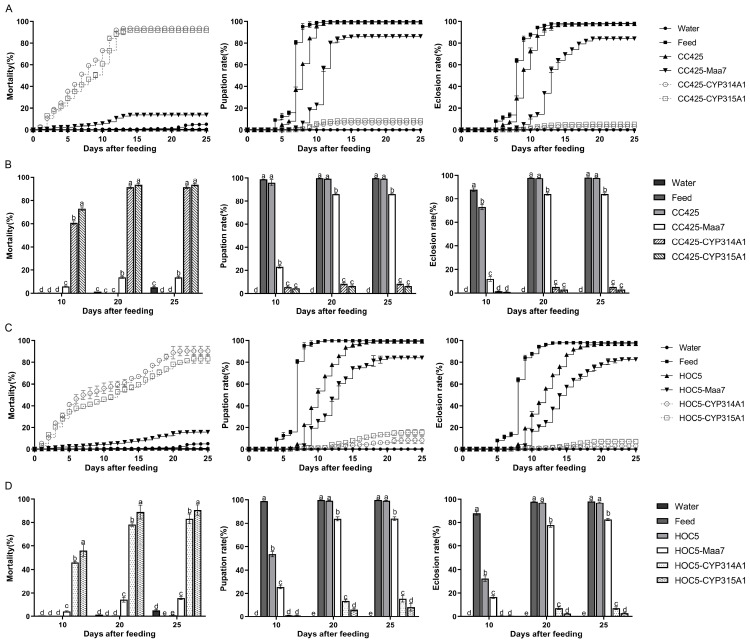 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7 Figure 8
Figure 8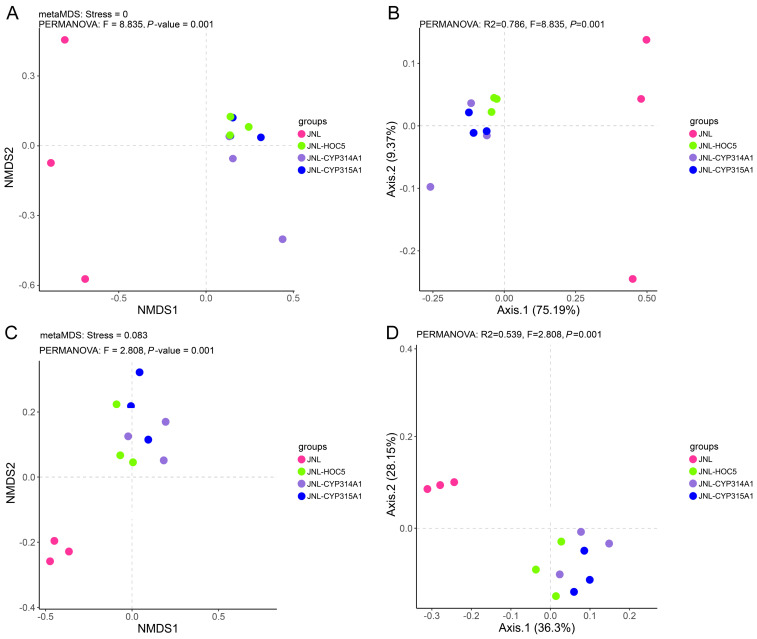 Figure 9
Figure 9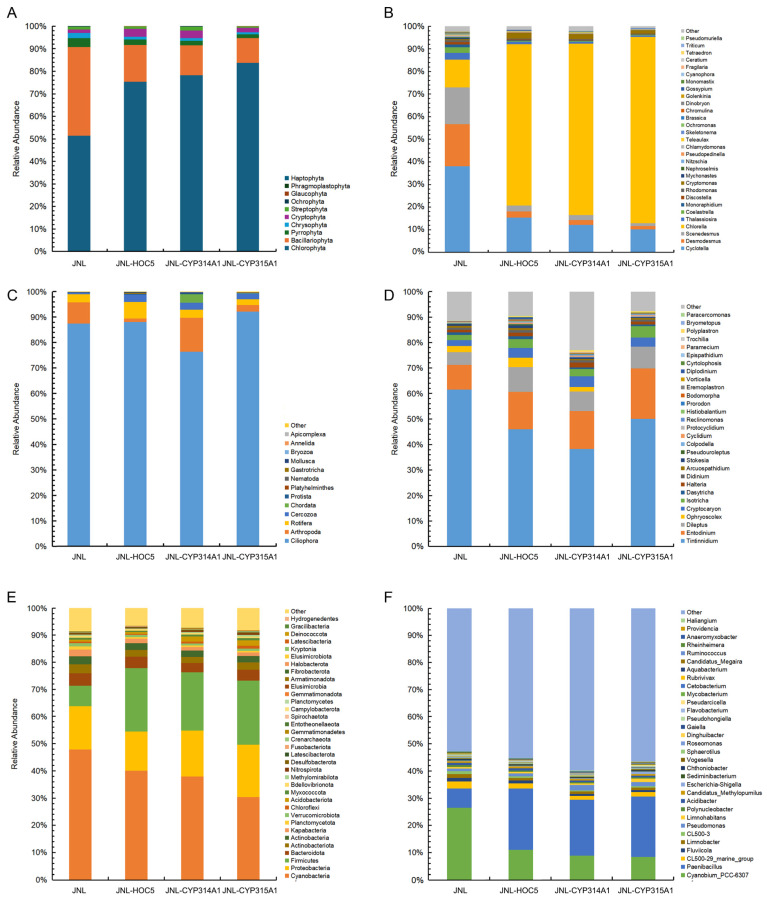 Figure 10
Figure 10 Figure 11
Figure 11- —Key Research and Development Projects of Hainan Province
- —National Natural Science Foundation of China
- —Hainan Medical University Academic Enhancement Support Program Project
- —Hainan Medical University Science and Innovation Program
- —Financial Fund of the Ministry of Agriculture and Rural Affairs, P. R. China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect Resistance and Genetics · Mosquito-borne diseases and control · Entomopathogenic Microorganisms in Pest Control
1. Introduction
Dengue fever is an acute arboviral disease primarily transmitted through Aedes mosquito bites [1]. According to World Health Organization (WHO) surveillance data (2023 update), reveal a dramatic tenfold increase in reported cases, from 500,000 in 2000 to 5.2 million in 2023. This increase has been accompanied by over 5000 dengue-related fatalities, representing the highest mortality figures documented since systematic monitoring began [2,3,4].
The primary vectors of Dengue fever are Aedes mosquitoes, notably Ae. aegypti and Ae. albopictus. These species alsotransmit Zika virus disease, Yellow fever, Rift Valley fever, and Chikungunya fever [5,6,7]. Ae. albopictus, in particular, demonstrates high ecological plasticity and has become the main dengue vector in China. It is distributed across a wide range of climates, from temperate regions (up to 45° north latitude) to tropical ecosystems.
Using biotechnological interventions to suppress vector populations represents an inevitable paradigm shift in the control of mosquito-borne diseases. Current advancements focus on two main strategies: Wolbachia-based population replacement and transgenic mosquito approaches that induce male sterility through genetic modification [8,9,10,11,12]. However, scalability remains challenging due to the substantial costs associated with the mass rearing of mosquitoes and the sex sorting protocols required for field implementation [13,14]. Microalgae-mediated biocontrol is a promising alternative that leverages natural trophic relationships in mosquito larval habitats. Microalgae such as Chlorella spp., Scenedesmus spp., Spirulina spp. and Chlamydomonas spp. are essential dietary components for developing Aedes larvae, and can be cultivated on a large scale at low cost. These microalgae offer distinct operational advantages. Compared to Wolbachia-infected or transgenic male-sterile Ae. aegypti strains, algal-based interventions are more economically feasible and easier to control environmentally. Strategic deployment in contained aquatic ecosystems enables rapid algal dominance, thus gradually reducing the number of Aedes mosquitoes in the area. This novel approach to vector suppression is particularly promising in interrupting the transmission networks of arboviruses such as dengue, Zika and yellow fever.
We have used microalgae expressing RNA interference fragments of the Aedes HR3 (Hormone receptor 3), 3HKT (3-Hydroxykynurenine transaminase) and CHSA (Chitin synthase A) genes to effectively control Aedes populations [15,16,17]. However, it has been shown that prolonged selection pressure for silencing against specific gene targets leads to tolerance (drug resistance) in mosquitoes to the targeted RNAi-interfering algal strains [18,19,20,21]. Cytochrome P450 (CYP450) is an ancient and widely functional superfamily of monooxygenases [22]. The genes encoding these enzymes are among the largest gene superfamilies in plants, animals and fungi and are essential for development, growth, and reproduction [23,24]. CYP450 are present in almost all organisms, including some viruses, with a few exceptions, such as Escherichia coli. Insects possess only four CYP450 clades, which is significantly fewer than the ten found in humans [25,26]. The functions of CYP450 in insects are diverse and include metabolic detoxification, growth and development regulation, and environment adaptation. In metabolic detoxification, insects adapt to the chemical defences of host plants by catalysing the hydroxylation or oxidation of toxic plant xenobiotics to produce less toxic or non-toxic compounds via CYP450 [27,28,29,30,31,32,33,34,35]. CYP450 can also degrade insecticides [36,37]. For instance, RNAi silencing of the MusiDN2722 CYP450 gene in Megalurothrips usitatus significantly increased susceptibility to acetamiprid, confirming the direct role of CYP450 in insecticide detoxification [38]. Additionally, insect CYP450s participate in the biosynthesis of ecdysone to regulate larval moulting, metamorphosis, and embryonic development [39,40,41,42]. These CYP450s include neverland (CYP307A1), spookier (CYP307A2), phantom (CYP306A1), disembodied (CYP302A1), shadow (CYP315A1), and shade (CYP314A1). CYP315A1 and CYP314A1 jointly catalyse the final conversion of cholesterol into the physiologically active 20-hydroxyecdysterone (20E) [43,44]. Given the crucial role of ecdysone in inset development and metamorphosis, this study selected the key enzymes CYP314A1 and CYP315A1 in the 20E anabolic pathway of Aedes mosquitoes as RNAi targets. The dsRNAs of these enzymes were then recombined into the genomes of Chlorella vulgaris and Chlamydomonas reinhardtii, and the microalgae were fed to Aedes mosquito larvae in order to inhibit the growth of Aedes mosquito populations.
This study aims to utilise low-cost microalgae by releasing recombinant strains into waters adjacent to mosquito breeding sites to control local mosquito populations, while concurrently assessing the environmental impact of algal blooms induced by these engineered microalgae. However, due to governmental regulations on genetically modified organisms (GMOs), the direct release of recombinant microalgae into natural water bodies (e.g., ponds and lakes) was prohibited. Consequently, we established a contained experimental area that simulated wild-type aquatic conditions in order to replicate microalgal growth and reproduction dynamics, while also monitoring of mosquito population numbers. From September 2022 to June 2023, we systematically monitored phytoplankton, zooplankton, and prokaryotic communities in the simulated aquatic environment using water quality assays and 18S and 16S rDNA high-throughput sequencing. The population densities and structural profiles of aquatic organisms were analysed under Chlorella bloom conditions. These findings provide foundational data for the future practical application of recombinant microalgae in the control of mosquito populations and the prevention of mosquito-borne diseases such as dengue fever.
2. Methods
2.1. Bioinformatic Analysis
The CYP314A1 and CYP315A1 proteins in Ae. albopictus and other organisms were identified using the Ae. albopictus CYP314A1 and CYP315A1 protein sequence as seed sequences in BLASTP (Version 2.13.0, Bethesda, MA, USA) searches of the Vectorbase (https://vectorbase.org/vectorbase/app, accessed on 7 May 2024) and NCBI (National Center for Biotechnology Information) (https://www.ncbi.nlm.nih.gov/, accessed on 7 May 2024) databases. An online InterProScan analysis (https://www.ebi.ac.uk/interpro/search/sequence/, accessed on 16 April 2025) was carried out to identify the domains of the CYP314A1 and CYP315A1 proteins. A phylogenetic tree was then constructed using MEGA11 software (Version 11, Temple University, Philadelphia, PA, USA) based on the alignment of the CYP314A1 and CYP315A1 orthologue in Ae. albopictus and other organisms. The tree was generated using the neighbour-joining (NJ) method with Poisson correction and pairwise deletion parameters. Based on sequence information obtained from online databases, a schematic representation of CYP314A1 and CYP315A1 homologous protein structures was created. To identify CYP450 homologous proteins, conserved domain sequences of Ae. albopictus CYP450, including WxxxR (Helix-C), GxE/DTT/S (Helix-I), ExxR (Helix-K), PxxFxPE/DRF (PERF), and PFxxGxRxCxG/A (heme-binding domain), were queried in the PFAM (http://pfam.xfam.org/, accessed on 16 April 2025), NCBI and VactorBase (https://vectorbase.org/vectorbase/app, accessed on 15 April 2025) databases After filtering out redundant, 100% identical transcripts from the protein-coding sequence, those encoding at least 300 amino acids were retained. All CYP450 amino acid sequences were collected and annotated using InterPro, and multiple sequences were aligned using ClustalW (Version 11, Dublin, Ireland) and MEGA11. The JTT model was selected for the above processed sequences using maximum likelihood tree. After constructing the tree, iTOL was used to classify it by clan.
2.2. Mosquito Maintenance
Mosquitoes rearing was performed as previously reported [15,16,17]. Local wild type Ae. albopictus were captured in Haikou, China, and propagated for five generations in the laboratory, which were kept in an insectary at the Chinese Academy of Tropical Agricultural Science. The mosquitoes were kept at a temperature of 25 °C and a relative humidity of 80%, with a 12 h cycle of darkness and light. The mosquito cages were also equipped with a sponge saturated with sugar water to provide nourishment. Chicken blood was used to feed the female mosquitoes. Following oviposition on moist filter paper, the eggs were harvested and kept in a dry state. The eggs were then transferred to incubation water containing rat food to facilitate hatching.
2.3. Microalgae Strains and Plasmids
The Chlorella vulgaris HOC5 strain was isolated from local waters in Hainan, China, and cultured in TAP medium. Chlamydomonas reinhardtii CC425 was purchased from the Freshwater Algae Culture Collection at the Institute of Hydrobiology (FACHB), Chinese Academy of Sciences. Liquid cultures were maintained at 25 °C in a shaker at 180 rpm under continuous light (150 µmol/(m^2^·s)) [16]. Plasmid pMD18-T was constructed locally by our research group and stored in our laboratory. The pMaa7 IR/XIR RNAi expression vector was purchased from the Chlamydomonas centre at Duke University in Durham, NC, USA.
2.4. Construction of RNAi Vectors
Total RNA was extracted from Ae. albopictus larvae using TRIzol reagent (Takara, Shiga, Japan) according to the manufacturer’s protocol [15]. Complementary DNA (cDNA) was then synthesised from the total RNA using oligo-dT primers (Shenggong, Shanghai, China) and served as the template for PCR amplification. The cyp314a1 (AALFPA_063667) and cyp315a1 (AALFPA_079547) DNA fragment were PCR-amplified using the primers listed in Supplementary Table S1, respectively. These primers target the corresponding cyp314a1 and cyp315a1 CDS regions from 390 to 805 and 5 to 324, respectively. The amplified fragments were then inserted into the pMD18-T plasmid backbone (pMD-cyp314a1/cyp315a1). These fragments were then digested with HindIII/BamHI and XbaI/SalI, before being inserted into the corresponding cloning sites of the pT282 plasmid, which contained the inverted repeat sequence of the respective genes. The RNAi (pMaa7 IR/XIR) and intermediate (pT282-cyp314a1/cyp315a1) vectors were both digested using EcoRI and then ligated to produce the RNAi recombinant vectors, pMaa7 IR/cyp314a1IR and pMaa7 IR/cyp315a1IR [45].
2.5. Microalgae Transfection
The Chlamydomonas/Chlorella was grown, collected by centrifugation, and resuspended in electroporation buffer. For transfection, 2 µg of either pMaa7 IR/cyp314a1IR or pMaa7 IR/cyp315a1IR was mixed with the 100 µL recipient Chlamydomonas/Chlorella cells (2 × 10^3^ cells µL^−1^) and the mixture was then incubated on ice for 15 min. The mixture was then transferred into an electroporation cuvette and subjected to an electric pulse in an electrometer (BTX, ECM 630, Shelton, WA, USA) with a 1600 V/cm voltage and a pulse time of 1 ms. The suspension was then transferred to TAP medium containing 60 mmol/L sorbitol and allowed to recover overnight. Finally, the cells were harvested by centrifugation at 10,000× g for 3 min and plated on TAP agar plates containing 10 µg/mL paromomycin. The plates were incubated at 23 °C under continuous illumination of 100 µmol m^−2^ s^−1^. Algal colonies appeared after 5–10 days [46,47].
2.6. Mosquito Feeding Experiments
After the RNAi vector, pMaa7IR/cyp314a1IR and pMaa7IR/cyp315a1IR had been transferred into Chlamydomonas/Chlorella. The transgenic microalgae were characterised using PCR to determine if the cyp314a1 or cyp315a1 inverted repeat expression cassette had been integrated into the Chlamydomonas/Chlorella genome. The positive clones were used to feed the mosquitoes in the laboratory. As previously described, ten and 300 mosquito feeding experiments were performed in the insectary [15,16]. In the 10-mosquito feeding experiment, the mosquitoes were divided into experimental and control groups. Each group contained 10 L1 larvae in 5 mL water supplemented with 75 mg fresh microalgae. The larvae in the experimental groups were fed the cyp314a1 and cyp315a1 RNAi transgenic Chlamydomonas/Chlorella strains, respectively. In contrast, the larvae in the control groups were fed C. reinhardtii CC425 or C. vulgaris HOC5, water, feed, and the empty plasmid pMaa7IR/XIR transgenic strain. The experiments were performed in triplicate, recording the mortality, pupation, and adult emergence rates.
In the 300-mosquito feeding experiment, 300 L1 larvae were placed into 50 mL of water supplemented with 750 mg of fresh microalgae. The larvae feeding the recombinant Chlamydomonas/Chlorella were set as the test treatments, whereas the larvae fed with C. reinhardtii CC425 or C. vulgaris HOC5, water, and feed were used as controls. The triplicate experiments included records of mortality, pupation, and adult emergence rates.
To measure body size, five third-instar mosquito larvae were randomly selected from each treatment and control group. Larva was placed a stereomicroscope (Keyence VHX-1000 Super Depth 3D Stereomicroscope, Osaka, Japan), and body length and width were measured using the built-in scale. Body width was determined at the widest segment, and body length was measured from the anterior margin of the head to the tip of the abdomen. The values represented are the mean measurements from the five larvae.
2.7. Measurement of Larval Superoxide Dismutase (SOD), Peoxidase (POD) and Catalase (CAT) Activities
The larvae were fed either C. reinhardtii CC425 or C. vulgaris HOC5 (used as controls). Some larvae were also fed recombinant Chlamydomonas CC425-CYP314A1 and CC425-CYP315A1, while others were fed recombinant Chlorella HOC5-CYP314A1 and HOC5-CYP315A1. Ten to fifteen L4 larvae were sampled from each group, mixed with 5 mmol/L phosphate-buffer solution (pH 7.8, Shenggong, Shanghai, China), ground and homogenised on ice, then centrifuged at 10,000× g for 20 min at 4 °C. The soluble protein content was determined using the G-250 method (Bradford Protein Concentration Assay Kit, Beyotime Biotechnology, Shanghai, China). SOD activity was measured using an SOD activity assay kit (Beyotime Biotechnology, Shanghai, China), POD activity was measured using a peroxidase (POD) activity assay kit (Beijing Solarbio Technology Co., Ltd., Beijing, China), and CAT activity using a catalase assay kit (Beyotime Biotechnology, Shanghai, China) [48,49,50,51].
2.8. The 20-Hydroxyecdysone(20E) Compensation Test
A total of six groups were tested. In each group, 30 Ae. albopictus larvae were fed a 15 mg/mL algal solution. 300 µL of 20E (1 mg/mL) was added to the relevant groups, and the mosquitoes were fed for 15 days [52,53]. Mosquitoes fed C. reinhardtii CC425 and C. reinhardtii CC425+20E were used as control groups. The treatment groups comprised mosquitoes fed cyp314a1 and cyp315a1 RNAi transgenic Chlamydomonas lines and mosquitoes fed cyp314a1 and cyp315a1 RNAi transgenic Chlamydomonas lines with the addition of 20E. The mortality rate, pupation rate and eclosion rate were recorded for each group. Total RNA was extracted from L4 larvae, and the expression levels of the relevant target genes (cyp314a1 and cyp315a1) were detected in the larvae by RT-qPCR. The experiment was repeated three times.
2.9. RT-qPCR
For the reverse transcription quantitative polymerase chain reaction (PCR), 20–30 L3 larvae were collected and pooled from each treatment. Total RNA was isolated from the larvae using TRIzol reagent (Takara, Shiga, Japan), following the manufacturer’s instructions [54]. Single-stranded cDNA was synthesised from the total RNA using oligo-dT primers (Shenggong, Shanghai, China). Real-time PCR was performed using a BioRad iCycler iQ Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA) and SYBR Green as a fluorescent dye. Primers targeting the Aedes RPS17 gene were used as internal controls [55]. The primers used to amplify the cyp314a1 and cyp315a1 target genes, as well as the primers used to amplify the genes involved in the clathrin-mediated endocytosis (CME) pathway, are listed in the Supplementary Table S1. The amplification rate (Ct) of each transcript was calculated using the PCR baseline subtraction method and iCycler software (version 2.3, Bio-Rad, Hercules, CA, USA) (constant fluorescence level). The cycle threshold (Ct) was determined in triplicate and the relative fold differences were calculated using the relative quantification method (2^−ΔΔCT^) [56]. Gene expression was determined relative to the endogenous control.
2.10. Simulated Field Trials
Simulated field trials were performed as described in the previous study [16]. An unused factory building on the outskirts of Haikou City was chosen for the experiment. The mosquitoes were reared in polyester mesh cages with a mesh size of 0.4 mm and a capacity of 1.5 m^3^ [57]. Four 1000 L buckets for microalgae production were placed in the cages, which were fitted with LED lights. An aeration pump was used to ensure constant water circulation. The buckets were then filled with 10 L of cultivated microalgae. From 10 L to 200 L, sufficient light and aeration were maintained for the Chlorella culture. Jinniu Ling reservoir water was then added to bring the volume to 800 L, and the incubation process continued until the concentration of Chlorella cells reached 2 × 10^6^–2 × 10^7^ cells/mL. Around 1000 L1 larvae were placed in each bucket. A 10% sucrose solution was prepared for the male adults, while the egg-laying females were given chicken blood. The number of adult Aedes mosquitoes was counted weekly, in a similar manner to how the abundance of Chlorella in the water was determined. Blood and sugar water were discontinued one day prior to counting the mosquitoes to temporarily reduce their physical strength. Mosquito counting was conducted between 3:00 and 6:00 a.m. A dim LED flashlight was used to illuminate the cages during the procedure [16].
Water quality parameters were analysed in accordance with the Environmental Quality Standards for Surface Water (GB 3838-2002) [58]. Concentrations of ammonium nitrogen (NH_3_-N), nitrate (NO_3_^−^), nitrite (NO_2_^−^) and chemical oxygen demand (COD) were quantified using a Hach DR3900 spectrophotometer (Hach Company, Loveland, CO, USA). Nitrogen (N) levels were determined through alkaline persulfate digestion followed by UV spectrophotometric detection, while phosphorus (P) concentrations were measured using the ammonium molybdate spectrophotometric method [59,60].
2.11. Sample Preparation and DNA Extraction of the Test Water
An 18S and 16S rDNA high-throughput sequencing analysis was performed on the water from simulated-field trials to detect the effect of recombinant RNAi Chlorella on eukaryotic and prokaryotic organisms in the test water. After feeding Ae. albopictus for 30 days, a 1 L water sample was collected from the bucket containing the mosquitoes and filtered through a 0.22 μm polycarbonate membrane (Millipore, Burlington, MA, USA) under a 30 kPa vacuum to collect the plankton. The membranes were stored at −80 °C until analysis. Genomic DNA was extracted using a modified Winnepenninckx et al. method [61].
2.12. Amplification and Sequencing of the Hypervariable Region
The V4 hypervariable region of the 18S rDNA and the V3-V4 region of the 16S rDNA were selected as targets for high-throughput sequencing analysis. The target regions were amplified using the primers listed in Supplementary Table S1 [62,63]. For the 18S rDNA sequencing, sequence libraries were constructed using the NEBNext Ultra DNA Library Prep Kit for Illumina (San Diego, CA, USA) in accordance with the manufacturer’s protocol. The libraries were then sequenced on the Illumina HiSeq2500 platform. For 16S rDNA sequencing, the purified PCR products were sequenced on an Illumina MiSeq (Model MiSeq, Illumina, San Diego, CA, USA) platform. Data optimisation was conducted using the Chen and Jiang approach [64]. Low-quality sequences (i.e., those less than 200 base pairs in length) and sequencing primer adapters located at the 3′ ends of the primers were eliminated. After identifying chimeric sequences, high-quality sequences exhibiting 97% sequence identity were grouped into OTUs. The taxonomic categorisation of the OTUs was determined using USEARCH (version 5.2.236, Robert C. Edgar, San Francisco, CA, USA). Alpha diversity analysis was facilitated by calculating indices such as Shannon, Observed Species and Chao1 using QIIME software (version 1.9.1, University of Colorado, Boulder, CO, USA). Differences in community structure between samples or groups were analysed using principal coordinate analysis (PCoA) plots generated with R software (version 2.15.3, R Foundation for Statistical Computing, Vienna, Austria) [65,66].
2.13. Statistical Analyses
Data analysis was performed using the Statistical Package for the Social Sciences (SPSS, version 26.0, IBM, Chicago, IL, USA) and results are presented as the mean ± standard deviation (S.D.). Duncan’s multiple range test was performed to examine significant differences between means. Values with a p-value of less than 0.05 were considered statistically significant. Asterisks indicate statistical significance in all cases: * p < 0.05; ** p < 0.01. Error bars show the standard deviation [67]. The 95% confidence interval (CI) was calculated using IBM SPSS Statistics 26 (version 26.0, IBM, Chicago, IL, USA). For PERMANOVA F-test, differences in microbial community structure among treatments were assessed using a permutational multivariate analysis of variance based on Bray–Curtis dissimilarity matrices. Grouping assignment (treatments) was used as the explanatory factor. The F-statistic was derived by comparing between-group variance to within-group variance, and significance was determined through 999 permutations of sample labels [67].
3. Results
3.1. CYP314A1 and CYP315A1 Proteins
A BlastP search was performed using Ae. albopictus CYP314A1 and CYP315A1 as source sequences to retrieve a large number of amino acid sequences. Several representative CYP314A1 and CYP315A1 orthologues from different species were selected for further analysis (Supplementary Tables S2 and S3), including characterisation of the structural domains of the nucleic acids and proteins (Supplementary Figures S1 and S2) and phylogenetic analyses. Analysis of the CYP314A1/CYP315A1 protein sequences using the online tool Interproscan (https://www.ebi.ac.uk/interpro/search/sequence/, accessed on 16 April 2025) revealed that all p450 domains were present. Results from protein sequence alignment showed that all p450 domains of CYP314A1 and CYP315A1 contained the following functional regions: WxxxR (Helix-C), GxE/DTT/S (Helix-I), ExxR (Helix-K), PxxFxPE/DRF (PERF), and PFxxGxRxCxG/A (heme-binding domain) (Supplementary Figures S3 and S4). Analysis of the CYP450 gene family in Ae. albopictus revealed that the species contains 81 genes distributed among clan 2 (nine genes), clan 3 (52 genes), and clan 4 (27 genes), as well as the MITO family (seven genes). The genes involved in ecdysone synthesis belonged to clan 2 (e.g., cyp306a1, cyp307a1, and cyp18a1) and the MITO family (e.g., cyp302a1, cyp315a1, and cyp314a1) (Supplementary Figure S5 and Table S4).
3.2. Cyp314a1 and cyp315a1 RNAi Recombinant Chlamydomonas/Chlorella Are Lethal to Ae. albopictus
The target genes selected for RNAi were cyp314a1 and cyp315a1 (AALFPA_063667 and AALFPA_079547), which are key enzymes in the Ae. albopictus 20E synthesis pathway. The cyp314a1 and cyp315a1 RNAi interference fragments were amplified using Ae. albopictus cDNA as a template. The amplified fragments were approximately 416 bp and 319 bp long, respectively. These fragments were then cloned forward and reverse into the pMaa7IR/XIR vector to create the recombinant RNAi vectors pMaa7IR/cyp314a1IR and pMaa7IR/cyp315a1IR. These RNAi vectors were subsequently transformed into C. reinhardtii CC425 and C. vulgaris HOC5. Several positive recombinant algal strains were identified through the implementation of PCR, and these were then employed in subsequent experiments (Supplementary Figure S6).
In the laboratory Ae. albopictus larvae feeding assay, larvae fed with the RNAi recombinant C. reinhardtii CYP314A(1-3) and CYP315A(1-3) began dying on the second day, with mortality rates of 93.3–100% (95% CI: 88.22–100%) and 96.7–100% (95% CI: 94.11–100%) on the 10th day, respectively. Conversely, no mortality was observed in the larvae fed C. reinhardtii CC425 and feed; only 16.7% and 30% of larvae died when fed water and the empty plasmid Maa7IR/XIR transgenic line, respectively. These results suggest that the oral administration of cyp314a1 and cyp315a1 RNAi recombinant C. reinhardtii is lethal to Ae. albopictus larvae (Figure 1).
3.3. Effects of RNAi Recombinant Chlorella on Larval Development, and Redox Stress and 20E Compensation Assay
Approximately 300 L1 Ae. albopictus larvae in each treatment group were subjected to a 25-day feeding experiment. The larvae fed with the recombinant C. reinhardtii CC425-CYP314A1 and CC425-CYP315A1 began to die on the second day and 93.67% (95% CI: 91.19–96.15%) and 91.56% (95% CI: 88.89–94.22%) died within 25 days, respectively. In the control groups, 5.11% (95% CI: 2.72–7.51%), 0.22% (95% CI: 0–0.69%), 0.67% (95% CI: 0.67–0.67%) and 13.78% (95% CI: 11.67–15.85%) of the larvae died when fed water, feed, C. reinhardtii CC425 and transgenic C. reinhardtii pMaa7IR/XIR, respectively (Figure 2A). Similarly, the larvae fed recombinant Chlorella HOC5-CYP314A1 and HOC5-CYP315A1 began dying on the second day, with 90.44% (95% CI: 79.8–100%) and 83.11% (95% CI: 73.09–93.13%) dying within 25 days, respectively. By contrast, in the control groups, only 5.11% (95% CI: 2.72–7.51%), 0.22% (95% CI: 0–0.69%), 0.78% (95% CI: 0–1.74%) and 15.67% (95% CI: 12.35–18.89%) of larvae died when fed water, feed, Chlorella HOC5 and pMaa7IR/XIR transgenic Chlorella, respectively (Figure 2B).
Larvae fed the control diet began pupating on day 4, and all had completed pupation by day 12. Those fed C. reinhardtii CC425 began pupating on day 5. By day 11, 99.33% (95% CI: 99.33–99.33%) of the larvae had pupated. Larvae fed pMaa7IR/XIR transgenic C. reinhardtii CC425 began pupating on day 7, with 86.22% (95% CI: 84.13–88.31%) having pupated by day 16. Larvae fed on recombinant Chlamydomonas CC425-CYP314A1 and CC425-CYP315A1 began to pupate on day 6. By day 14, 6.33% (95% CI: 3.85–8.81%) and 8.44% (95% CI: 5.78–11.11%) of the larvae had pupated, respectively (Figure 2A). Similarly, the larvae fed the recombinant Chlorella HOC5-CYP314A1 and HOC5-CYP315A1 began to pupate on day 7. By day 25, 8.22% (95% CI: 0.19–16.62%) and 15.44% (95% CI: 8.54–22.34%) of the larvae had pupated, respectively. In the control groups, 0%, 99.78% (95% CI: 99.31–100%), 99.22% (95% CI: 98.26–100%) and 84.00% (95% CI: 80.21–87.8%) of the larvae pupated when fed water, feed, Chlorella HOC5 and pMaa7IR/XIR transgenic Chlorella, respectively (Figure 2B).
The emergence rates of adult mosquitoes showed that those fed feed began to emerge on day 5, with 98.11% of pupae emerging within 25 days. Those fed C. reinhardtii CC425 began to emerge as adults on day 6, with 97.78% (95% CI: 97.31–98.25%) of pupae ultimately emerging. However, Aedes pupae emerged as adults at lower rates of 2.89% (95% CI: 0–6.34%) and 5.11% (95% CI: 0.12–10.1%) when fed recombinant Chlamydomonas CC425-CYP314A1 and CC425-CYP315A1 strains, respectively (Figure 2A). Similarly, only 2.89% (95% CI: 1.94–3.84%) and 7.11% (95% CI: 4.21–10.01%) of Aedes pupae emerged as adults when fed the recombinant Chlorella HOC5-CYP314A1 and HOC5-CYP315A1 strains, respectively. In the control groups, adult mosquito emergence was observed to be 0%, 98.11% (95% CI: 96.38–99.84%), 96.89% (95% CI: 95.16–98.62%) and 82.56% (95% CI: 80.16–84.95%) when the larvae were fed water, feed, Chlorella HOC5 or pMaa7IR/XIR transgenic Chlorella, respectively (Figure 2B).
The Ae. albopictus fed with feed had the longest L3 larval body length (5.56 mm/5.89 mm), followed by mosquitoes fed with C. reinhardtii CC425 or C. vulgaris HOC5, with body lengths of 4.63 mm and 4.35 mm, respectively. The body length of mosquitoes fed with water was the shortest (1.84 mm), whereas the body length of the mosquitoes fed with recombinant lines was significantly lower (p < 0.05) than that of the control group (fed with feed, C. reinhardtii CC425 or C. vulgaris HOC5) (Figure 3A–D). In terms of body width, the Ae. albopictus fed with feed had the widest L3 larval body width (0.80 mm), followed by mosquitoes fed with C. reinhardtii CC425 or C. vulgaris HOC5, with body widths of 0.59 mm and 0.57 mm, respectively. The body width of mosquitoes fed with water was the narrowest (0.21 mm), whereas the body width of other mosquitoes fed with recombinant strains was significantly lower (p < 0.05) than that of the control group (fed with feed, C. reinhardtii CC425 or C. vulgaris HOC5) (Figure 3A–D). A notable observation was that Ae. albopictus fed recombinant Chlorella exhibited a reduction in body length and width when compared with Ae. albopictus fed recombinant C. reinhardtii. (Figure 3A–D).
Activity assays of superoxide dismutase (SOD), peroxiredoxin (POD) and catalase (CAT) in Ae. albopictus larvae revealed significantly higher enzyme activity in larvae fed cyp314a1 and cyp315a1 RNAi recombinant Chlamydomonas or Chlorella than in the control group (fed C. reinhardtii CC425, or C. vulgaris) (Figure 4).
Results from the 20E compensation assay revealed that Ae. albopictus fed on recombinant Chlamydomonas supplemented with 20E exhibited a substantial reduction in larval mortality, as well as a notable increase in pupation and emergence rates (Figure 5A). Expression analysis of cyp314a1 and cyp315a1 revealed that mRNA levels of both target genes were significantly reduced (p < 0.05) in Ae. albopictus fed either cyp314a1 or cyp315a1 RNAi recombinant Chlamydomonas, regardless of whether 20E was supplemented (Figure 5B).
3.4. The Expression of the RNAi Target Genes and the Genes in the Clathrin-Mediated Endocytosis (CME) Pathway
Real-time PCR was used to examine cyp314a1 and cyp315a1 gene expression in Ae. albopictus larvae fed with recombinant Chlamydomonas and Chlorella strains. Larvae fed with C. reinhardtii CC425 or C. vulgaris HOC5 were used as controls. The expression levels of the cyp314a1 and cyp315a1 in larvae fed the recombinant Chlamydomonas or Chlorella strains were significantly lower (p < 0.05) than in the control larvae (Figure 6A,B). The most substantial decline in gene expression was observed in larvae fed the cyp314a1 RNAi recombinant Chlamydomonas or Chlorella strains (82.5% and 80.6% decreases, respectively). These observations collectively indicate that the cyp314a1 and cyp315a1 genes in Ae. albopictus were effectively silenced by the recombinant Chlamydomonas or Chlorella strains (Figure 6A,B).
Furthermore, an evaluation was conducted of the levels of several gene transcripts involved in the clathrin-mediated endocytosis (CME) pathway (Supplementary Table S5). These include coat assembly proteins (clathrin adapter AP50 and clathrin heavy chain [Chc]), vesicle formation and budding proteins (liquid facets [lqf], liquid facets-related [lqfR], and dynamin), early endosome proteins (Vha16 and VhaSFD), endosome maturation protein Rab7 and ADP-ribosylation factor-like 1 (Arf72A). Two genes of the flotillin-mediated endocytosis pathway are also included flot-1 and flot-2. The results demonstrated that the mRNA levels of flot-2, Rab7, Chc, Dynamin and vha16 were significantly increased in larvae fed with cyp314a1 shRNA or recombinant Chlorella dsRNA (Figure 6C). Furthermore, the mRNA levels of flot2, Rab7, Chc, lqfR, Dynamin and vha16 were also found to be significantly increased in larvae fed with cyp315a1 shRNA or recombinant Chlorella dsRNA (Figure 6D). These results suggested that knockdown of cyp314a1 or cyp315a1 influences the expression of CME pathway-related genes more than feeding with ds/shRNA-free Chlorella does.
3.5. Simulated-Field Trials
Large-scale simulated field tests were conducted over a period of 16 weeks. The results demonstrated that the Ae. albopictus population fed on Jinniu Ling reservoir water (JNL) increased from 1000 to a maximum of 5063 individuals over 10 weeks. After this, the population slowly decreased to around 4497 individuals after 16 weeks. Ae. albopictus raised in Jinniu Ling reservoir water supplemented with wild-type C. vulgaris HOC5 (JNL-HOC5) increased to a maximum of 6895 individuals after 10 weeks and then decreased slowly to around 6574 individuals after 16 weeks. In the treatment groups where the water was supplemented with recombinant Chlorella HOC5-CYP314A1 or HOC5-CYP315A1, the quantity of Ae. albopictus increased in the first three weeks and then decreased from 1100 (JNL-CYP314A1) or 1090 (JNL-CYP315A1) to zero after 13–15 weeks (Figure 7A–C). These results demonstrate the ability of recombinant Chlorella HOC5-CYP314A1 and HOC5-CYP315A1 to suppress the Ae. albopictus population under simulated field conditions.
To understand the proliferation of RNAi recombinant Chlorella in natural water bodies and its effect on water nutrients, a series of water quality tests and Chlorella density measurements were conducted on each group of water samples. Following a rapid logarithmic growth phase (days 5–7), the recombinant algal strains entered a stabilisation period (days 8–11), after which they entered a period of reduced growth (days 12–90). The removal rates of nitrogen (N), phosphorus (P), nitrate, nitrite, ammonia and chemical oxygen consumption (COD) from the water were found to be positively correlated with the density of Chlorella (Figure 7D). On day 30, the removal rates of N, P, nitrate, nitrite, ammonia, and COD by C. vulgaris HOC5, RNAi recombinant Chlorella HOC5-CYP314A1 and HOC5-CYP315A1 were 86.96–90.51%, 85.39–91.38%, 91.97–93.03%, 80.88–90.54%, 86.79–89.25% and 92.28–93.32%, respectively (Figure 7E). These values corresponded to water samples containing nitrogen, phosphorus, nitrate, nitrite, ammonia, and COD at concentrations of 2.96–3.95 mg/L, 0.07–0.13 mg/L, 2.14–2.44 mg/L, 0.007–0.013 mg/L, 0.03–0.04 mg/L and 31.08–35.56 mg/L, respectively (Figure 7F). By contrast, the removal rates of N, P, nitrate, nitrite, ammonia, and COD in the control Jinniu Ling Reservoir water samples were only 3.9%, 8.89%, 0.04%, 2.41%, 11.66% and 2.73%, respectively. These levels corresponded to concentrations of 37.71 mg/L, 0.81 mg/L, 32.96 mg/L, 0.08 mg/L, 0.31 mg/L and 496.65 mg/L, respectively. By adding RNAi recombinant Chlorella, the levels of N, P, nitrate, nitrite, ammonia and COD in the severely eutrophic Jinniu Ling Reservoir water were reduced to Class V levels (N ≤ 2.0, P ≤ 0.4, nitrate ≤ 10, nitrite ≤ 1, ammonia ≤ 2.0, and COD ≤ 40), in accordance with China’s environmental quality standard for surface water (GB 3838-2002) (Figure 7E,F) [58].
3.6. Plankton Abundance and Diversity in Simulated Field Test Waters
To evaluate the impact of RNAi recombinant Chlorella on aquatic biological communities, high-throughput analyses of 18S rDNA and 16S rDNA was performed on water samples containing the recombinant Chlorella strains HOC5-314A1 and HOC5-315A1. For the 18S rDNA sequencing, 988,356 qualified tags were obtained from the samples after several quality controls steps. After removing unclassified and unique tags from the dataset, the total number of high-quality tags was 818,788. These sequences were then clustered into OTUs with 97% concordance and a total of 1009 OTUs were obtained. Similarly, 1,125,735 qualified tags were obtained from the 16S rDNA sequencing after several quality controls. The total number of high-quality tags was 761,131 and 1213 OTUs were obtained.
To assess and compare planktonic biodiversity across samples, we computed richness and diversity indices, including Chao1, observed species and Shannon indices, for phytoplankton, zooplankton, and prokaryote communities. These indices were based on OTUs clustered at 97% sequence similarity, followed by α-diversity analysis (Figure 8). Compared with the control group (JNL group), the phytoplankton Chao1 index and observed species index in the JNL-CYP314A1 treatment group (supplemented with RNAi recombinant Chlorella strain HOC5-CYP314A) showed no significant differences. The zooplankton Chao1 index and observed species index exhibited an upward trend but did not reach statistical significance. In contrast, the zooplankton Chao1 index and observed species index in the JNL-CYP315A1 treatment group (supplemented with recombinant strain HOC5-CYP315A1) decreased significantly (p < 0.05) compared with the JNL group (Figure 8B). With respect to prokaryotic communities, the JNL-CYP315A1 group displayed a significant increase (p < 0.05) in the observed species index relative to the JNL control group (Figure 8C).
Shannon index analysis revealed that, compared with the JNL group, the phytoplankton community diversity in the JNL-CYP314A1 treatment group decreased significantly (p < 0.05), whereas zooplankton diversity increased markedly (Figure 8A,B). In addition, both phytoplankton and prokaryotic diversity in the JNL-CYP315A1 group declined significantly (p < 0.05) compared with the JNL group (Figure 8A,C). These findings indicate that the supplementation of recombinant Chlorella altered the patterns of aquatic biodiversity in the Jinniu Ling Reservoir.
The NMDS and PCA analyses of the eukaryotic communities revealed that JNL (pink points) was clearly separated from the other groups, indicating that the baseline water community structure differed substantially from that of the treatments with added Chlorella. JNL-HOC5 (green points) clustered closely with JNL-CYP314A1 (purple points) and JNL-CYP315A1 (blue points), suggesting that the community structure became more similar after the addition of Chlorella. However, differences still remained between the RNAi-transgenic Chlorella treatments (JNL-CYP314A1, JNL-CYP315A1) and the wild-type HOC5. The PCA results showed that the first principal component (Axis1) explained 75.19% of the variance, indicating that the major community differences were determined by the addition of Chlorella and its genotype (Figure 9A,B).
The NMDS and PCA analyses of the prokaryotic communities demonstrated that JNL was distinctly separated from the other groups, indicating that the addition of Chlorella had a pronounced effect on the prokaryotic community. JNL-HOC5 overlapped to some extent with JNL-CYP314A1 and JNL-CYP315A1, but the RNAi-treated groups (JNL-CYP314A1, JNL-CYP315A1) displayed a more dispersed distribution, suggesting that the effects of recombinant Chlorella on prokaryotic communities were more complex and variable. The PCA results showed that the Axis1 explained 36.3% of the variance and the Axis2 explained 28.15%, indicating that multiple factors contributed to the differences in the prokaryotic community rather than a single dominant factor (Figure 9C,D).
3.7. Changes in Plankton in Simulated Field Test Waters
A total of ten groups of eukaryotic microalgae were identified at the phylum level in the Jinniu Ling Reservoir (JNL, control) sample, including Chlorophyta, Chrysophyta, Cryptophyta, Bacillariophyta, Pyrrophyta, Streptophyta, Ochrophyta, Phragmoplastophyta, Glaucophyta and Haptophyta (Supplementary Table S6). Of these, 366 OTUs within 83 genera were identified in JNL. Chlorophyta exhibited the highest number of OTUs. A total of 120 OTUs were identified within 28 genera of Chlorophyta. The second largest class was Chrysophyta, comprising 72 OTUs within 15 genera. Cryptophyta, Bacillariophyta and Pyrrophyta also exhibited a relatively high number of OTUs. By contrast, Phragmoplastophyta, Glaucophyta and Haptophyta exhibited lower numbers of OTUs (Supplementary Table S6). In the Jinniu Ling Reservoir water plus Chlorella HOC5 group (JNL-HOC5), 356 OTUs belonging to 86 genera were observed. Chlorophyta exhibited the highest number of OTUs, with 126 across 27 genera. As shown in Supplementary Table S6, Chrysophyta was the second largest phylum, comprising 63 OTUs across 17 genera. In the JNL-CYP314A1 treatment group, 383 OTUs belongs to 92 genera were observed. This was a greater number than in the JNL or JNL-HOC5 control groups. Of these, the number of Cryptophyta, Bacillariophyta and Streptophyta species increased. However, the JNL-CYP315A1 treatment group exhibited a mere 334 OTUs across 80 genera, falling short of the OTUs and genera observed in the control groups (JNL, or JNL-HOC5). Regarding species abundance, Chlorophyta, Bacillariophyta, Pyrrophyta, Chrysophyta and Cryptophyta were found to be more prevalent in the treatment groups (JNL-CYP314A1 and JNL-CYP315A1) and in the control group (JNL, JNL-HOC5). Compared with the control group, the relative abundance of Bacillariophyta, Pyrrophyta and Chrysophyta decreased in the treatment group, while the relative abundance of Chlorophyta increased (Figure 10A).
Analysis at the Phytoplankton genus level showed that adding recombinant Chlorella increased its abundance in the JNL-CYP314A1 and JNL-CYP315A1 treatment groups to 75.80% and 82.41%, respectively, compared to the control JNL group at 12.25%. The abundance of most phytoplankton was decreased in the JNL-CYP314A1 and JNL-CYP315A1 treatment groups, except for Chlorella, Cryptomonas and Teleaulax, whose abundance increased (Figure 10B and Figure 11A). For example, the abundance of Aulacoseira, Cyclotella, Fragilaria, Skeletonema, Thalassiosira, Discostella and Nitzschia in the Bacillariophyta decreased. The abundance of Mychonastes, Tetraedron, Coelastrella, Monoraphidium, Desmodesmus, Scenedesmus and Golenkinia in the Chlorophyta also decreased. Rhodomonas in the Cryptophyta, Ceratium in the Dinophyta, and Dinobryon, Ochromonas and Chromulina in the Chrysophyta, also decreased (Figure 10B and Figure 11A).
Analysis of zooplankton at the phylum level revealed that Ciliophora, Cercozoa, Arthropoda and Rotifera occupied the top positions in terms of species number and abundance (Supplementary Table S7 and Figure 10C). Specifically, Ciliophora exhibited abundances of 87.40%, 88.01%, 76.35% and 92.03% in JNL, JNL-HOC5, JNL-CYP314A1 and JNL-CYP315A1 water, respectively. Analysis of the normalised zooplankton genus-level data showed that the genera Tintinnidium, Entodinium and Dileptus occupied the top three positions in terms of species abundance. Tintinnidium was the most prevalent species in JNL, JNL-HOC5, JNL-CYP314A1 and JNL-CYP315A1 water, with abundances of 61.56%, 45.87%, 38.28% and 49.99%, respectively (Figure 10C).
Analysis of differences in species abundance revealed higher abundances of Cilicophora (Histriculus, Cyclidium, Polyplastron, Histiobalantium and Vorticella) and Protozoa (Heteromita, Reclinomonas and Bodomorpha) in treatment groups JNL-CYP314A1 and JNL-CYP315A1 than in the control group, JNL. In contrast, the Cilicophora (Pseudouroleptus, Trochilia, Bryometopus, Cryptocaryon, Halteria, Didinium, Dileptus, Paracercomonas and Epispathidium), and rumen-specialised groups of ruminants (Entodinium, Ophryoscolex and Dasytricha), as well as other protists such as Colpodella, Cytolphosis, Arcuospathidium and Tintinnidium (which consume diatoms and dinoflagellates), were reduced in abundance (Figure 10C,D and Figure 11B).
3.8. Changes in Prokaryotes in Simulated Field Test Waters
Analysis of the water samples using 16S rDNA high-throughput sequencing revealed the presence of high concentrations of Cyanobacteria, Proteobacteria, Firmicutes, Bacteroidota, Actinobacteriota and Actinobacteria in the control (JNL and JNL-HOC5) and treatment (JNL-CYP314A1 and JNL-CYP315A1) groups (Figure 10E). Cyanobacteria were the most prevalent of these, with proportions of 47.57%, 39.96%, 37.78% and 30.10% in JNL, JNL-HOC5, JNL-CYP314A1 and JNL-CYP315A1, respectively. At the genus level, the top three positions in terms of abundance were occupied by Cyanobium_PCC-6307, Paenibacillus, and CL500-29_marine_group. As shown in Figure 10F, Cyanobium_PCC-6307 was the most prevalent species in the JNL, JNL-HOC5, JNL-CYP314A1 and JNL-CYP315A1 water samples, with concentrations of 25.57%, 10.68%, 8.65% and 8.19%, respectively.
Cyanobium_PCC-6307 belongs to the family Cyanobiaceae, a group of photosynthetic, autotrophic prokaryotes that are widely distributed in freshwater, marine and wetland environments. It has a thylakoid structure typical of cyanobacteria and fixes carbon through photosynthesis, while also being involved in nitrogen metabolism. It is one of the dominant bacterial genera in natural or artificial waters and is an indicator species of eutrophication [68,69,70,71]. The abundance of Cyanobium_PCC-6307 in the JNL-CYP314A1 and JNL-CYP315A1 waters decreased as the N and P contents decreased. Paenibacillus is a genus of Gram-positive bacteria widely distributed in the natural environment. It belongs to the phylum Firmicutes, the class Bacilli, and the family Paenibacillaceae. The genus currently comprises over 150 species, around 20 of which have the ability to fix nitrogen, secrete organic acids to dissolve insoluble phosphorus in water, degrade organic pollutants and reduce the eutrophication of water [72,73,74,75]. The results showed that the Paenibacillus abundance in treated JNL-CYP314A1 and JNL-CYP315A1 water increased significantly (19.92% and 21.54%, respectively) compared to the control (JNL, 6.79%) (Figure 10F). The CL500-29_marine_group belongs to the phylum Actinobacteriota, which contains the dominant bacteria in freshwater and offshore environments. The CL500-29_marine_group is an important phylum of Actinobacteria with significant ecological functions. It can efficiently utilise a variety of carbon sources, such as amino acids, carboxylic acids and lipids. Its abundance is positively correlated with lake carbon concentration and nitrogen and phosphorus removal efficiency, suggesting that it is closely related to water purification capacity [76,77,78,79,80]. The sequencing results obtained in this study align with the following observations: the introduction of recombinant Chlorella into the treatment group let to lower concentrations of carbon, nitrogen and phosphorus in the water, as well as a decrease in the abundance of the CL500-29_marine_group.
Heat map-based analysis of differences in the abundance of prokaryotic species at the phylum level after normalisation revealed that the abundances of Gemmatimonadota, Acidobacteriota, Elusimicrobiota, Armatimonadota, Crenarchaeota, Entotheonellaeota, Kryptonia, Chloroflexi, Methylomirabilota, Latescibacteria, Nitrospirota, Myxococcota and Latescibacterota were higher in the treatment groups (JNL-CYP314A1 and JNL-CYP315A1) than in the control group (JNL). Conversely, the abundances of Cyanobacteria, Actinobacteria, Halobacterota, Kapabacteria, Fusobacteriota, Gemmatimonadetes, Actinobacteriota, Bdellovibrionota, Fibrobacterota, Planctomycetota, Verrucomicrobiota, and Deinococcota were markedly lower in the treatment groups than in the control group (Figure 10E and Figure 11C). At the genus level, the analysis revealed that the abundances of Vogesella, Limnohabitans, Pseudarcicella, Aquabacterium, Rheinheimera, Escherichia-Shigella, Providencia, Gaiella, Haliangium, Pseudomonas, Candidatus, Methylopumilus, and Paenibacillus were higher in the treatment groups (JNL-CYP314A1 and JNL-CYP315A1) than in the control group (JNL). By contrast, the abundance of the following bacteria decreased in the treatment groups compared to the control group: Mycobacterium, Flavobacterium, Anaeromyxobacter, Sphaerotilus, Fluviicola, Cetobacterium, Cyanobium_PCC-6307, Polynucleobacter, Pseudohongiella, Sediminibacterium, Acidibacter, Dinghuibacter, Roseomonas, Rubrivivax. CL500-29_marine_group, Limnobacter and Chthoniobacter were lower in the treatment groups than in the control group (Figure 10F and Figure 11D).
4. Discussion
Ae. albopictus is recognised as a vector of significant public health vector, primarily transmitting viruses such as the Dengue, Yellow fever, Chikungunya and Zika viruses. These diseases cause substantial human mortality each year [81]. In the absence of effective therapies or vaccines, control the vectors remains the most effective strategy for preventing the spread of these pathogens. Extensive research has demonstrated that RNAi-based biopesticides represent a new generation of pesticides and are a powerful tool for controlling insect populations. Compared to other strategies, RNAi biopesticides are more environmentally friendly. However, they also face challenges related to target-site resistance, as evidenced by the emergence of resistance issues following the widespread adoption of transgenic insect-resistant cotton [82,83,84,85]. While the high efficiency and specificity of RNAi biopesticides have garnered significant attention, the environmental impacts of their large-scale deployment and application have not yet been comprehensively evaluated [86,87].
In this study, the cyp314a1 and cyp315a1 genes of Ae. albopictus were selected as RNAi targets. CYP450 is an ancient and multifunctional superfamily of monooxygenases [22]. Insects possess four CYP450 clans with roles in metabolic detoxification, growth regulation and development [25,26]. CYP450 enzymes are involved in ecdysone biosynthesis. Specifically, shadow (sad, CYP315A1) and shade (shd, CYP314A1) catalyse the final steps of cholesterol conversion into the biologically active 20-hydroxyecdysone (20E) [88,89,90]. As they are critical to ecdysone synthesis, the cyp314a1 and cyp315a1 genes are predominantly found in arthropods, suggesting they have minimal impact on humans, higher animals and plants. Our analysis of Ae. albopictus CYP450 genes revealed that the species harbours 81 such genes, distributed across Clan 2 (9 genes), Clan 3 (52 genes), Clan 4 (27 genes) and MITO (7 genes). Genes involved in ecdysone synthesis belong to Clan 2 (e.g., cyp306a1, cyp307a1 and cyp18a1) and the MITO family (e.g., cyp302a1, cyp314a1 and cyp315a1) (Supplementary Figure S5).
Observations demonstrated that Ae. albopictus larvae fed with cyp314a1 or cyp315a1 recombinant RNAi Chlamydomonas exhibited mortality rates of 93.3–100% or 96.7–100%, respectively, within a 10-day period (Figure 1). In a feeding trial involving 300 mosquitoes, larvae fed cyp314a1 or cyp315a1 recombinant RNAi Chlamydomonas showed mortality rates of 93.67% and 91.56% over 25 days, respectively. Those fed cyp314a1 or cyp315a1 recombinant RNAi Chlorella exhibited mortality rates of 90.44% and 83.11%, respectively (Figure 2). These results confirm that the oral administration of cyp314a1 or cyp315a1 RNAi Chlamydomonas/Chlorella has a lethal effect on Ae. albopictus larvae. Pupation rates were 6.33% and 8.44% for larvae fed with cyp314a1 or cyp315a1 RNAi recombinant Chlamydomonas, respectively, and 8.22% and 15.44% for larvae fed with cyp314a1 or cyp315a1 RNAi recombinant Chlorella, respectively (Figure 2). With regard to adult emergence, the emergence rates of larvae fed with cyp314a1 or cyp315a1 RNAi recombinant Chlamydomonas were 2.89% and 5.11%, respectively, while the emergence rates of those fed with cyp314a1 or cyp315a1 RNAi recombinant Chlorella were 2.89% and 7.11%, respectively (Figure 2). These findings suggest that recombinant algal strains expressing Cyp314a1 or Cyp315a1 RNAi significantly disrupt the pupation and adult emergence processes of Ae. albopictus.
Using Ae. albopictus larvae as a model, we observed that feeding them cyp314a1 or cyp315a1 RNAi recombinant algal strains significantly increased the enzymatic activities of SOD, POD and CAT compared to control groups (fed C. reinhardtii CC425 or C. vulgaris HOC5), indicating an enhanced redox stress (Figure 4). The 20E rescue assay revealed that supplementing cyp314a1 or cyp315a1 RNAi recombinant Chlamydomonas with exogenous 20E substantially reduced larval mortality and restored pupation and adult emergence rates (Figure 5A). 20E regulates processes such as moulting, metamorphosis, and tissue apoptosis, and is the key hormone required for maintaining the transition from larva to adult. Deficiency or reduction of 20E leads to developmental arrest and moulting failure in larvae. Redox stress induces cellular damage, mitochondrial disfunction, and energy metabolism disorders. The combined effects of these two factors ultimately result in larval death.
Studies have demonstrated that the dsRNA and shRNA of cyp314a1 or cyp315a1 likely penetrate the interior of mosquitoes via the clathrin-mediated endocytosis (CME) pathway in intestinal epithelial cells. In contrast, invertebrates such as nematodes use the SID pathway to transport dsRNA into and between cells [91]. However, insects primarily absorb exogenous dsRNA through CME [92,93]. Following the knockout of the cyp314a1 or cyp315a1 gene, the mRNA levels of certain key CME pathway genes increased (see Figure 6C,D), corroborating the findings of Abbasi et al. (2020) regarding Ae. aegypti [94].
Subsequent simulated-field trials revealed that RNAi-recombinant Chlorella reduced the Ae. albopictus population from 1000 to zero within 16 weeks (see Figure 7A–C). It also reduced the levels of nitrogen, phosphorus, ammonia, nitrates, nitrites and COD in the experimental water (Jinniu Ling Reservoir), upgrading it to China’s Class V surface water standard (GB 3838-2002) (see Figure 7E,F) [58].
To understand the effect of RNAi recombinant Chlorella on the test water’s biological populations, 18S and 16S rDNA high-throughput sequencing analyses were performed. The results showed that the rapid reproduction of the recombinant Chlorella caused a bloom that consumed nutrients such as C, N and P at a rapid rate. Competition for nutrient salts directly inhibits the growth of other algae, such as diatoms (e.g., Aulacoseira, Cyclotella and Thalassiosira) and green algae (e.g., Scenedesmus and Coelastrella) [95,96,97]. Competition for light resources results in decreased light transmittance (increased turbidity), leading to the decline of photosynthesis-dependent non-planktonic algae (e.g., the attached diatom Fragilaria and the green algae Desmodesmus, Tetraedron and Monoraphidium) due to insufficient light [68]. However, motile species such as Cryptomonas and Teleaulax can acquire light energy through vertical migration, giving them a competitive advantage (see Figure 10A,B and Figure 11A) [98].
During the algal bloom triggered by recombinant RNAi in Chlorella, the water body’s dissolved oxygen (DO) levels exhibited significant diurnal fluctuations: photosynthesis resulted in DO supersaturation during the day, whereas respiration caused hypoxic stress at night. These extreme redox oscillations periodically stressed aerobic zooplankton (Didinium and Halteria) physiologically, driving the dynamic remodelling of the microbial community structure. The drastic decrease in DO at night significantly inhibited the proliferation of the aerobic groups Cyanobium PCC-6307, Limnobacter, Chthoniobacter and Actinobacteria. Meanwhile, the abundance of facultative and specialised anaerobic microorganisms such as Methylomirabilota and Nitrospirota increased. The biomass of the strict anaerobe Chloroflexi and Elusimicrobiota increased significantly compared to the control due to the enhanced anaerobic microenvironment in the bottom sediment (Figure 11C) [99,100,101]. DOM and POM, produced by the decomposition of Chlorella residues during the decline of the algal bloom, provided preferential carbon sources for heterotrophic flora, including Pseudarcicella, Aquabacterium, Vogesella, Rheinheimera, Pseudomonas, Paenibacillus and Haliangium, as well as the Myxococcota. The release of nitrogen and phosphorus derived from the algae, activates the metabolic activities of the nitrifying bacterium Crenarchaeota, the denitrifying bacterium Nitrospirota and the sulfur-oxidising bacterium Gemmatimonadota. Conversely, Cyanobacteria, Planctomycetes and Halobacteria decreased in abundance due to inferior competition for carbon and nitrogen (Figure 10E,F and Figure 11C,D) [102,103,104,105,106].
Analysis based on alpha diversity data showed that adding the wild type of C. vulgaris HOC5 slightly altered the biotope but did not significantly reduce diversity (similar Chao1 and Shannon indices) or impact community stability overall. However, the alpha diversity (e.g., observed species and Shannon index) of the JNL-CYP315A treatment group differed significantly from that of the control JNL group. This indicates that the RNAi recombinant algal strain significantly altered the structure of the biological community, resulting in fewer zooplankton species and decreased diversity (see Figure 8). The significant difference in observed species and Shannon index between the JNL-CYP314A1 and JNL-CYP315A1 treatment groups suggests that the difference in biological impact on the surrounding environment may be due to variations between the CYP314A1 and CYP315A1 RNAi recombinant algal strains. Analysis of the dsRNA target fragments revealed multiple differential fragments between the two strains. BLAST comparison of fragments belonging to the CYP315A1 dsRNA target region in the NCBI database showed that multiple fragments were homologous (≥80%) with some of the less abundant zooplankton species. This may explain the reduction in zooplankton species in the JNL-CYP315A treatment group (Figure 8B and Supplementary Table S8).
In conclusion, changes in environmental water bodies are mainly due to the release of RNAi recombinant Chlorella and community reconfiguration due to resource competition (e.g., nitrogen–phosphorus competition between Chlorella and diatoms), which is induced by algal blooms. Other factors include anaerobic bacterial proliferation and aerobic bacterial decline caused by anaerobic conditions.
While this study has made some progress, the impact of exogenous DNA fragments (particularly antibiotic resistance genes) carried by RNAi recombinant Chlorella on environmental organisms (including those in the direct and indirect food chains) requires further evaluation. Lake algal blooms generally go through a cycle of ‘recovery-emergence-decay-dormancy’ [107]. During the bloom stage, the productivity of phytoplankton increases significantly [108], whereas during the decay and decomposition stage, the apoptosis of algal cells releases large quantities of sugars, proteins, and lignin into the water. These substances then become an important source of DOM in the water [109]. However, this study only examined the biotope composition and structure during the RNAi recombinant Chlorella bloom, and did not consider what happened after the bloom ended. Future studies will explore this to clarify the environmental impact of RNAi recombinant Chlorella algal blooms. Current evidence suggests that the environmental impact of such blooms is limited and similar to that of green algal blooms in urban eutrophic lakes. As the bloom enters the decay and dormancy stage, the biological populations in the water are restructured as nutrients are depleted, which increases species diversity and restores the population balance.
5. Conclusions
In this study, we selected the cyp314a1 and cyp315a1, two genes involved in the ecdysone biosynthesis pathway of Ae. albopictus, as targets for RNA interference (RNAi). The recombinant Chlamydomonas/Chlorella mosquito biocide obtained through genetic transformation exhibited high lethality towards Ae. albopictus larvae. Simulated field trials of the recombinant Chlorella biocide effectively controlled the Ae. albopictus population. The recombinant Chlorella induced a bloom, altering the population structure of the water and inhibiting the growth of most phytoplankton through trophic competition. The anaerobic environment created by the bloom subsequently triggered the proliferation of anaerobic bacteria and the decline of aerobic bacteria. This study is an important addition to research on the use of recombinant microalgae as mosquito biocides.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Waggoner J.J. Gresh L. Vargas M.J. Viremia and Clinical Presentation in Nicaraguan Patients Infected with Zika Virus, Chikungunya Virus, and Dengue Virus Clin. Infect. Dis.2016631584159010.1093/cid/ciw 58927578819 PMC 5146717 · doi ↗ · pubmed ↗
- 2Bhatt S. The global distribution and burden of dengue Nature 201349650450710.1038/nature 1206023563266 PMC 3651993 · doi ↗ · pubmed ↗
- 3WHO Ten Threats to Global Health in 2019 WHO Geneva, Switzerland 2019
- 4Xiang B. Gao P. Kang Y.F. Importation of Zika virus in China: A significant risk in southern China J. Infect.20177432833010.1016/j.jinf.2017.01.00428109676 · doi ↗ · pubmed ↗
- 5Chen J. Lu H.Z. Yellow fever in China is still an imported disease Biosci. Trends 20161015816210.5582/bst.2016.0105127052094 · doi ↗ · pubmed ↗
- 6Liu W. Sun F.J. Tong Y.G. Rift Valley fever virus imported into China from Angola Lancet Infect. Dis.201616122610.1016/S 1473-3099(16)30401-727788984 · doi ↗ · pubmed ↗
- 7Wu D. Wu J. Zhang Q.L. Chikungunya outbreak in Guangdong province, China, 2010 Emerg. Infect. Dis.20121849349510.3201/eid 1803.11003422377135 PMC 3309566 · doi ↗ · pubmed ↗
- 8Hemingway J. Averting a malaria disaster: Will insecticide resistance derail malaria control?Lancet 20163871785178810.1016/S 0140-6736(15)00417-126880124 PMC 6215693 · doi ↗ · pubmed ↗