Genome-Wide Identification and Expression Analysis of the CaM Gene Family in Tree Peony (Paeonia ostii) During the Pistil Pollination Process
Guodong Zhao, Shuran Lv, Yuxin Zhao, Yuying Li, Xiaogai Hou

TL;DR
This study identifies and analyzes calmodulin genes in tree peony during pollination, revealing their expression patterns and potential roles in the process.
Contribution
The study provides the first genome-wide identification and expression analysis of the CaM gene family in tree peony during pollination.
Findings
Six CaM family members were identified, located on three chromosomes and one non-chromosome.
Gene promoters of the CaM family contain numerous hormone and stress response elements.
CaM gene expression in pistil tissue increases then decreases during pollination, suggesting functional involvement.
Abstract
Tree peony is an important horticultural plant with both ornamental and oil value. The tree peony genome and databases were used to search for calmodulin family genes to explore their function in the pollination of tree peony. The CaM gene family was identified, and then the basic protein characteristics of the family members, such as gene structure, isoelectric point, molecular weight, subcellular localization, and conserved protein domain, were analyzed. The expression levels of these genes in the pistil tissue of Paeonia ostii ‘Fengdan’ at different developmental stages after pollination were also analyzed. Further, qRT-PCR was used to detect the expression levels of six PsCaMs during the development process of the pistil under bee pollination conditions. The results showed that there were six CaM family members located on three chromosomes and one non-chromosome. There were a large…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
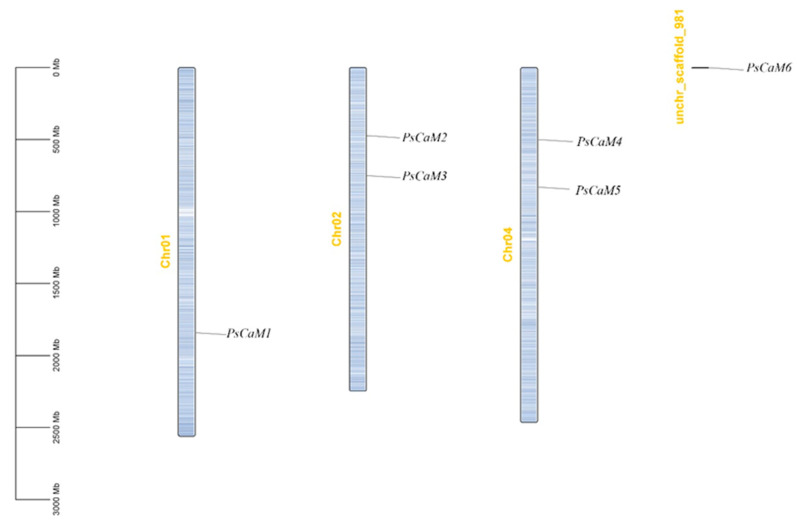 Figure 1
Figure 1 Figure 2
Figure 2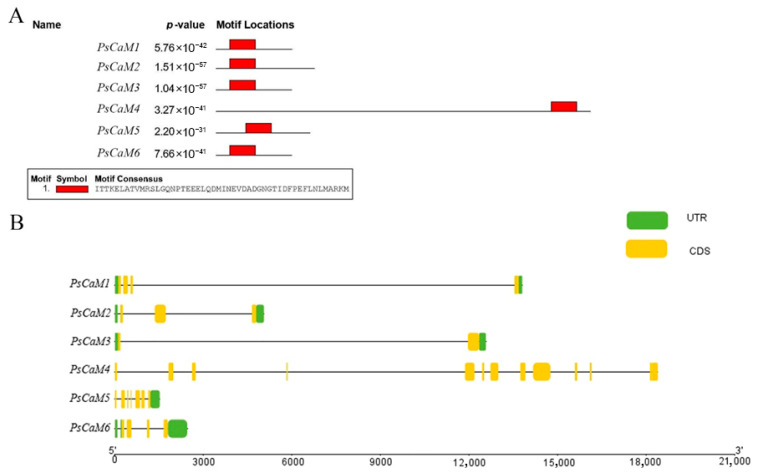 Figure 3
Figure 3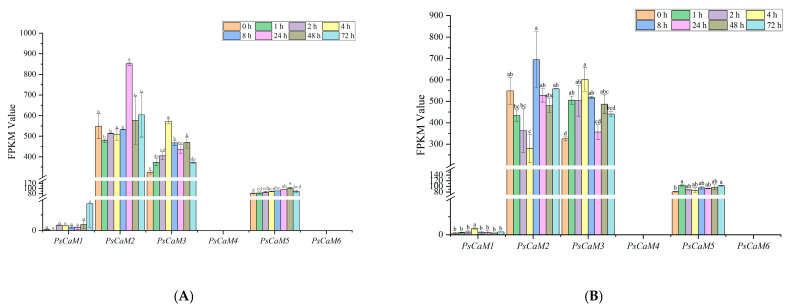 Figure 4
Figure 4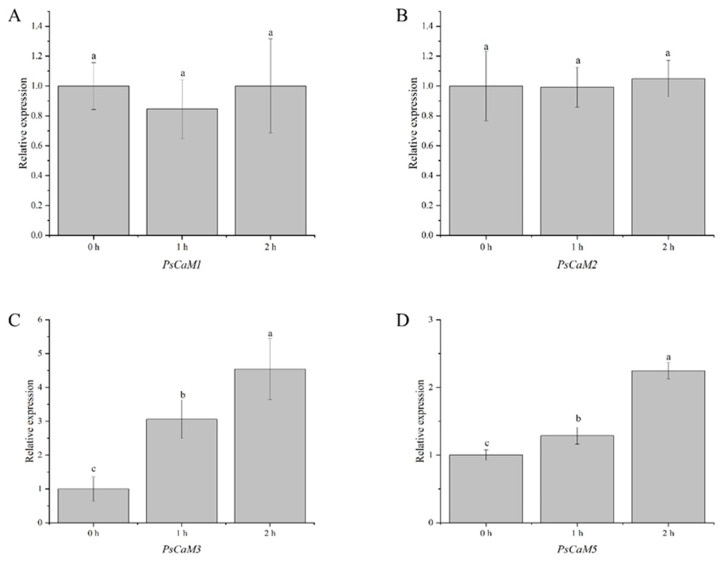 Figure 5
Figure 5- —Key Research Project Sub-Topic of the National Key R&D Program
- —Henan Province’s Science and Technology Research Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Reproductive Biology · Plant Molecular Biology Research · Plant biochemistry and biosynthesis
1. Introduction
Paeonia section Moutan DC., a woody plant belonging to the family Paeoniaceae, is widely recognized for its large, full blossoms, diverse flower forms, vibrant colors, and intense fragrance. It is revered as the “King of Flowers” and a symbol of wealth and prosperity, with significant ornamental value [1]. In addition to its aesthetic appeal, tree peony has emerged as a novel woody oil crop with substantial economic potential [2]. The seed oil of tree peony contains up to 92.26% unsaturated fatty acids, including over 42% α-linolenic acid. Therefore, the exploration and utilization of Paeonia germplasm rich in α-linolenic acid are of great application value [3].
Most species within the Paeonia section Moutan exhibit self-incompatibility [4]. This is particularly pronounced in Paeonia ostii ‘Fengdan’, where the self-pollination seed set rate is extremely low, with an average of only 0.4 seeds per flower [5]. Bee pollination has been shown to significantly improve seed yield and mitigate the adverse effects of late spring frost on seed development [6]. Studies have indicated that pollination is influenced by various chemical factors. For example, pollen germination increases with sucrose concentrations up to 150 g·L^−1^. Moreover, 80 mg·L^−1^ boric acid significantly enhances pollen germination [7]. Boric acid facilitates Ca^2+^ influx by interacting with calcium channels on the plasma membrane, triggering intracellular calcium mobilization and opening internal Ca^2+^ stores, thereby elevating cytosolic Ca^2+^ levels [8].
Calcium (Ca^2+^) plays a crucial role in pollen tube growth. At low Ca^2+^ concentrations, pollen tube elongation slows or ceases, whereas excessive Ca^2+^ levels also inhibit growth [9]. Prior to pollen tube germination, Ca^2+^ accumulates near the germination pore. Following germination, a calcium gradient forms along the pollen tube, regulating vesicle trafficking, fusion, and secretion from the Golgi apparatus. This ensures a continuous supply of cell wall and plasma membrane materials to the tube tip, supporting its polarized growth [10].
Calmodulin (CaM) proteins, ubiquitous in plants, are encoded by multigene families producing identical or highly similar proteins [11]. CaMs are central regulators in a wide range of physiological processes, including cell morphogenesis, division, flowering, autophagy, protoplast wall regeneration, and pollen tube germination [12]. Additionally, CaMs are involved in responses to abiotic stresses such as mechanical stimulation, drought, chilling [13], heat, and high salinity [14]. With the advent of whole-genome sequencing, the CaM gene family has been identified and characterized in many plant species. In Arabidopsis, AtCaM4 interacts with PALT1 to repress CBF gene expression, negatively regulating cold tolerance [15], while AtCaM2 regulates pollen tube orientation and signaling by modulating calcium levels during growth [16]. In maize, CaM promotes pollen germination and tube elongation, whereas CaM inhibitors such as CPZ, W7, TFP, and compound 48/80 strongly inhibit these processes [17]. In Cucumis sativus, CsCaM3 enhances thermotolerance by upregulating HSP70 and HSP90 expression and alleviating membrane damage [18]. In apple, MdCaM2 interacts with MdCRF4 in a Ca^2+^-dependent manner, phosphorylating and suppressing MdCRF4-induced MdACS1 transcription, thereby reducing ethylene biosynthesis and delaying fruit ripening [19]. In rice, OsCaM interacts with OsCNGC13, a channel critical for transmitting pollen through the style; loss of OsCNGC13 impairs pollen transport to the ovary [20]. These findings demonstrate that CaMs participate in complex calcium-mediated signaling pathways, playing indispensable roles in plant development, stress responses, and hormone signaling.
In this study, pistil tissues of Paeonia ostii ‘Fengdan’ at different developmental stages after bee pollination were used as experimental materials. Based on the Paeonia ostii genome and other public databases, we identified CaM gene family members in tree peony, analyzed their physicochemical properties, conserved motifs, phylogenetic relationships, chromosomal locations, and expression patterns. This work lays a foundation for further investigation into the molecular mechanisms underlying pollination in tree peony.
2. Materials and Methods
2.1. Plant Materials
Experimental materials were collected from the Tree Peony Experimental Base of Henan University of Science and Technology (112°25′16.28″ E, 34°36′8.00″ N). Twelve-year-old healthy Paeonia ostii ‘Fengdan’ plants with consistent growth were selected. Bee pollination was performed prior to flowering in April 2023. Pistil tissues of Paeonia ostii ‘Fengdan’ were harvested at 0 h (non-pollinated), 1 h, and 2 h after pollination. Five pistil tissues from each plant were collected, and the samples were immediately frozen in liquid nitrogen and stored at –80 °C for subsequent analysis.
2.2. Experimental Methods
2.2.1. Identification of CaM Gene Family Members and Protein Characterization in Tree Peony
Seven annotated Arabidopsis thaliana CaM protein sequences were downloaded from the TAIR database (http://www.arabidopsis.org/, accessed on 20 April 2024). These sequences were used as queries for BLAST searches against the Paeonia genome [21] using NCBI-BLASTp (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 20 April 2024). Candidate homologous sequences with >50% similarity was retained. After removing redundant sequences, putative CaM family members in Paeonia were identified.
The physicochemical properties of PsCaM proteins, including isoelectric point, molecular weight, and conserved domains, were predicted using ExPASy (https://web.expasy.org), SMART (http://smart.embl-heidelberg.de/), Pfam (http://pfam.xfam.org/), and TBtools software (v. 2.031). Subcellular localization was predicted using Cell-Ploc 2.0 (http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/, accessed on 25 May 2024), BUSCA (http://busca.biocomp.unibo.it), and YLoc (https://github.com/KohlbacherLab/YLoc, accessed on 25 May 2024). Chromosomal localization was visualized using TBtools. All the above-mentioned online websites and software use default parameters.
2.2.2. Phylogenetic Analysis of CaM Gene Family Members
Multiple sequence alignments of CaM proteins from Paeonia and Arabidopsis were performed using ClustalX (www.clustal.org, accessed on 16 June 2024). A phylogenetic tree was constructed using the Neighbor-Joining (NJ) method implemented in MEGA software (www.megasoftware.net, accessed on 19 June 2024). The program parameters were set as follows: Bootstrap replication times were 1000, Poisson model, complete state deletion, and other parameters were set to default.
2.2.3. Conserved Motif and Gene Structure Analysis of CaM Genes
The conserved motifs of PsCaM proteins were identified using MEME (https://meme-suite.org) based on the expectation-maximization (EM) algorithm. The results were visualized using TBtools. Gene structures and cis-acting elements located in the 2000 bp upstream promoter regions of candidate genes were also analyzed using TBtools (v. 2.031), and key elements were selected for further visualization.
2.2.4. Expression Analysis of PsCaM Genes Based on RNA-Seq Data
Based on previously obtained RNA-seq data from pistil tissues of P. ostii ‘Fengdan’ under bee and self-pollination conditions, the time expression patterns of PsCaM genes were analyzed during pistil development.
2.2.5. qRT-PCR Validation of PsCaM Gene Expression
Total RNA was extracted from pistil tissues of P. ostii ‘Fengdan’ at 0, 1, and 2 h after bee pollination, with 2 g used for each sample, using the RNAprep Pure Plant Kit for polysaccharide- and polyphenol-rich tissues (DP441, TIANGEN Biotech, Beijing, China). RNA concentration and quality were assessed using 1.0% agarose gel electrophoresis and a NanoDrop spectrophotometer.
qRT-PCR was performed using the Evo M-MLV Reverse Transcription Kit (AG11705, Accurate Biology, Hunan, China) and SYBR Green Pro Taq HS Premix Kit (AG11701, Accurate Biology, Hunan, China) on the BIO-RAD CFX Connect Real-Time PCR Detection System. Each sample was subjected to a reverse transcription reaction using 100 ng of RNA. Gene-specific primers for PsCaMs were designed using Primer Premier (Table 1), with PsEF-1α as the internal reference gene, and the final volume of the qRT-PCR reaction was 20 μL [22]. Three biological replicates were included for each sample. Relative gene expression levels were calculated using the 2^−ΔΔCt^ method, and statistical significance was determined via one-way analysis of variance (ANOVA).
3. Results
3.1. Identification and Physicochemical Properties of CaM Gene Family Members in Paeonia suffruticosa
A total of six CaM family members were identified from the P. ostii genome (Table 2). Based on sequence homology with A. thaliana CaMs, these genes were designated as PsCaM1 to PsCaM6. The amino acid lengths of PsCaMs ranged from 450 to 2217, with PsCaM4 being the longest. The average molecular weight was approximately 63,719.06 ± 24,269.72 Da, and the average isoelectric point (pI) was 5.21 ± 0.05. The instability indices ranged from 19.16 to 41.17, the aliphatic indices from 29.11 to 36.20, and the grand average of hydropathicity (GRAVY) values fell between 0.67 and 0.75, indicating that all PsCaMs are hydrophobic proteins. Subcellular localization predictions suggested that five of the six PsCaMs were primarily localized to the plasma membrane, with PsCaM6 uniquely localized to the vacuole.
3.2. Chromosomal Localization of PsCaM Genes
Paeonia ostii has relatively few chromosomes (2n = 10) but large chromosome sizes. The six PsCaM genes were distributed unevenly across three chromosomes: Chr1, Chr2, and Chr4 (Figure 1). Specifically, PsCaM1 was located on Chr1; PsCaM2 and PsCaM3 on Chr2; and PsCaM4 and PsCaM5 on Chr4. PsCaM6 was not anchored to any chromosome in the current genome assembly.
3.3. Phylogenetic Analysis of CaM Family Members
Multiple sequence alignment revealed high conservation among the 13 CaM proteins from Paeonia ostii and A. thaliana (Figure 2A). These CaMs were classified into six groups. The six PsCaMs were distributed across groups 1, 4, 5, and 6, containing 2, 2, 1, and 1 members, respectively. PsCaM2 and PsCaM3 showed high sequence similarity with AtCaM6 and AtCaM7; PsCaM1 and PsCaM6 were more closely related to AtCaM1 and AtCaM4. In contrast, PsCaM4 and PsCaM5 showed lower homology with other CaM proteins (Figure 2B).
3.4. Conserved Motif and Gene Structure Analysis of PsCaMs
Motif analysis using MEME revealed that all six PsCaMs shared a highly conserved motif with the amino acid sequence: ITTKELATVMRSLGQNPTEEELQDMINEVDADGNGTIDFPEFLNLMARKM with a maximum p-value of 2.20 × 10^–31^ (Figure 3A), confirming their identity as CaM family members. The motifs in PsCaM1, PsCaM2, PsCaM3, PsCaM5, and PsCaM6 were mainly located in the central regions of the protein sequences.
Gene structure analysis based on genome annotation showed that PsCaM4 had the longest gene length (~18,000 bp) and included 13 CDS regions, though lacking both 5′UTR and 3′UTR. PsCaM5 was the shortest (~1500 bp), containing 7 CDS regions and a 5′UTR but lacking a 3′UTR. The remaining members contained 2–4 CDS regions and both UTRs (Figure 3B).
3.5. Cis-Regulatory Elements in Promoters of PsCaMs
Cis-element analysis of the 2000 bp upstream promoter regions revealed 11–19 regulatory elements per PsCaM gene, including hormone-responsive (ABRE), low-temperature responsive (LTR), light-responsive (G-box), drought-inducible (MBS), and anaerobic-inducible (ARE) elements (Table 3). All PsCaMs possessed abundant light- and hormone-responsive elements. Only PsCaM3 and PsCaM5 had drought-responsive elements, suggesting that most PsCaMs may respond to phytohormone signaling and light stimuli.
3.6. Expression Patterns of PsCaM Family Genes During Pistil Development Following Pollination in P. ostii ‘Fengdan’
RNA-seq data from previous experiments indicated that, except for PsCaM4 and PsCaM6, the remaining PsCaMs were expressed in pistil tissues under both bee and hand pollination. Expression levels varied among members (Figure 4). PsCaM2 and PsCaM3 exhibited higher expression across developmental stages. PsCaM2 showed a dynamic pattern of initial downregulation followed by a sharp increase and subsequent decline, peaking at 24 h post-bee pollination (p < 0.05; Figure 4A) and at 8 h under hand pollination (p < 0.05; Figure 5B). PsCaM3 showed a peak expression at 4 h post-pollination under both conditions (p < 0.05; Figure 4A,B).
qRT-PCR analysis validated the expression of PsCaMs at 0, 1, and 2 h post bee pollination (Figure 5), consistent with RNA-seq results. PsCaM1 and PsCaM2 exhibited an initial decrease followed by an increase, but changes were not statistically significant (p > 0.05; Figure 5A,B). In contrast, PsCaM3 and PsCaM5 showed significantly increased expression, peaking at 2 h post-pollination (p < 0.05; Figure 5C,D), suggesting a potential shared regulatory function. PsCaM4 and PsCaM6 showed no detectable expression at any time point post-pollination.
4. Discussion
Calmodulin (CaM) is an acidic protein composed of 148 amino acids, with a molecular mass ranging from 16.7 to 16.8 kDa and a pI of approximately 4.0 [23]. It lacks easily oxidizable residues such as cysteine and tryptophan, which confer high thermal stability, even retaining bioactivity at 90 °C. CaMs were highly conserved across eukaryotes, with up to 91% sequence similarity between plant and vertebrate CaMs, 61% similarity with yeast, and 84–100% similarity among plants and algae [24]. Structurally, CaM contains four Ca^2+^-binding EF-hand motifs, seven α-helices, and two antiparallel β-sheets [25]. The presence of only four EF-hand domains and no additional functional motifs is a defining feature of the CaM gene family [26]. Although CaM genes have been studied in many species, little is known about their function and number in tree peony. This study analyzed the sequence characteristics of CaM gene members and their expression patterns during pollination of tree peony, which laid a foundation for the study of the CaM gene in tree peony.
CaMs were widely distributed in various species. In A. thaliana, seven CaM genes encode four isoforms differing by only 1–5 amino acids [27]. In rice, five genes encode three isoforms with >97% similarity [28]. And in wheat, 40 genes encode 13 highly similar isoforms [29]. Similarly, three CaM genes in grape belong to a single subfamily, and analogous CaM gene families have also been reported in soybean, tobacco, and maize [30,31,32,33]. In this study, six PsCaMs were identified from the P. ostii ‘Fengdan’, of which PsCaM1, PsCaM2, PsCaM3, and PsCaM6 exhibited high sequence conservation. We performed multiple sequence alignments between PsCaM and AtCaM, and the results showed that PsCaM7 had the closest homology with AtCaM2. Among them, PsCaM4 showed moderate similarity, while PsCaM5 was more divergent. All six contained four canonical EF-hand motifs without additional conserved domains and were primarily localized to the cytoplasm or plasma membrane, which is consistent with the characteristics of the typical calmodulin family [14]. These findings lay the groundwork for future functional investigations of CaMs in Paeonia ostii ‘Fengdan’.
CaM gene expression varies among developmental stages and tissues. In Pyrus bretschneideri, CaMs were differentially expressed in young leaves, mature leaves, flower buds, and petals, which is likely to reflect the diversity of their functions [34]. In fig (Ficus carica), five CaM genes show distinct responses to cold, light, drought, and ABA treatments, suggesting coordinated regulation among family members under stress conditions [35]. CaM was also implicated in multiple aspects of pollen germination. Although CaMs were ubiquitous in the cytoplasm, their activation strictly depends on Ca^2+^ binding, leading to localized activation in regions with high Ca^2+^ gradients, such as pollen tube tips, where active CaM can negatively regulate targets like cyclic nucleotide-gated channels (CNGCs) [36]. Furthermore, CaM interacts with Ca^2+^-dependent kinases to regulate pollen tube growth and directional guidance [37]. In this study, under bee pollination, PsCaM1 and PsCaM2 showed transient downregulation followed by upregulation in pistil tissues, whereas PsCaM3 and PsCaM5 were significantly upregulated post-pollination, likely reflecting roles in post-transcriptional or translational regulatory networks, warranting further investigation [38]. We analyzed the expression pattern of the CaM gene in different stamen development stages of tree peony under bee pollination and self-pollination conditions. It was found that the expression patterns of CaM2 and CaM3 in self-pollinated and honey-pollinated tree peony were slightly different, suggesting that they may be involved in the regulation of the tree peony bee pollination process.
Hormone signals also play a significant role in pollen germination and pollen tube growth, and there is a close interaction between calcium signals and hormone signals. Current data indicate that pollen germination, both in vitro and in vivo, is accompanied by endogenous plant hormones. The identification of CaM family members in peonies has revealed 2–7 hormone responses, suggesting that PsCaMs may be involved in the hormone-induced response process of tree peonies, thereby facilitating the participation of calcium signals in regulating aspects such as pollen germination and pollen tube elongation to regulate the growth and development of plants. This provides clues for further in-depth molecular mechanism research on these genes in the future.
5. Conclusions
Six PsCaMs were identified and obtained from the genome of the tree peony. Each member was unevenly located on three chromosomes. A large number of hormone response and stress response elements were present in the promoters of these six PsCaMs. During the development process of the pistil tissue of Paeonia ostii ‘Fengdan’ after pollination, the expression levels of CaM family genes showed a trend of first increasing and then decreasing, which may be related to their functions during the pollination process.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Li Y.Y. Guo L.L. Wang Z.Y. Zhao D.H. Guo D.L. Carlson J.E. Yin W.L. Hou X.G. Genome-wide association study of twenty-three flowering phenology traits and four floral agronomic traits in tree peony (Paeonia section Moutan DC.) reveals five genes known to regulate flowering time Hortic. Res.202310 uhac 26310.1093/hr/uhac 26336793754 PMC 9926158 · doi ↗ · pubmed ↗
- 2Li Y.Y. Guo Q. Zhang K.Y. Wang H. Jia C.S. Guo D.L. Guo L.L. Hou X.G. Dormancy-release, germination and seedling growth of Paeonia ostii ‘Fengdan’ seeds under measures of physical and chemical treatment and sowing P Lo S ONE 202217 e 027076710.1371/journal.pone.027076735789330 PMC 9255771 · doi ↗ · pubmed ↗
- 3Zhang K.Y. Wang X. Bao J.Y. He X.N. Leri Y. He C.L. Hou X.G. Bumblebee pollination ensures the stability of both yield and quality of the woody oil crop Paeonia ostii ‘Fengdan’Basic Appl. Ecol.202479384510.1016/j.baae.2024.03.001 · doi ↗
- 4Zhang K.L. Yao L.J. Zhang Y. Baskin J.M. Baskin C.C. Xiong Z.M. Tao J. A review of the seed biology of Paeonia species (Paeoniaceae), with particular reference to dormancy and germination Planta 201924929130310.1007/s 00425-018-3017-430276471 · doi ↗ · pubmed ↗
- 5Wang H. Wei S. He Y.L. Wang X.H. Li Y.Y. Wei D.F. Wang Z.Y. Guo L.L. Shaaban M. Hou X.G. Characterization of agronomic and seed oil features for different cultivars of tree peony Plants 202312311210.3390/plants 1217311237687358 PMC 10490233 · doi ↗ · pubmed ↗
- 6Zhang K.Y. He C.L. Hou X.G. Influence of pollination methods on fruit development, fruit yield and oil quality in oil tree peony Sci. Hortic.202229511087710.1016/j.scienta.2022.110877 · doi ↗
- 7Shi J. Wang X.Y. Zhang S.L. Huo Z.P. Liu S.D. The influence of culture medium components on the germination of peony pollen and the growth of pollen tubes J. Henan Univ. Sci. Technol. (Nat. Sci. Ed.)20133475–80+1
- 8Wang H. Zhang C.H. Huang Y.L. Research progress on factors affecting plant pollen and preservation methods Chin. Agric. Sci. Bull.2023394954