Dna2 Responds to Endogenous and Exogenous Replication Stress in Drosophila melanogaster
Ivan Rivera, Sabah Shammari, Hamiya Sohail, Christian Villegas, Zoha Wasim, Sze Hang Ip, Vada Becker, Kathryn P. Kohl, Eric P. Stoffregen, Christina I. Swanson, Elyse Bolterstein

TL;DR
This study shows that DNA2 in fruit flies is essential for handling DNA replication stress during development and responding to DNA-damaging agents.
Contribution
The study reveals domain-specific roles of DNA2 in replication stress response and developmental genomic stability in Drosophila.
Findings
Dna2 mutants show increased sensitivity to replication stress and DNA-damaging agents like MMS and topotecan.
Dna2 mutants have reduced fertility and increased DNA damage in germline cells.
Adult lifespan is unaffected in Dna2 mutants, suggesting compensatory mechanisms.
Abstract
Background/Objectives: DNA2 is a conserved nuclease–helicase that plays a crucial role in DNA replication and repair by responding to replication stress. Previous studies have established the role of DNA2 in Okazaki fragment processing, the recovery of stalled replication forks, and double-strand break repair. This study investigates the role of Drosophila melanogaster Dna2 in response to exogenous DNA damage and replication stress as well as during developmental stages involving intensive DNA replication. Methods: We used the Drosophila mutant alleles, Dna2D1 and Dna2D2, which differ in the presence of the helicase 1A domain, to assess sensitivity to mutagens that cause various types of replication stress and DNA damage. We examined reproductive fitness through Mendelian ratio calculations, fecundity, egg viability assays, and assessed DNA damage via immunostaining of ovarian germaria.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
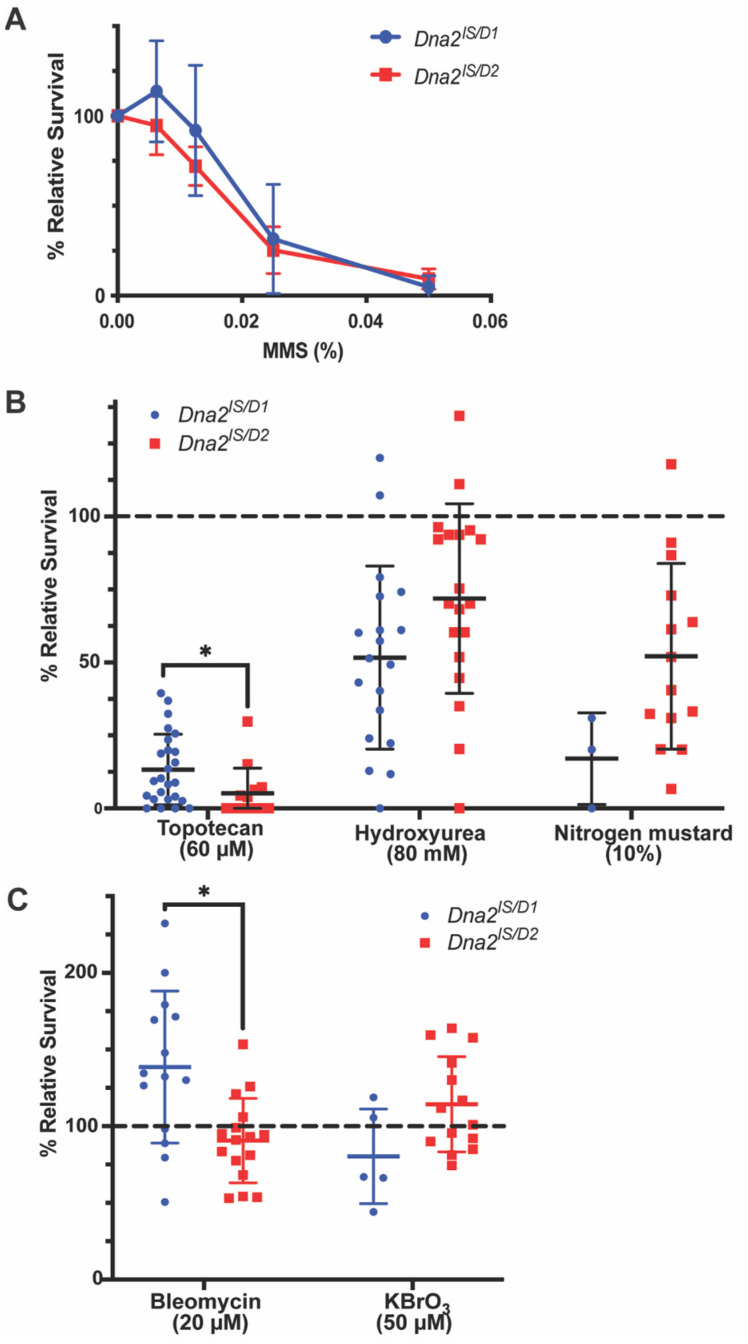 Figure 1
Figure 1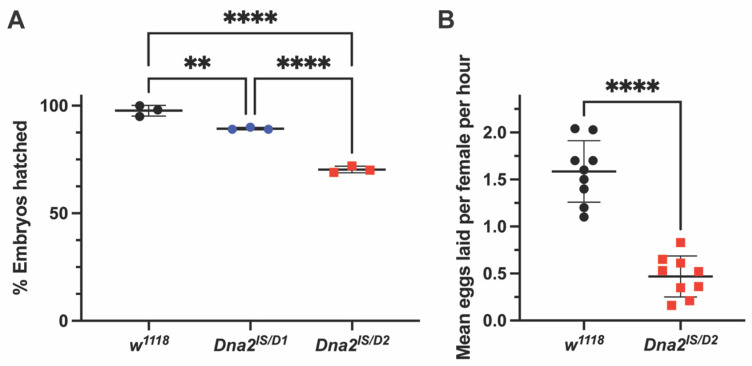 Figure 2
Figure 2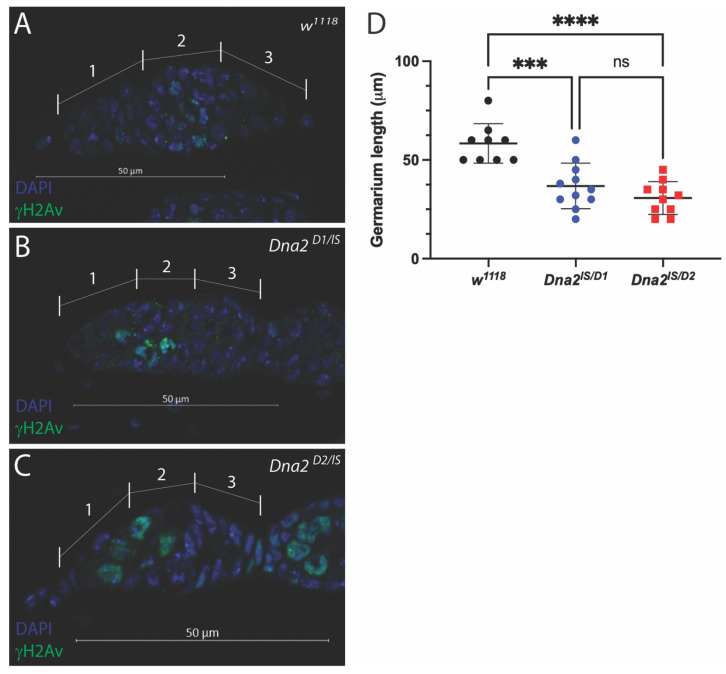 Figure 3
Figure 3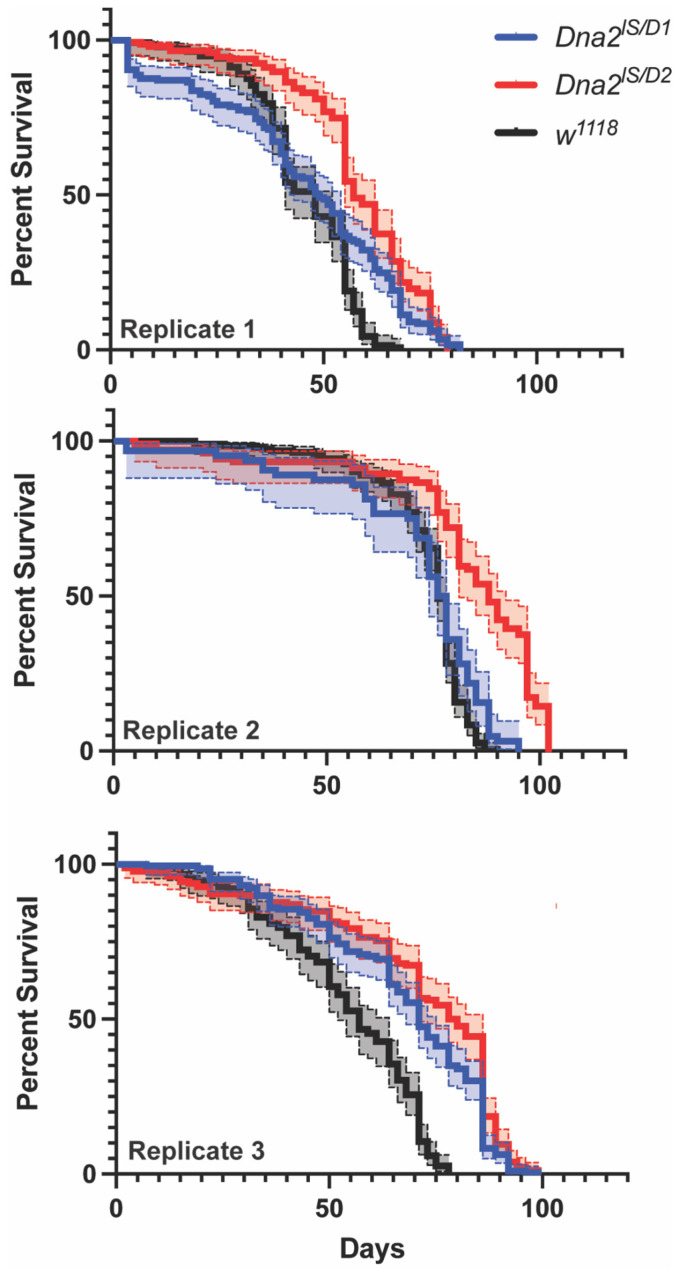 Figure 4
Figure 4- —National Institute of General Medical Sciences of the National Institutes of Health
- —National Science Foundation IUSE
- —Northeastern Illinois University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDNA Repair Mechanisms · Plant Genetic and Mutation Studies · Mitochondrial Function and Pathology
1. Introduction
Cells are continuously challenged by a variety of DNA damage arising from both endogenous sources, such as reactive oxygen species and replication errors, and exogenous sources like UV light and chemical mutagens. To maintain genomic integrity and ensure accurate transmission of genetic information, cells rely on DNA repair proteins that detect, process, and resolve DNA damage [1]. Failures in repair pathways are linked to a wide range of diseases, including cancer, neurodegeneration, and developmental disorders [2]. Because of their potential in treating diseases, it is important to develop models in which individual DNA repair proteins are characterized for further research.
The DNA2 protein is an essential and conserved nuclease–helicase involved in several DNA repair and replication pathways. A primary role of DNA2 is the removal of 5′ RNA-DNA flaps from Okazaki fragments during lagging strand synthesis in coordination with FEN1 [3]. In the absence of DNA2, excess single-stranded DNA can trigger DNA damage checkpoints [3,4]. Another well-known role of DNA2 is homologous recombination (HR)-mediated recovery or restart of stalled replication forks in both HR-dependent and HR-independent pathways [4,5,6,7]. DNA2 has also been shown to participate in end-resection during double-strand break (DSB) repair, specifically in replicating cells [3,8,9,10]; however, this is considered a secondary role to the involvement of DNA2 in replication fork recovery and restart. DNA2 overexpression has been shown to positively correlate with disease outcomes in several human cancers, leading to its exploration as a possible therapeutic target [6,10]. DNA2’s roles in replication and repair are accomplished by its two main enzymatic domains: a structure-specific nuclease, which plays the dominant role in DNA replication and repair [11,12] and two DNA-activated helicase/ATPases that shape a tunnel-like configuration that selectively guides single-stranded DNA to the nuclease active site [13].
Dna2 in Drosophila melanogaster (formerly mus109) is an X-linked gene that shows high protein sequence homology with DNA2 in humans, mouse, and Caenorhabditis elegans, including the presence of a nuclease domain and two helicase domains (helicase 1A and 2A) connected by a stalk protein [14]. All commercially available Dna2 mutant alleles (Dna2^lS^, Dna2^D1^, and Dna2^D2^) are hypersensitive to the alkylating agent methyl methanesulfonate (MMS), nitrogen mustard, and ɣ-radiation [14,15,16,17], which are known to cause bulky DNA adducts [18], DNA crosslinks [19], and DNA strand breaks through generation of reactive oxygen species [20], respectively. Dna2 mutants’ sensitivity to these reagents suggest that Dna2 is heavily involved in DNA replication and repair. Additionally, there are slight, but non-significant differences in MMS sensitivity between the alleles, possibly due to their domain-specific mutations: Dna2^lS^ contains a mutation in the nuclease domain and is homozygous lethal, while Dna2^D1^ and Dna2^D2^ contain mutations in the helicase domains and are considered hypomorphic (see Figure 3 in reference [14] for mutation map).
In this study, we further investigate the roles of Drosophila Dna2 in DNA replication and repair. We specifically focus on the alleles Dna2^D1^ and Dna2^D2^ to determine differential responses in the presence of the helicase 1A domain, which is present in Dna2^D1^ but not Dna2^D2^ [14]. Based on DNA2 function in other organisms, we predicted that Dna2 mutants would show sensitivity to mutagens that cause replication stress and show deficiencies in developmental timepoints that are highly dependent upon rapid and accurate DNA replication. Because it is common for deficiencies in replication and repair to impact longevity [21], we also investigated the impact of the Dna2 mutation on lifespan. Together, we show a connection linking the roles of Dna2 in replication to fly development and organismal physiology.
2. Materials and Methods
2.1. Fly Stocks and Maintenance
All fly stocks were maintained on solid cornmeal agar (BF Formula, Genesee Scientific, El Cajon, CA, USA) and kept at 25 °C under a 12 h:12 h light–dark cycle. Plates containing grape agar (Genesee Scientific) were used for egg collection. The following fly stocks were obtained from the Bloomington Drosophila Stock Center (BDSC): Dna2^D1^ (BDSC# 2320), Dna2^D2^ (BDSC# 2307), and Dna2^lS^ (BDSC# 4168). All experiments used mutants that were created in trans with the Dna2^lS^ (Dna2^lS/D1^ and Dna2^lS/D2^) allele to mitigate genetic background effects and because the transheterozygotes have shown stronger sensitivity to MMS [14]. w^1118^ flies served as the wildtype control when appropriate.
2.2. Mutagen Sensitivity Assays
Mutagen sensitivity assays were performed similarly to previous studies [14,22,23]. Briefly, Brood 1 (Day 0) was established by crossing 7–10 Dna2^lS^/FM7c females with 2–3 Dna2^D1^ or Dna2^D2^ males per vial. On day 3, the flies were flipped into new vials to establish Brood 2. On day 4, Brood 1 vials were treated with 250 μL of an aqueous mutagen solution: 20 mM Bleomycin (Thermo Fisher Scientific, Waltham, MA, USA), 50 μM Camptothecin (Thermo Fisher Scientific), 80 mM Hydroxyurea (Thermo Fisher Scientific), 0.00625–0.05% MMS (TCI America, Corona, CA, USA), 10% Nitrogen Mustard (Sigma Aldrich, St. Louis, MO, USA), 50 μM Potassium Bromate (Thermo Fisher Scientific), and 60 μM Topotecan (Apexbio Technology LLC, Houston, TX, USA). On day 5, the adult flies were discarded from Brood 2 vials, and on day 6, Brood 2 vials were mock-treated with 250 μL water. Adult offspring were collected every 2–3 days until day 18 (Brood 1) or day 21 (Brood 2) to ensure counting of only the first generation. Offspring were scored by sex and eye phenotype using the “bar-eye” phenotype on the FM7c balancer as an indicator of non-mutants and a “round-eye” phenotype (non-FM7c) for mutants. For each vial, relative survival was calculated by dividing the percentage of mutant flies in each treated vial (Brood 1) by the percentage of mutant flies in its corresponding control vial (Brood 2). Vials with fewer than 15 progeny in either Brood 1 or 2 were excluded from analysis. Statistical significance between Dna2^lS/D1^ and Dna2^lS/D2^ alleles was determined by unpaired t-test. Statistical significance for mutagen sensitivity was determined by unpaired t-tests comparing each mean relative survival with 100% survival, which would represent no sensitivity to the mutagen. Statistical analysis and graphing were performed using GraphPad Prism 10.5.0.
2.3. Mendelian Ratios
The offspring of the mock-treated controls in the mutagen sensitivity assays were used to determine Mendelian ratios, calculated by chi-square using the following classes of offspring: mutant females (Dna2^lS/D1^ or Dna2^lS/D2^), non-mutant females (Dna2^lS^/FM7c), and non-mutant males (FM7c/Y). The mutant male class (Dna2^lS^/Y) was excluded from calculation because of the lethality of the Dna2^lS^ allele. Calculations were performed using GraphPad Quickcalcs using an expected value of ⅓ for each class calculated. A small percentage (0.31–0.66%) of “round eye” (non-FM7c) males were observed. These males were presumed to be due to non-disjunction and not true Dna2^lS^/Y and were therefore omitted from calculations.
2.4. Fecundity
20–25 virgin female flies of the desired genotype (w^1118^ or Dna2^lS/D2^) were combined with 10 w^1118^ male flies in an embryo collection chamber (Dna2^lS/D2^ was prioritized due to the stronger fertility phenotype). All flies were 0–2 days old when added to the chamber. Chambers were maintained at 25 °C and fed three times daily (9:00 a.m., 12:00 p.m., and 5:00 p.m.) during the assay period. We used the 9:00 a.m.–12:00 p.m. collection for our fecundity assay data as collections taken later in the day were found to be highly variable, even for wild-type flies. Assay data from days 3–9 were used for data analysis. The number of eggs laid was divided by the number of females present and by the length of the collection (3 h) to calculate the average number of eggs laid per female per hour. Statistical analysis via unpaired t-test and graphing were performed using GraphPad Prism 10.5.0.
2.5. Egg Viability
Embryo chambers were set up with 0–2 day old females of the desired genotype (w^1118^, Dna2^lS/D1^, or Dna2^lS/D2^) and w^1118^ males. Collections began 2–3 days after the chambers were set up. Embryos were collected on grape-agar plates for a four-hour period, then transferred to fresh plates for accurate counting. At least 30 embryos were transferred per genotype per experiment. Embryos were incubated for an additional 24 h at 25 °C. The number of hatched and unhatched embryos were counted and embryo viability was calculated using the formula: (hatched embryos/total embryos) × 100. Statistical analysis via one-way ANOVA with multiple comparisons and graphing were performed using GraphPad Prism 10.5.0.
2.6. Ovary Dissection and Fixation
Virgin female flies of the desired genotype were collected and crossed with w^1118^ males. Flies were maintained in a well-fed vial at 25 °C for three days before dissection. Ovaries were dissected in PBS and fixed in 3.7% formaldehyde for 20 min. Fixed ovaries were washed three times in PBS-Tx before proceeding to immunostaining.
2.7. Immunostaining
Fixed ovaries were incubated in primary antibody solution overnight at 4 °C on a rotating platform. The primary antibody solution was prepared with mouse anti-ɣH2aV (DSHB) at a concentration of 1:500 in PBS-Tx + 5% NGS. After washing three times in PBS-Tx, ovaries were incubated in secondary antibody solution for two hours at room temperature on a rotating platform. The secondary antibody solution was prepared with goat anti-mouse AlexaFluor488 (Thermo Fisher Scientific) at a concentration of 1:1000 in PBS-Tx. After the secondary antibody was removed, ovaries were incubated for two minutes in PBS-Tx + 1 µg/mL DAPI then washed twice in PBS-Tx. Stained ovaries were mounted in ProLong (Thermo). Imaging was performed using a Zeiss AxioObserver.Z1 with Apotome 2.0. For quantification of ɣH2aV staining, ten w^1118^, five Dna2^lS/D1^, and ten Dna2^lS/D2^ germaria were assessed. Statistical analysis via Fisher’s exact test was performed using GraphPad Prism 10.5.0
2.8. Imaging and Quantification of Germarium Size
Imaging was performed using a Zeiss AxioObserver.Z1 with Apotome 2.0. Germaria were measured using the ZEN 3.5 software measurement tool. At least nine germaria were measured per genotype. Statistical analysis via one-way ANOVA with multiple comparisons and graphing were performed using GraphPad Prism 10.5.0.
2.9. Lifespan Assays
Dna2^lS^/FM7c females were crossed with Dna2^D1^ or Dna2^D2^ males. w^1118^ were used as a wild type control instead of Dna2^lS^/FM7c non-mutants to mitigate balancer effects on lifespan and physiology. Progeny were allowed to mate for 24–48 h following eclosion and then Dna2^lS/D1^ or Dna2^lS/D2^ females were separated under CO_2_ anesthetization and placed into narrow vials. Flies were transferred to new food and dead flies counted every 2–3 days for the duration of the experiment. Three biological replicates were completed. All analyses accounted for experimental batch structure by stratifying on experiment. Initially, stratified Cox proportional hazards regression was attempted, but Schoenfeld residual tests revealed violations of the proportional hazards assumption (global test p < 0.05). Therefore, stratified log-rank tests were performed as our primary analysis, which provide robust comparisons of survival distributions without assuming constant hazard ratios. Stratified log-rank tests were performed using R statistical software (version 4.5.0) with the survival package [24,25,26]. Claude Sonnet 4 (Anthropic, San Francisco, CA, USA) was used for assistance with R code development for survival analysis and statistical methodology guidance for handling experimental batch effects. Graphs were made using GraphPad Prism 10.5.0.
3. Results
3.1. Dna2 Responds to Specific Types of DNA Damage
We have previously confirmed sensitivity of all Dna2 alleles in this study to 0.05% MMS, including slight, but non-significant, differences between alleles possibly due to the presence of helicase 1A in Dna2^D1^ [14]. To further examine possible differences between the alleles, we exposed Dna2 mutants to lower concentrations of MMS. Dna2^lS/D1^ and Dna2^lS/D2^ mutants responded similarly to every concentration of MMS tested, displaying no sensitivity at 0.00625% MMS and near total lethality at 0.05% MMS (Figure 1A). There was no difference in MMS sensitivity between Dna2^lS/D1^ and Dna2^lS/D2^ mutants (p = 0.25 for interaction by two-way ANOVA) suggesting that there is no difference in Dna2 function between the two alleles when responding to MMS-induced damage.
The role of Dna2 in responding to DNA damage was further explored by treating Dna2 mutant and non-mutant heterozygous larvae with various mutagens that cause specific DNA damage and calculating relative survival to adulthood. Consistent with previous data in other models [7,10,27,28], Drosophila Dna2 mutants show low relative survival when treated with mutagens that cause replication fork stalling or collapse: topotecan (topoisomerase I inhibitor), hydroxyurea (ribonucleotide reductase inhibitor; depletes pool of available dNTPs) and nitrogen mustard (alkylating agent; causes DNA crosslinks) compared with 100% survival indicative of no sensitivity (Figure 1B). In contrast, Dna2 mutants were not sensitive to bleomycin (causes DSBs) or potassium bromate (causes strand breaks through oxidative stress). Relative survival for bleomycin-treated Dna2^D1^ larvae was significantly higher than 100% survival, which may indicate resistance to bleomycin-induced damage (Figure 1C, p < 0.05). We also observed a modest, but significant difference in survival between the two Dna2 alleles where Dna2^lS/D1^ showed higher survival when treated with topotecan and bleomycin, possibly indicating a helicase-specific role in responding to certain types of damage.
3.2. Dna2 Mutants Exhibit Reduced Viability and Reproductive Fitness
Dna2 mutants do not eclose in expected Mendelian ratios, even when factoring in the lethal Dna2^lS^ allele. Offspring from the parental cross (Dna2^lS^/FM7c females × Dna2^D1^/Y or Dna2^D2^/Y males) produce significantly fewer non-mutant males (FM7c/Y) and more females than expected for both Dna2^D1^ and Dna2^D2^ crosses (Table 1, Χ^2^ p < 0.0001), which may be due to viability deficiencies with the FM7c balancer [29,30]. We next investigated the role of Dna2 in reproductive fitness by assessing mutant fertility and fecundity. Eggs produced by Dna2 mutant females hatch at significantly lower rates than those produced by wild-type females, indicating reduced fertility (Figure 2A, p < 0.01 for Dna2^lS/D1^ and p < 0.0001 for Dna2^lS/D2^). Hatching frequency of eggs laid by Dna2^lS/D2^ females was also significantly lower than hatching frequency of eggs laid by Dna2^lS/D1^ females, suggesting a difference in allele function (Figure 2A). A follow-up fecundity assay for the more severely affected Dna2^lS/D2^ females revealed that Dna2 mutants produce fewer eggs compared to w^1118^ (Figure 2B, p < 0.001), suggesting a role for Dna2 in oogenesis.
3.3. Dna2 Is Required to Prevent Replication Stress in the Female Germline, Independent of Meiosis
To better understand the cause of reduced fecundity and fertility in Dna2 mutant females, we next examined ovarian morphology. We used immunostaining to label phosphorylated histone H2A variant (ɣH2Av), which is present during DNA damage and repair, and can also be used to visualize the resolution of induced double-strand breaks during meiosis [31,32,33,34,35]. Meiotic double-strand breaks are normally observed in regions 2A and 2B of the germarium, but should be absent in region 1, where germline stem cells and cystoblasts are undergoing mitotic divisions. Meiotic double-strand breaks should also be resolved by the time cysts transition into region 3 of the germarium [33]. As expected, we observed ɣH2Av-positive cells within regions 2A and 2B but not regions 1 or 3 of wild-type germaria, likely labeling cells undergoing meiosis I (Figure 3A). ɣH2Av-positive cells were also observed in regions 2A and 2B but not region 3 in Dna2 mutant germaria, suggesting that meiosis proceeds normally in these mutants (Figure 3B,C). However, we sometimes observed ɣH2Av-positive cells in region 1 of Dna2 mutant germaria (Figure 3C). We observed ɣH2Av-positive cells in region 1 in 40% of Dna2^lS/D1^ germaria and in 90% of Dna2^lS/D2^ germaria, which are significantly more than w^1118^ (p < 0.0001 by Fisher’s exact test). ɣH2Av staining in mitotically dividing cells in region 1 suggests DNA damage in germline stem cells and/or mitotically dividing cystoblasts. We also noted abnormal morphology of germaria and reduced overall size of mutant germaria (Figure 3C,D).
3.4. Dna2 Mutations Do Not Reduce Lifespan
We examined the effect of Dna2-deficiency in adults by measuring fly lifespan. There was great variability across our three biological replicates, with each genotype and replicate showing different survivorship curves for w^1118^, Dna2^lS/D1^, and Dna2^lS/D2^ (Figure 4, p < 0.0001 by stratified log-rank analysis). Surprisingly, Dna2 mutants showed longer lifespans compared to w^1118^, with Dna2^lS/D2^ flies living longer than Dna2^lS/D1^.
4. Discussion
Proper DNA replication is critical for organismal survival, especially during early stages of development. Our study shows that Dna2 is required during oogenesis, as well as at critical developmental timepoints in early embryogenesis and metamorphosis. This requirement, which was assessed via loss-of-function analysis (sufficiency was not tested), is evidenced by reduced fecundity, fertility, and survival of larvae to adulthood following exposure to mutagens that induce replication stress. Additionally, we observed increased ɣ-H2Av in mitotically dividing regions in Dna2 mutant germaria indicating that Dna2 may prevent damage in these tissues. Together, these findings indicate that Dna2 responds to stress during DNA replication in Drosophila.
Both the nuclease and helicase activity of Dna2 are critical for promoting replication fork recovery. The DNA2 nuclease removes 5′ RNA-DNA flaps during Okazaki fragment processing in coordination with the nuclease FEN1 [3,11]. DNA2 also promotes replication restart through degradation of reversed replication forks [36] and DNA end resection to initiate homology-directed repair of double-strand breaks along with RecQ helicases (reviewed in [3]), while protecting against excessive replication intermediates [5]. Evidence for essential helicase-dependent roles of Dna2 is more limited, showing that Dna2 supports homologous recombination (HR) in replication fork recovery in yeast [7] and at centromeric regions in human cells [37]. Interestingly, helicase activity is only strongly observed when nuclease activity is attenuated [8,38], further implicating the nuclease as the dominant enzymatic activity of Dna2. Our transheterozygous Dna2 mutants contain one allele that lacks nuclease sequence (Dna2^lS^) and one allele that contains the nuclease (Dna2^D1^ or Dna2^D2^). Despite the presence of one copy of the nuclease domain, these mutants are still hypersensitive to DNA damage that likely involves nuclease-dependent repair. This may suggest that the nuclease domain is compromised by improper protein folding in the Dna2^D1^ and Dna2^D2^ alleles, which will need to be investigated through biochemical analysis of the mutant alleles. The mutant alleles also differ in the presence of the helicase 1A domain where Dna2^D1^ contains the helicase and Dna2^D2^ does not. We observed modest, but significantly higher relative survival of Dna2^lS/D1^ mutants treated with topotecan and bleomycin, possibly indicating helicase-dependent roles in responding to breaks at replication forks.
DNA2 responds to replication stress through restarting/resolving stalled replication forks or attending to DSBs caused by replication fork collapse. DNA2 has been shown to respond to DSBs in mitotic cells through its interactions with Exo1, MRN/X, and sgs1/Blm [12,39,40]. In the absence of DNA2, persistent replication intermediates accumulate, which can interfere with chromosome segregation and metaphase plate formation during mitosis [5,7,10,37,41]. DNA2 also responds to replication stress during meiosis [42], however the evidence is more limited due to redundancy with required proteins such as Exo1 and sgs1 [40]. Our data show that there is no increase in DSBs in the meiotically dividing cells in region 2 of Dna2 mutant germaria. However, we observed anincrease in DSBs in mitotically dividing cells in region 1 of germaria indicating a Dna2 response to replication stress during mitosis only. Similarly, the low hatching frequency of Dna2 mutant eggs is likely due to replication stress during syncytial division in early embryogenesis, similar to mutant phenotypes observed for other repair genes [22,43,44,45,46]. These differences in mechanism may be due to the inherent differences between mitotic and meiotic DSBs: meiotic DSBs are tightly regulated and processed by meiosis-specific nucleases and recombinases, in which Dna2 is less essential. Therefore, Dna2 may be limited predominantly to mitotic cells where it is required for end resection of breaks caused by replication fork collapse.
DNA2 has also been shown to participate in end-resection during DSB repair [12], specifically in replicating cells [10]. While likely not its primary mechanism, Dna2 may also be responding to DSBs caused by collapsed replication forks in Drosophila. Our evidence similarly suggests involvement of Dna2 in DSB repair based on induction of ɣH2Av foci in region 1 in the germaria and mutant sensitivity to ɣ-radiation [15,17]. Our Dna2 mutants are also sensitive to topotecan which in doses used in this study, has been shown to induce sensitivity in HR-deficient flies, further indicating involvement in DSB repair [47]. Surprisingly, our Dna2 mutants are not sensitive to the radiomimetic bleomycin, which is well-known to cause DSBs. The damage mechanism of bleomycin has been shown to be dose-dependent where low doses (<10.6 μM) incite more DSBs based on involvement with Rad52 and high doses (>10.6 μM) induce single-strand breaks (SSBs) based on involvement with Rad6 [48,49,50]. Due to differences in dosing and organism, it is difficult to make a direct comparison, however it is possible that our dose of 20 μM is inducing more SSBs, which are repaired by a Dna2-independent mechanism. It is also possible that DSBs are being induced and repaired by another mechanism in Drosophila, such as the end-resection proteins CtIP and Rif1 [51,52], as Dna2 is most active at replication forks in dividing cells and might not be an important player in canonical DSB repair.
In many cases, a mild amount of stress can elicit a positive protective response. For example, mild stress induction has been shown to extend lifespan in flies and C. elegans [53,54,55,56]. More specific to this work, flies lacking the DNA repair protein, WRNexo, show both increased antioxidant activity and thermal stress resistance, suggesting lack of WRNexo may heighten other stress responses [23]. Despite known roles of DNA2 in DSB repair and base excision repair [3], we observed surprisingly high survival rates of Dna2 mutants when treated with both bleomycin, which causes DSBs, and potassium bromate, which causes oxidative stress. Additionally, Dna2^lS/D1^ showed significantly higher than 100% relative survival, which may indicate potential resistance to bleomycin-induced damage. Therefore, while Dna2 may not respond directly to DSBs or oxidative DNA damage, Dna2-deficiency may heighten other protective mechanisms to allow greater survival when exposed to these exogenous stressors.
Heightened stress responses may also be a factor in differences in longevity in Dna2 mutants. We had hypothesized that lack of Dna2 would reduce fly lifespan by limiting repair capacity. Instead, we found that Dna2 mutants have significantly longer lifespans compared to wild-type flies in an allele-dependent manner. Interestingly, low fecundity has been observed in long-lived flies, suggesting a trade-off in resource allocation from reproduction to longevity [57]. Therefore, low fecundity in Dna2 mutants could lower their reproductive cost leading to increased longevity. Still, we interpret these results with caution as it is rare for DNA repair deficiency to extend lifespan and we acknowledge that genetic background differences in our w^1118^ controls as well our Dna2 alleles may have contributed to our results.
Together our results support a requirement for Drosophila Dna2 in responding to replication stress, both caused by DNA damaging reagents and during developmental processes that require rapid DNA replication. These findings are consistent with those in other organisms and support using Drosophila as a model for studying Dna2 function. We expect that these findings will support future studies into the molecular roles of Dna2 in Drosophila and help build the foundation for understanding how this protein contributes to genome maintenance and disease.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hoeijmakers J.H.J. DNA Damage, Aging, and Cancer N. Engl. J. Med.20093611475148510.1056/NEJ Mra 080461519812404 · doi ↗ · pubmed ↗
- 2Jackson S.P. Bartek J. The DNA-damage response in human biology and disease Nature 20094611071107810.1038/nature 0846719847258 PMC 2906700 · doi ↗ · pubmed ↗
- 3Zheng L. Meng Y. Campbell J.L. Shen B. Multiple roles of DNA 2 nuclease/helicase in DNA metabolism, genome stability and human diseases Nucleic Acids Res.202048163510.1093/nar/gkz 110131754720 PMC 6943134 · doi ↗ · pubmed ↗
- 4Appanah R. Jones D. Falquet B. Rass U. Limiting homologous recombination at stalled replication forks is essential for cell viability: DNA 2 to the rescue Curr. Genet.2020661085109210.1007/s 00294-020-01106-732909097 PMC 7599155 · doi ↗ · pubmed ↗
- 5Falquet B. Ölmezer G. Enkner F. Klein D. Challa K. Appanah R. Gasser S.M. Rass U. Disease-associated DNA 2 nuclease–helicase protects cells from lethal chromosome under-replication Nucleic Acids Res.2020487265727810.1093/nar/gkaa 52432544229 PMC 7367196 · doi ↗ · pubmed ↗
- 6Hudson J.J.R. Rass U. DNA 2 in Chromosome Stability and Cell Survival—Is It All about Replication Forks?Int. J. Mol. Sci.202122398410.3390/ijms 2208398433924313 PMC 8069077 · doi ↗ · pubmed ↗
- 7Ölmezer G. Levikova M. Klein D. Falquet B. Fontana G.A. Cejka P. Rass U. Replication intermediates that escape Dna 2 activity are processed by Holliday junction resolvase Yen 1Nat. Commun.201671315710.1038/ncomms 1315727779184 PMC 5093310 · doi ↗ · pubmed ↗
- 8Levikova M. Pinto C. Cejka P. The motor activity of DNA 2 functions as an ss DNA translocase to promote DNA end resection Genes Dev.20173149350210.1101/gad.295196.11628336515 PMC 5393063 · doi ↗ · pubmed ↗