Genome-Wide Identification and Expression Analysis of Adenylate Kinase Family Members in Pepper Under Abiotic Stress
Bingxue Han, Kexu Sun, Jingyuan Zhou, Junwei Xu, Aidi Feng, Xiaohong Zhao

TL;DR
This study identifies and analyzes 15 adenylate kinase genes in pepper and their roles in responding to environmental stress.
Contribution
First comprehensive analysis of the CaADK gene family in pepper under abiotic stress.
Findings
Pepper has 15 ADK genes grouped into six conserved evolutionary clusters.
CaADK9 is ubiquitously expressed and upregulated under salt stress.
CaADK11 and CaADK12 are downregulated under low-temperature and salt stress.
Abstract
Adenylate kinase (ADK), a highly conserved and ubiquitously expressed enzyme in plants, serves as a critical regulator of cellular energy homeostasis and abiotic stress adaptation. While ADK families have been characterized in model species (e.g., Arabidopsis thaliana, Oryza sativa) and crops such as tomato (Solanum lycopersicum), the molecular features and stress-responsive roles of ADK genes in pepper (Capsicum annuum L.) remain uncharacterized. Here, we systematically identified 15 ADK genes in pepper (named by chromosomal location) and revealed their evolutionary relationships with orthologs from four plant species, clustering into six conserved groups. The promoters of CaADKs were found to contain cis-acting elements linked to stress responses, including those responsive to abscisic acid, gibberellin, and low-temperature conditions. Tissue-specific expression profiling highlighted…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
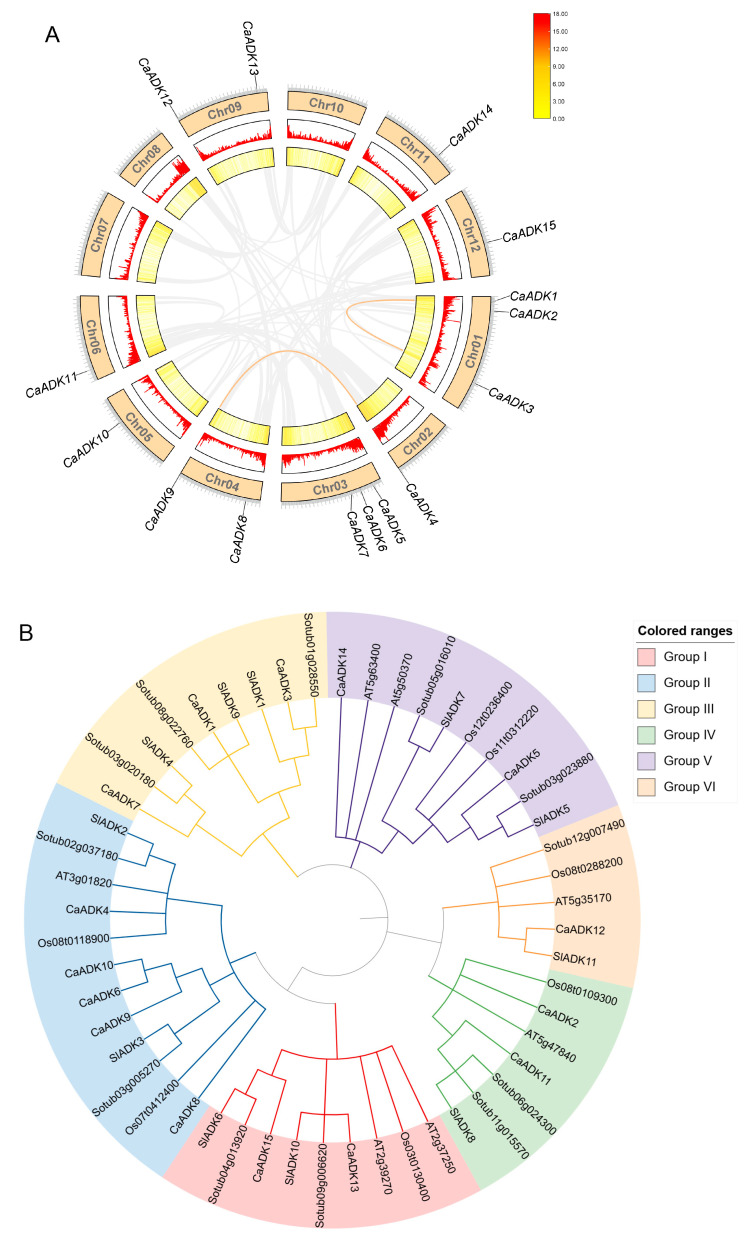 Figure 1
Figure 1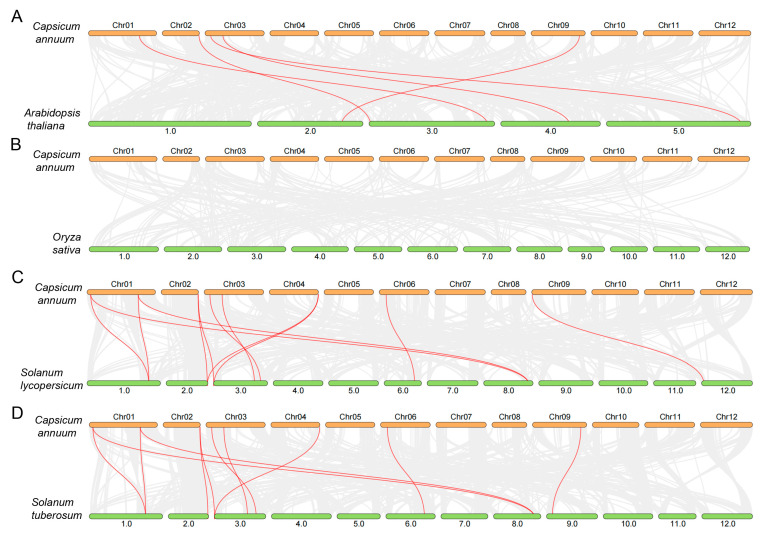 Figure 2
Figure 2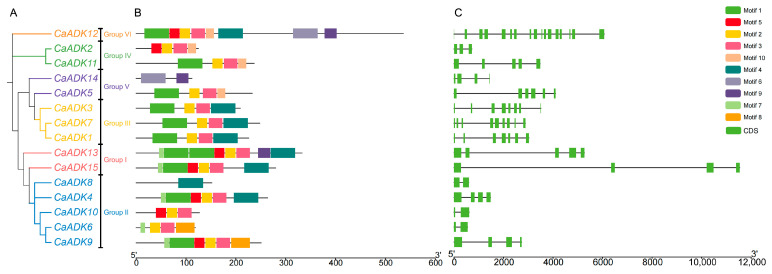 Figure 3
Figure 3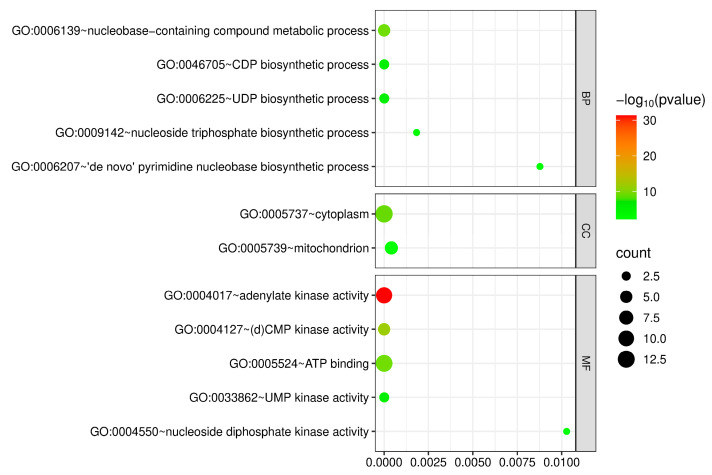 Figure 4
Figure 4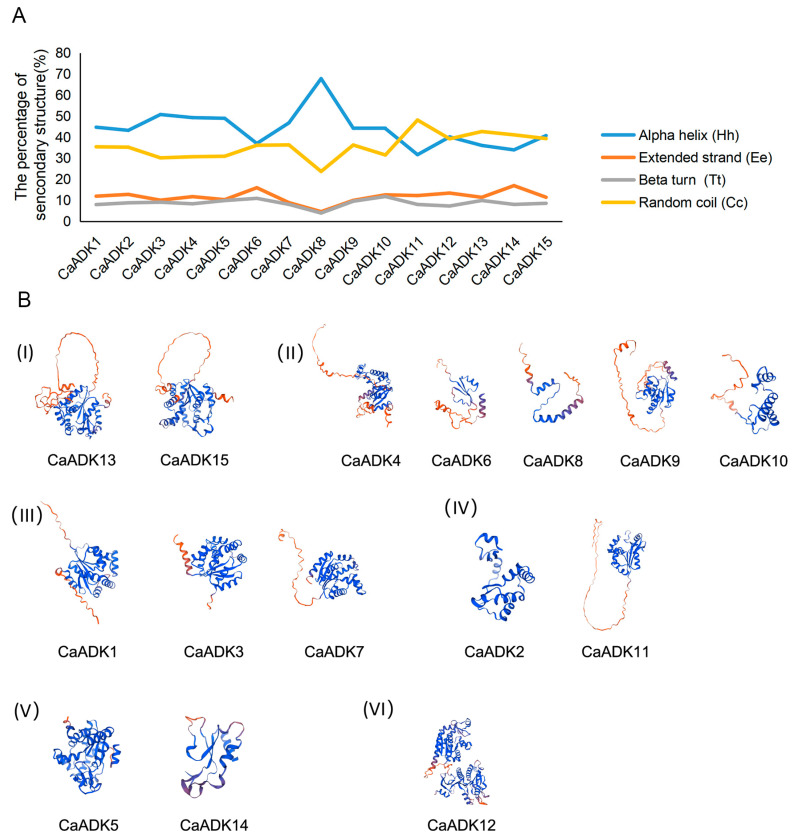 Figure 5
Figure 5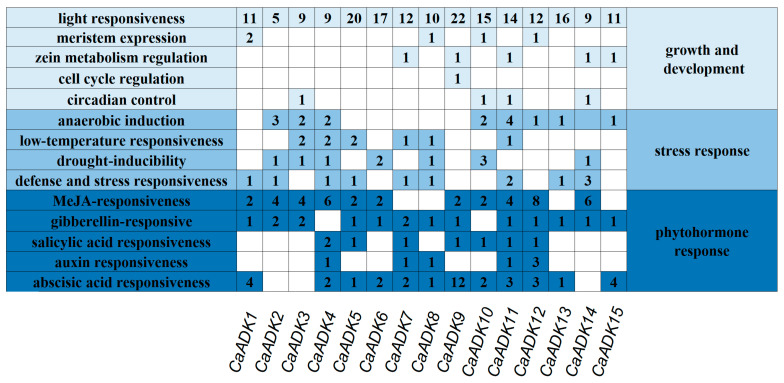 Figure 6
Figure 6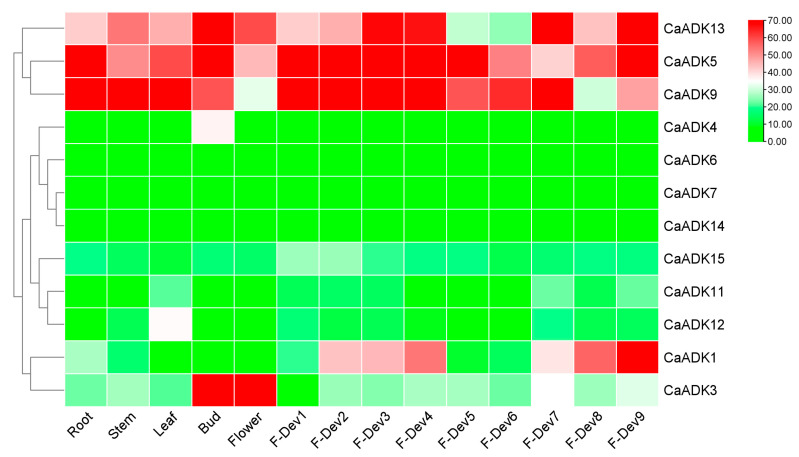 Figure 7
Figure 7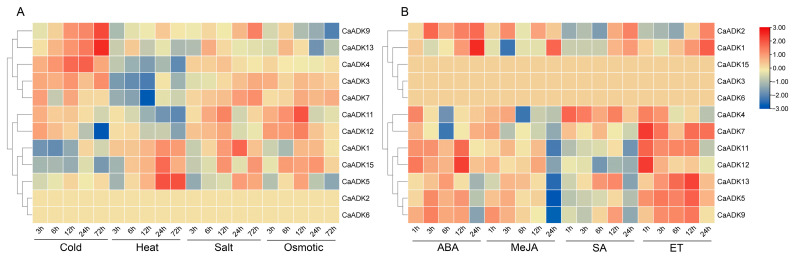 Figure 8
Figure 8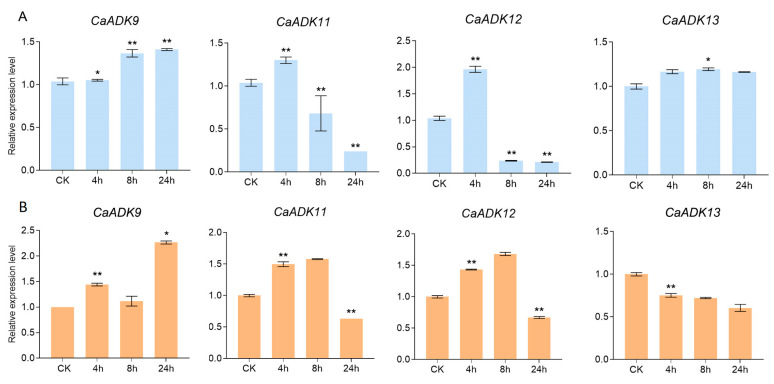 Figure 9
Figure 9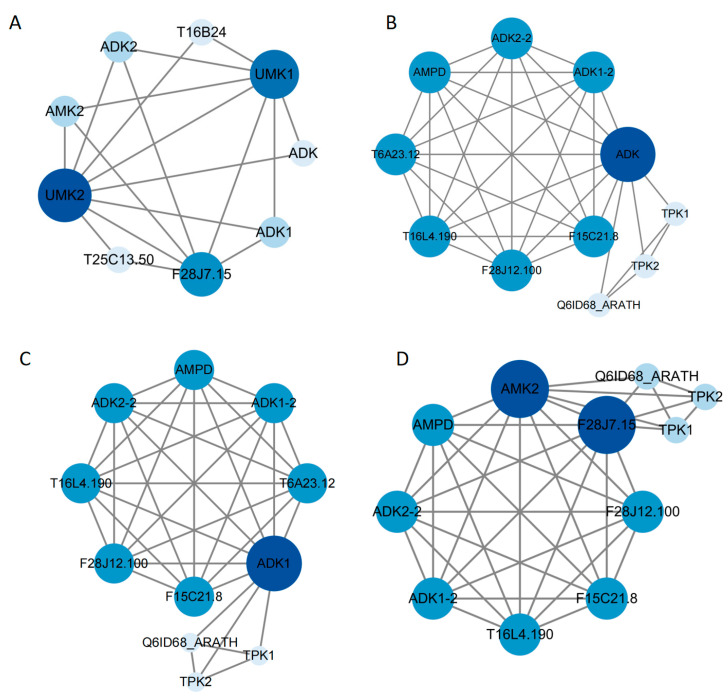 Figure 10
Figure 10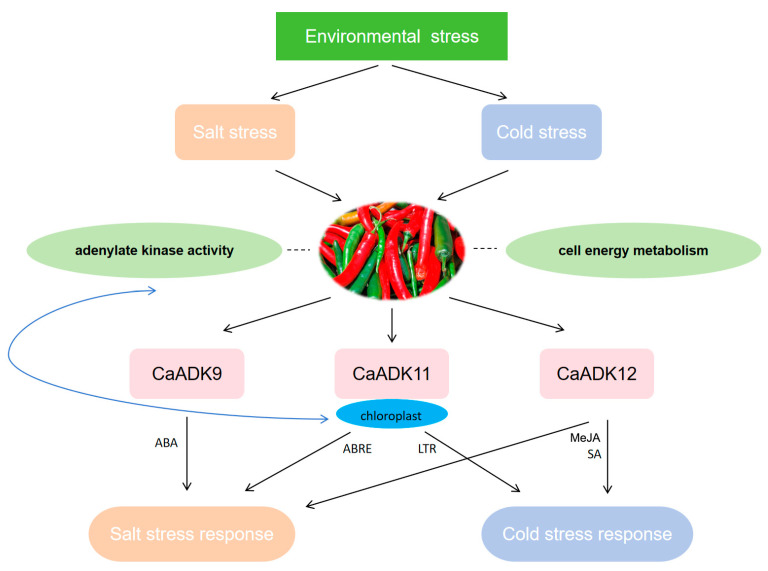 Figure 11
Figure 11- —Postdoctoral Research Startup Fund
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant-Microbe Interactions and Immunity · Piperaceae Chemical and Biological Studies · Plant tissue culture and regeneration
1. Introduction
Adenylate metabolism serves as a fundamental pillar of primary metabolism in living organisms. At its core lies the adenylate cycle, where the interconversion of ATP, ADP, and AMP acts as the “energy currency” driving cellular proliferation and developmental transitions in plants [1]. Adenylate kinase (ADK), a ubiquitously distributed and evolutionarily conserved enzyme, plays a pivotal role in this cycle. It catalyzes the reversible reaction of phosphate transfer between ATP and AMP (ATP + AMP ⇋ 2ADP), a process critical for maintaining adenylate homeostasis within cells [2,3]. Early investigations revealed that ADK also exerts control over the efficiency of energy transfer from mitochondria to hexokinase, laying essential groundwork for understanding its involvement in cellular energy balance regulation [4]. Given the centrality of energy homeostasis in enabling plants to cope with abiotic stresses, ADK is hypothesized to hold vital importance in mediating the stress response of pepper [4].
The “energy charge ratio”, determined by the relative concentrations of AMP, ADP, and ATP, stands as a key regulator of carbohydrate metabolism. This ratio further exerts a profound influence on plant growth, development, and their ability to adapt to external environmental challenges [1,5,6]. Structurally, ADK proteins typically feature three conserved domains: an AMP-binding domain, an ATP-binding domain, and a large central CORE domain [7,8].
Extensive functional studies conducted on model plants and major crops have underscored the biological significance of ADK. It is highly conserved across organisms, and its activity has been functionally characterized in model plants (Arabidopsis thaliana, Oryza sativa) with subcellular localization observed in the cytoplasm, mitochondria, nucleus, and plastids [9,10,11]. In Arabidopsis thaliana, disruption of the ADK gene At5g47840 leads to a loss of chloroplast integrity, manifesting as a bleached phenotype during the critical transition from early embryo to seedling development [5]. Another study revealed that disruption of the ADK gene At2g37250 via T-DNA insertion exhibited elevated amino acid accumulation and promoted root elongation [12]. In Solanum tuberosum (potato), the downregulation of StADK expression results in alterations to adenylate levels and starch content [6]. Beyond regulating developmental processes, ADK orchestrates abiotic stress adaptation through dynamic adjustments of adenylate pools, which directly influence stress-responsive gene expression [13]. In Glycine max, the expression of ADK genes was higher in salt-tolerant varieties than in salt-sensitive ones after salt stress, suggesting that GmADK is involved in the salt response [14]. In Solanum lycopersicum (tomato), microarray analysis revealed that the ADK homolog (SGN-U214214) was downregulated under salt stress, while the ADK gene (SGN-U232826) in drought-tolerant species showed significant upregulation under drought conditions [15,16]. Collectively, these studies highlight the conserved yet diverse roles of ADK in plant growth, development, and stress resistance, providing a compelling rationale for exploring its function in other crop species, including pepper.
Pepper (Capsicum annuum L.), a globally significant vegetable and spice crop, plays a pivotal role in agricultural production, particularly in China. It is rich in capsaicin, carotenoids, and vitamin C and is widely used in food and pharmaceutical industries [17,18]. However, abiotic stresses such as salt and low-temperature stress pose severe threats to pepper metabolism, ultimately leading to reduced yield and quality [19]. Despite the well-documented roles of ADK in stress adaptation across other plant species, the molecular characteristics and stress-responsive functions of ADK genes in pepper remain largely uncharacterized. This knowledge gap impedes our understanding of how pepper regulates energy homeostasis under stress and restricts the development of stress-tolerant pepper varieties to some extent.
To address these critical knowledge gaps, this study aims to systematically identify and characterize the ADK gene family in pepper. We hypothesize that CaADK genes in pepper have undergone lineage-specific diversification, with distinct members integrating energy metabolism and hormone-mediated signaling to regulate adaptations to salt, cold, and combined stresses. Their functional divergence is shaped by cis-regulatory elements and phylogenetic relationships within Solanaceae. Specifically, we will identify pepper ADK genes, analyze their evolutionary relationships, structural properties, and tissue-specific expression patterns, and investigate their transcriptional dynamics under low-temperature and salt stress to unravel their roles in abiotic stress adaptation. By achieving these aims, we hope to deliver the first full-scale analysis of the CaADK family in pepper. This work will lay a solid foundation for understanding ADK-mediated stress adaptation in Solanaceous crops, facilitate the development of stress-resistant pepper varieties, and deepen insights into plant stress physiology.
2. Results
2.1. Genome-Wide Identification and Characterization of CaADK Family Members
Adenylate kinases (ADKs) are evolutionarily conserved enzymes critical for abiotic stress adaptation, including cold, salinity, and drought responses. To identify potential ADK homologs in Capsicum annuum, we performed a BLAST search against the pepper genome using ADK protein sequences from Arabidopsis thaliana as queries, followed by validation of the conserved ADK domain (PF00406). A total of 15 ADK genes (annotated as CaADK) were identified throughout the Capsicum annuum (pepper) genome. These genes were distributed on eight chromosomes and named CaADK1 to CaADK15 based on their chromosomal positions (Table 1).
To comprehensively understand the basic characteristics of CaADK gene family members, we analyzed their genomic locations, the size of their proteins, instability index (II), grand average of hydropathicity (GRAVY), molecular weights (MW), and isoelectric points (pI). The protein lengths of CaADKs ranged from 112 amino acids (CaADK14) to 536 amino acids (CaADK12), the GRAVY values ranged from −0.466 (CaADK7) to 0.302 (CaADK14), and molecular masses ranged from 12,137.1 Da (CaADK14) to 59,842.7 Da (CaADK12). The pI spanned from 4.67 (CaADK14) to 8.13 (CaADK13). Subcellular localization predictions revealed compartment-specific distributions: six cytoplasmic-targeted, three chloroplast, two mitochondrial, three nuclear, and one cytoskeletal. These findings highlight structural and functional diversification within the CaADK family.
2.2. Phylogenetic Analysis and Classification of CaADKs
To investigate evolutionary relationships within the ADK family, we mapped the chromosomal locations of CaADK genes and identified 15 members localized across nine chromosomes (Figure 1A). Chromosomes 01 and 03 had the greatest number of predicted CaADKs, with three genes each, and no CaADK existed on chromosomes 07, 08, and 10. Two genes were present on chromosomes 04 and 09, respectively, and only one CaADK was found on each of the other five chromosomes.
A maximum-likelihood phylogenetic tree was constructed using full-length ADK protein sequences from Capsicum annuum (15), Arabidopsis thaliana (7), Oryza sativa (7), Solanum lycopersicum (11), and Solanum tuberosum (12). The phylogenetic tree resolved ADKs into six distinct clades (Group I–Group VI), with CaADK genes represented in all clades (Figure 1B). Group II was the largest, containing 12 ADKs: 5 from pepper, 1 from Arabidopsis, 2 from rice, and 2 each from tomato and potato, suggesting conserved roles in basal metabolism. Clade V, the second-largest clade, included 10 ADKs: 2 of each of the five species. Clades I and III each contained 9 ADKs, Group IV had 7 members, and Group VI had the fewest, with only 5 members from pepper, Arabidopsis thaliana, rice, potato, and tomato. These phylogenetic relationships indicate that CaADKs share evolutionary conservation with orthologs in other plants, particularly Solanaceous species, while also exhibiting lineage-specific diversification.
Gene duplication, including tandem and segmental duplication events, serves as a key mechanism driving functional innovation and adaptive evolution in plants. Our analysis revealed two segmental duplication events in the pepper genome: CaADK1/CaADK3 and CaADK4/CaADK9 (Figure 1A). Additionally, homology analyses were performed among pepper, Arabidopsis, rice, tomato, and potato ADK genes. These analysis results identified 11 orthologous ADK pairs between pepper and potato (Figure 2D), 11 with tomato (Figure 2C), and 5 with Arabidopsis (Figure 2A), while no orthologs were detected in rice, reflecting monocot-dicot divergence (Figure 2B). Studies on the evolution of Solanaceous plants have revealed their genome evolution and polyploidization events, which may be important forces shaping the evolution of the CaADK gene family [20].
2.3. Diversity Among CaADK Family Members
To unravel the structural divergence of CaADK family members, we systematically analyzed their genetic architectures. A maximum likelihood phylogenetic tree constructed with MEGA X is shown in Figure 3A, which is consistent with the result in Figure 1B. To further explore conservation patterns within the family, we analyzed conserved motifs and exon-intron organizations in the context of this phylogenetic tree. Ten motifs were identified in CaADK proteins and annotated as Motifs 1 to 10 (Figure 3B and Table 2). Conserved motif analysis of CaADK proteins revealed that Motif 2 and Motif 3 were ubiquitously conserved, present in 13 of the 15 genes. Motif 1 was also highly conserved, existing in 10 genes (67%). In contrast, Motif 6 was restricted to CaADK12 and CaADK14, while Motif 8 was lineage-specific—found only in CaADK6 and CaADK9 of Group II—suggesting it may contribute to clade-specific regulatory functions. Notably, within Group II, CaADK8 contained 1 motif, differing from other members in the clade; such divergence may result from functional specialization or genome annotation inaccuracies, potentially leading to distinct expression profiles and functional modifications among family members.
Beyond motif conservation, structural divergence was evident in exon-intron organization. The number of exons in the CaADK family ranged from 2 to 17: CaADK12 had the maximum number of exons, while CaADK8, CaADK10, and CaADK6 (all in Group II) had the shortest gene length with 2 exons. We observed that exon lengths within each clade were highly conserved, whereas introns exhibited substantial size variation, reflecting selection for conserved coding sequences and flexible regulatory evolution through intronic plasticity (Figure 3C). For example, in Group I, CaADK13 and CaADK15 had long gene lengths due to considerably extended introns, despite containing only 5 and 4 exons, respectively, which may enable fine-tuning of their expression through intronic regulatory elements.
2.4. GO Analysis of ADKs in Pepper
Gene ontology (GO) enrichment analysis of CaADKs was conducted, categorizing terms into biological process (BP), cellular component (CC), and molecular function (MF) (Figure 4). In the BP category, CaADKs were enriched in nucleotide metabolism-related processes, including nucleobase-containing compound metabolic process (GO:0006139), CDP biosynthetic process (GO:0046705), UDP biosynthetic process (GO:0006225), de novo’ pyrimidine nucleobase biosynthetic process (GO:0006207), and nucleoside triphosphate biosynthetic process (GO:0009142). For CC terms, the most prominent categories were cytoplasm (GO:0005737) and mitochondrion (GO:0005739), which were consistent with the subcellular localization prediction results. The MF category exhibited the strongest enrichment, with CaADKs significantly associated with adenylate kinase activity (GO:0004017). Collectively, these results indicate that CaADKs primarily function in nucleotide metabolism and adenylate kinase activity, with their subcellular localization supporting roles in cellular energy homeostasis.
2.5. Protein Secondary and Tertiary Structure Analysis of CaADKs
To explore structural conservation and diversification of ADK proteins, we performed comparative analysis focusing on pepper CaADKs, which helps elucidate conserved catalytic domains and lineage-specific adaptations underlying functional divergence across plant species. Secondary structure analysis of ADK proteins in pepper revealed four elements: alpha helix (Hh), extended strand (Ee), beta turn (Tt), and random coil (Cc) components (Figure 5A). Alpha helixes and random coils dominated CaADK secondary structures, whereas beta turns were minimal, suggesting limited rigid loops. Such variations in secondary structure—particularly in the proportion of alpha helices (critical for catalytic domain stability) and random coils (involved in flexible protein interactions)—may contribute to functional divergence, such as differences in allosteric regulation among stress-responsive isoforms.
Building on the secondary structure analysis, we further predicted the tertiary structures of CaADK proteins through homology modeling, which results from additional coiling and folding of secondary structural elements. SWISS-MODEL analysis revealed that members of the same phylogenetic subgroup shared high similarity in tertiary structure (Figure 5B), aligning with their conserved evolutionary origin and suggesting functional redundancy within subgroups. Visualizing these tertiary conformations provides mechanistic insights into how structural differences may drive functional divergence among distinct CaADK members.
2.6. Cis-Acting Element Analysis of CaADKs
To investigate the regulatory mechanisms of the ADK genes, we analyzed cis-acting elements in the 2-kb promoter regions upstream of CaADK coding sequences using PlantCARE. These elements included those associated with stress responses (e.g., low-temperature responsiveness, drought-inducibility) and phytohormone signaling (e.g., abscisic acid responsiveness, gibberellin responsive), with detailed information provided in Table S5.
We found variable numbers of cis element across CaADK promoters, with stress- and hormone-related elements being most abundant (Figure 6). Specifically, stress-responsive elements included those for anaerobic induction, low-temperature responsiveness, drought-inducibility, and defense/stress responsiveness; phytohormone-responsive elements such as abscisic acid responsiveness, MeJA responsiveness, gibberellin responsive, and salicylic acid responsiveness. Notably, all CaADK promoters contained light-response elements, implicating their potential role in coordinating with light-dependent processes. Approximately 87% (13 of 15) of these genes harbored gibberellin responsive elements, 80% (12 of 15) contained abscisic acid responsiveness elements, and 73% (11 of 15) had MeJA-responsiveness elements. The number and type of cis elements differed among the promoters of each CaADK gene, reflecting their distinct functional requirements. Collectively, these data—particularly the enrichment of stress- and hormone-related elements—support that CaADK genes are integrated into abiotic stress regulatory networks, with their unique cis element profiles enabling specific responses to diverse environmental cues.
2.7. Tissue-Specific Expression Patterns of CaADKs
Tissue-specific expression profiles of 15 CaADKs (analyzed using transcriptome data from ‘Zunla-1’) revealed distinct transcriptional patterns across five vegetative tissues and nine fruit developmental stages (Figure 7, Table S2). Six CaADK genes (CaADK4, CaADK7, CaADK14, CaADK15, CaADK11, and CaADK12) showed minimal expression across all tissues, suggesting they may be functionally specialized or induced only under specific conditions, whereas CaADK6 was silenced in different parts of pepper and fruit growth stages. In contrast, CaADK13, CaADK5, and CaADK9 were highly expressed in multiple tissues, including roots, stems, leaves, and fruits, indicating potential housekeeping roles in basal biological processes. Notably, CaADK3 exhibited preferential expression in floral tissues, implying a role in reproductive development. CaADK1 showed peak expression at 7 days post-color transition in fruits, suggesting involvement in fruit ripening or pigment accumulation. These tissue-specific patterns highlight functional diversification within the CaADK family, with some members contributing to general cellular processes and others to tissue-specific development.
2.8. Transcriptome Analysis of CaADKs in Response to Abiotic Stresses and Exogenous Hormone Treatments
To elucidate the functional roles of the ADK gene family in stress adaptation, we profiled CaADK expression under abiotic stress and exogenous hormone treatments. The expression dynamics of these genes were consistent with the timing of their upregulation or downregulation in the transcriptome (Figure 8).
The expression levels of CaADK genes varied under different treatments (Figure 8A). Several genes exhibited significant and consistent expression trends. Under low-temperature treatment, the expression levels of CaADK9 and CaADK13 were upregulated, with the highest expression observed after 72 h of treatment—this sustained upregulation suggests that these two members may contribute to long-term cold acclimation. CaADK5 expression was upregulated under both heat and salt stresses, while CaADK7 was specifically upregulated under salt stress, indicating potential roles in mediating responses to multiple or specific stressors. In contrast, CaADK11 and CaADK12 were suppressed under both cold and heat stress, implying they may be involved in negative regulation or energy conservation under these conditions. These findings demonstrate that CaADKs are critical regulators of abiotic stress adaptation, particularly in low-temperature and salinity stresses.
Exogenous hormone treatments revealed that most CaADKs experienced transient suppression (1–6 hpt) followed by gradual recovery (6–24 hpt) (Figure 8B), suggesting a conserved early response mechanism to hormonal cues. Conversely, CaADK1 was significantly upregulated at 12 h under salicylate treatment, while CaADK9 was suppressed under MeJA treatment, indicating member-specific interactions with hormone signaling pathways. Notably, CaADK2 and CaADK6 were transcriptionally silent under abiotic stresses, and CaADK3/6/15 remained unresponsive to hormonal treatments, suggesting that they are insensitive to changes in the external environment. These distinct expression patterns under stress and hormone treatments indicate that the CaADK family employs a specialized regulatory network to mediate pepper’s adaptation to environmental challenges, with specific members likely acting as key nodes in stress signaling pathways.
2.9. Expression Patterns of CaADKs Under Abiotic Stress
To validate the expression dynamics of CaADK genes under abiotic stress, four candidate genes (CaADK9, CaADK11, CaADK12, and CaADK13) in pepper—selected based on their differential expression in transcriptome data—were analyzed using RT-qPCR analysis at 0 h, 4 h, 8 h, and 24 h post treatment (hpt) under both low-temperature and salt stress (Figure 9).
Under low-temperature stress, CaADK11 and CaADK12 were significantly downregulated by over 4-fold after 24 h of low-temperature stress (p < 0.01, t-test), consistent with transcriptome results. CaADK9 showed upregulation at 8 hpt, while CaADK13 exhibited a slight upregulation throughout the treatment period (Figure 9A). Under salt stress, CaADK11, CaADK12, and CaADK13 were significantly downregulated at 24 hpt, whereas CaADK9 was markedly upregulated at all time points (Figure 9B). Notably, CaADK11 and CaADK12 showed a transient upregulation at 4 h under salt stress, indicating they may participate in early stress sensing, while CaADK9 maintained elevated expression, suggesting a role in sustained salt stress response. These results confirm that CaADK genes exhibit dynamic and divergent expression patterns in response to low-temperature and salt stress, with functional specialization likely enabling fine-tuning of pepper’s stress adaptation mechanisms.
2.10. PPI Network Prediction Analysis of ADK Genes
Proteins play crucial roles in mediating molecular interactions and cellular signaling pathways. To explore the molecular networks underlying ADK-mediated stress responses, we predicted protein-protein interaction (PPI) networks using Arabidopsis homologs of CaADKs (leveraging Arabidopsis’ well-characterized PPI data) and inferred potential interactions of pepper ADKs based on these orthologous relationships, aiming to identify functional partners in nucleotide metabolism and stress signaling (Figure 10A). The results showed that four Arabidopsis homologs of CaADKs interacted with conserved protein partners: At2g37250 (homologs of CaADK13 and CaADK15, Figure 10B), At5g63400 (homolog of CaADK5, Figure 10C), At5g47840 (homologs of CaADK2 and CaADK11, Figure 10D), and At3g01820 (homologs of CaADK4, CaADK6, CaADK9, and CaADK10; Figure 10D). These homologs interacted with core proteins, including ADK1-1, ADK1-2, AMPD (adenylate deaminase), F15C21.48, F28J12.100, and T16L4.190, with low-degree interactions with TPK1, TPK2, and Q6ID68_ARATH. Notably, interacting proteins such as AMPD (involved in adenylate catabolism) and ADK1 homologs may regulate adenylate kinase activity by modulating nucleotide pool balance. Consistent with this, previous studies reported that ADK1 elevates intracellular ATP levels to fuel glycolysis and oxidative phosphorylation [21], supporting a conserved role of these interactions in energy metabolism. Collectively, these predictions suggest that CaADKs coordinate metabolic adaptation to stress through a conserved network of interactions with nucleotide homeostasis-related proteins, highlighting their role in integrating energy metabolism and stress signaling.
3. Discussion
Adenylate kinase (ADK) is a ubiquitous enzyme that mediates the reaction generating ADP (ATP + AMP ⇋ 2ADP), thereby maintaining cellular energy homeostasis. Beyond its role in energy balance, ADK modulates plant growth, developmental transitions, and adaptation to abiotic stresses [12,22]. However, the ADK gene family in pepper (Capsicum annuum L.) has not been thoroughly studied. In this study, we identified 15 CaADK genes in the pepper genome, a number comparable to ADK families in other crops—13 in rice [23], 11 in tomato [24], and 11 in alfalfa [25]—suggesting conserved expansion patterns of the ADK family across angiosperms. We characterized their physicochemical properties, phylogenetic relationships, structural features, GO enrichment, protein structures, cis elements, protein interactions, and expression patterns under abiotic stresses. These results enhance our understanding of CaADK genes and their roles in plant responses to abiotic stresses.
Phylogenetic analysis classified CaADKs from five plant species into six clades, with genes within the same clade sharing closer evolutionary relationships. This clustering—CaADK13/15 in Clade I, CaADK4/6/8/9/10 in Group II, and so on—revealed conserved functional architectures, as evidenced by clade-specific motif distributions and exon-intron organizations. CaADK1, 3, and 7 in Clade III shared homology with UMP-CMP kinases and exhibited strong sequence homology to their Arabidopsis orthologs. The 10 identified conserved motifs, particularly their consistent arrangement within clades, suggest potential functional redundancy among closely related members. For example, in Clade III, CaADK1, 3, and 7, which share homology with UMP-CMP kinases and exhibit strong sequence homology to their Arabidopsis orthologs, may have overlapping functions in nucleotide metabolism. On the other hand, lineage-specific motifs (e.g., Motif 8 in Group II) may drive functional specialization [26]. These unique motifs could enable interactions with distinct sets of regulatory or substrate proteins, allowing ADKs to participate in novel stress-responsive pathways in pepper. Exon counts ranging from 2 to 17 further highlight the evolutionary dynamism of the family. Conserved exon lengths suggest that the core catalytic domains of ADKs are under strong purifying selection, ensuring the maintenance of their fundamental enzymatic activity. Notably, homology analyses revealed that pepper ADKs share the most orthologous pairs with potato and tomato, underscoring close evolutionary ties among Solanaceous crops [27]. The Solanaceae family has evolved in diverse ecological niches, and the conservation of ADK orthologs among its members suggests that these genes have been co-opted to mediate stress responses relevant to the family’s typical habitats, such as arid or saline environments. As depicted in Figure 2, orthologous genes were not identified in rice. The absence of orthologous pairs with rice aligns with prior observations of limited collinearity between monocots and dicots [28], reinforcing the evolutionary divergence of ADK families between these lineages. Collectively, these results underscore the evolutionary divergence of the ADK gene family between monocotyledonous and dicotyledonous plant species. Therefore, we can predict the gene functions of pepper based on those of other nightshade plants.
Tissue-specific expression profiling demonstrated that CaADK genes exhibited ubiquitous distribution across various tissues, mirroring the tissue-dependent specialization reported for ADKs in other plants [28]. CaADKs were detected in all examined tissues and throughout fruit development stages in pepper. Several CaADK family members, including CaADK5, CaADK9, and CaADK13, were identified as key regulators of pepper growth and development, as evidenced by their widespread expression in both vegetative and reproductive tissues. In plants, energy demands vary across tissues and developmental stages. The high expression of these CaADKs in roots, stems, and leaves aligns with roles in carbohydrate metabolism, a critical process for plant growth. Carbohydrates are not only energy sources but also signaling molecules, and ADKs may play a dual role in regulating both energy availability and carbohydrate-mediated signaling in these tissues.
In this study, we analyzed 14 cis-regulatory elements associated with stress responses and hormone regulation, as detailed in Table S5 and Figure 6. Cis-acting element analysis and stress-responsive expression data collectively illuminate the roles of specific CaADKs in abiotic stress adaptation. CaADK9, whose promoter contains abscisic acid (ABA)-responsive elements, was consistently upregulated under salt stress in both RNA-seq and RT-qPCR analyses. Abscisic acid (ABA) regulates seed germination, development, and stress responses [29]. Previous studies have shown that ABA is a typical plant hormone associated with abiotic stress, promoting the expression of stress-related genes and playing an important role in responses to external stress, such as salt stress [30,31]. In addition, expression analysis could provide insight into gene functions. ADK genes responded rapidly to biotic and abiotic stresses such as salt stress [32] and also affect ATP metabolism and adenylate balance. The upregulation of CaADK9 under salt stress suggests that it may act through ABA-dependent pathways. One possible mechanism is that CaADK9 regulates ATP regeneration to fuel stress-responsive processes such as ion transport. Under salt stress, plants need to maintain ion homeostasis, which requires energy for the operation of ion transporters.
CaADK12 emerges as a critical regulator of combined salt and cold stress tolerance. Analysis of cis-acting elements within the promoter region of CaADK12 revealed the presence of MeJA, GA, and SA responsive elements, positioning it at the intersection of multiple hormone-mediated stress response pathways. Gibberellin (GA) regulates stress responses by preventing protein degradation through the activation of DELLA protein, which is a repressor on the GA signaling pathway [33]; salicylic acid in plants mediates cross-talk between stress-signaling pathways and enhances cold tolerance [34,35]; and MeJA drives cold-stress responses in tomato by upregulating abscisic acid (ABA) biosynthesis [36]. Exogenous MeJA mediates the expression of genes across diverse pathways. For example, it impacts processes such as chlorophyll biosynthesis and degradation, antioxidant enzyme systems, and jasmonic acid (JA) biosynthesis, thereby enhancing the plant’s tolerance to abiotic stresses [37]. CaADK12 was found to possess a substantial number of MeJA-responsive cis elements. This discovery strongly implies its potential involvement in the regulatory mechanisms of the MeJA response to stress. Additionally, results from exogenous hormone treatments indicated that the expression level of CaADK12 was significantly downregulated after MeJA and SA treatments. Transcriptomic and RT-qPCR data confirm downregulation of CaADK12 under both salt and cold stress, with prolonged suppression suggesting an energy-conserving strategy to prioritize critical stress-acclimation processes. When plants are exposed to multiple stresses, energy becomes a scarce resource, and downregulating certain ADKs may redirect energy to more essential processes such as the synthesis of stress-protective proteins or osmolytes. The role of CaADK12 in this energy reallocation under combined stress conditions highlights the complexity of ADK-mediated stress responses.
Structural analyses further support functional specialization. Secondary structures dominated by alpha helices and random coils provide flexibility for allosteric regulation, which is crucial for enzymes such as ADKs, as it allows them to respond to changes in the cellular environment. While conserved tertiary structures within clades ensure core catalytic functionality, allowing ADKs to perform their basic role in nucleotide phosphorylation-dephosphorylation reactions [21,38]. Subcellular localization predictions indicated that CaADK proteins are localized to mitochondria and chloroplasts. Consistent with our findings, ADKs in plants are predominantly localized in the chloroplast stroma and mitochondrial intermembrane space, where they account for 90% of total enzymatic activity [38,39]. These features, combined with protein-protein interaction predictions linking CaADKs to nucleotide homeostasis proteins (AMPD, ADK1 homologs), reinforce their role as integrators of energy metabolism and stress signaling. For instance, interactions with AMPD (adenosine monophosphate deaminase) could allow CaADKs to sense and respond to changes in AMP levels, which are key indicators of cellular energy status. In times of stress, when AMP levels rise, this interaction could trigger a cascade of events to restore energy homeostasis, either by activating ADK to convert AMP and ATP to ADP or by initiating other metabolic or signaling pathways.
Previous studies demonstrate that the ADK gene At5g47840 is vital for chloroplast integrity and ensures plants grow properly from embryo to seedling development [5]. CaADK11, a chloroplast-localized homolog of Arabidopsis At5g47840, contains abundant anaerobic induction and low-temperature responsive elements. Both transcriptome and RT-qPCR data collectively confirmed that the expression of CaADK11 was downregulated under salt and cold stress conditions, implying its role in coordinating chloroplast function under stress. By reducing non-essential metabolic activity in the chloroplast, CaADK11 may preserve resources to maintain basic chloroplast functions, such as the synthesis of essential proteins and lipids. This aligns with subcellular localization predictions, as chloroplast and mitochondrial ADKs are known to regulate energy balance in organelles critical for stress adaptation. Based on these findings, we propose that the regulation of CaADK11 in response to stress thus represents a fine-tuned mechanism to balance energy consumption and organelle function under adverse conditions.
In summary, our findings reveal that the CaADK family has evolved conserved and lineage-specific features, with distinct members mediating pepper’s response to cold and salt stress through differential expression, structural specialization, and interactions with nucleotide metabolism proteins. These provide a foundational characterization of CaADKs.
4. Materials and Methods
4.1. Identification of ADK Genes in the Pepper Genome and Subcellular Localization Analysis
The genome of ‘Zunla-1’, sequenced and assembled in China, served as the primary reference genome for this study (accession no. PRJcA025503). Genome sequences of Oryza sativa (rice), Solanum lycopersicum (tomato), and Solanum tuberosum (potato) were retrieved from the NCBI database (https://www.ncbi.nlm.nih.gov/, accessed on 2 February 2025) for comparative analysis. To annotate CaADK genes in the pepper genome, the full protein sequences of ADKs in Arabidopsis were obtained from the TAIR database (https://www.arabidopsis.org/, accessed on 2 February 2025). Sequences of Arabidopsis served as queries for a BLAST searches (E-value < 1 × 10^−10^) against the pepper genome. Pfam domain databases (https://www.ebi.ac.uk/interpro/, accessed on 13 February 2025) were employed to construct a genome-wide protein domain model for pepper [40,41].
The physicochemical properties of pepper ADK gene family members were predicted using the ExPASy technologies server (http://web.expasy.org/, accessed on 23 February 2025), and the subcellular distribution was analyzed using WoLF PSORT (http://wolfpsort.org/, accessed on 23 February 2025) [40,42].
4.2. Chromosomal Localization, Synteny, and Phylogenetic Analysis
The chromosomal locations of CaADK family genes were mapped using TBtools, and intra-genomic synteny analysis was conducted with MCScanX [43,44]. Amino acid (aa) sequences of CaADKs and ADK genes from other species were aligned via the MUSCLE algorithm. Phylogenetic inference was performed in MEGA X, with all major clades having bootstrap support > 95% (1000 replicates) [45]. Phylogenetic trees were visualized and annotated using iTOL (https://itol.embl.de/, accessed on 24 February 2025) [46].
4.3. Motif, Gene Structure, and Conserved Domain Analysis
Coding sequence (CDS) and gff3 format files of Capsicum annuum were downloaded from the pepper genome database (http://www.bioinformaticslab.cn/PepperBase/, accessed on 24 February 2025) [47]. The intron-exon architectures were visualized using TBtools software. Conserved protein motifs were identified using the online software MEME suite version 5.5.8 [48]. In MEME (Multiple Expectation Maximization for Motif Elicitation) analysis, the maximum number of motifs was set to 10, and the occurrences of a single motif were set to zero or one per sequence. These were visualized using “Visualize Motif Pattern” (from the MEME suite) in TBtools.
4.4. Protein Secondary and Tertiary Structure Analysis
PRABI was used to analyze the secondary structure of CaADK proteins (https://prabi.ibcp.fr/htm/site/web/app.php/home, accessed on 24 February 2025), and SWISS-MODEL (https://swissmodel.expasy.org/, accessed on 25 February 2025) was employed for the protein tertiary structure analysis [49].
4.5. Cis-Acting Element Analysis and Gene Ontology Analysis
The 2-kb sequences upstream of the CaADK family genes were selected for cis-acting element analysis using the online tool PlantCARE with default parameters [50]. A gene ontology (GO) analysis of the CaADK genes was conducted with default parameters at the DAVID website (http://david.ncifcrf.gov, accessed on 26 February 2025) [51].
4.6. Tissue-Specific Expression
To profile tissue-specific expression patterns of CaADK genes, the transcriptome annotation files of roots, stems, leaves, buds, flowers, and fruit development stages of pepper ‘Zunla-1’ were obtained through the GEO database (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc%20=%20GSE45037, accessed on 24 February 2025). Tissue-specific expression data were analyzed using TBtools (v1.120).
4.7. Transcriptome Data Analysis of ADK Gene Family in Pepper
Publicly available transcriptome data for ‘Zunla-1’ was downloaded from the NCBI GEO database (accession numbers GSE132824 and GSE149037). Gene expression data for CaADK genes was collected in leaves from 0 to 24 h (sampled at 1, 3, 6, 12, and 24 h) after different abiotic stresses, such as cold stress (10 °C), heat stress (40 °C), salt stress (400 mM NaCl), and osmotic stress (400 mM mannitol), as well as different phytohormone treatments, including SA (5 mM sodium salicylate) and ET (5 mM ethephone). The gene expression level of CaADKs was calculated by the log_2_ FPKM algorithm, and expression data were analyzed using TBtools.
4.8. RNA Extraction, cDNA Synthesis, and RT-qPCR
The test materials (‘Hangjiao-1’ pepper) were exposed to cold stress (4 °C for 0, 4, 8, and 24 h) and salt stress (150 mM NaCl for 0, 4, 8, and 24 h) at the College of Life Sciences, South China Agricultural University (Guangzhou, Guangdong, China). Control (CK) seedlings were maintained under normal conditions. Following stress treatment, the materials were rapidly frozen in liquid nitrogen for further experiments. RNA integrity was evaluated by 1% agarose gel electrophoresis, and the RNA concentration and purity were measured using a spectrophotometer Thermo NanoDrop One (Thermo Fisher Scientific, Waltham, MA, USA).
cDNA was synthesized by reverse transcription using HiScript^®^ II Q RT SuperMix with a qPCR sample kit (Vazyme Biotech Co., Ltd., Nanjing, China), then diluted ten-fold and stored at −20 °C. RT-qPCR was conducted in 20 µL volume containing 2 × SYBR qPCR Master Mix (10.0 μL), upstream and downstream primers (10 μmol∙L^−1^, 0.4 μL each), cDNA template (1.0 μL), and ddH2O (8.2 μL). Thermal cycling conditions included initial denaturation at 95 °C for 30 s; 40 cycles of 95 °C for 10 s (denaturation) and 60 °C for 22 s (annealing/extension); followed by melt curve analysis at 95 °C for 25 s, 60 °C for 60 s, and 95 °C for 7 s. Actin was used as the internal control. The 2^−∆∆CT^ method was used to analyze relative expression levels [52].
4.9. PPI Network Prediction Analysis of ADK Genes
The STRING online website (https://cn.string-db.org/, accessed on 27 February 2025) was used to predict the PPI relationships with default parameters [53], and Cytoscape v3.9.1 was used to construct the interaction network [54].
5. Conclusions
In this study, we systematically identified 15 adenylate kinase genes (CaADKs) in the pepper (Capsicum annuum L.) genome, representing the first comprehensive characterization of the ADK family in this crop. Our analysis enriches the current understanding of ADK gene family evolution and function in Solanaceous plants, complementing existing studies in tomato and potato. Phylogenetic and collinearity analyses revealed that pepper ADKs share a significantly higher level of homology with other Solanaceous crops, including tomato and potato, providing evidence supporting monocot-dicot divergence in the ADK gene family. Through integrated approaches, including subcellular localization prediction, cis element profiling, stress-responsive expression dynamics, and RT-qPCR validation, we identified distinct roles of specific members: CaADK9, characterized by upregulation under salt stress and presence of abscisic acid-responsive promoter elements, mediates salt stress adaptation; CaADK11 and CaADK12, which exhibit significant downregulation under both low-temperature and salt stress, act as critical regulators of combined stress tolerance (schematically summarized in Figure 11).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Fujisawa K. Regulation of Adenine Nucleotide Metabolism by Adenylate Kinase Isozymes: Physiological Roles and Diseases Int. J. Mol. Sci.202324556110.3390/ijms 2406556136982634 PMC 10056885 · doi ↗ · pubmed ↗
- 2Pradet A. Raymond P. Adenine nucleotide Ratios and Adenylate Energy Charge in Energy Metabolism Annu. Rev. Plant Physiol.19833419922410.1146/annurev.pp.34.060183.001215 · doi ↗
- 3Park J. Gupta R.S. Adenosine Meta Bolism, Adenosine Kinase, and Evolution Springer New York, NY, USA 201210.1007/978-1-4614-3903-5_2 · doi ↗
- 4Dzheia P. Kal’venas A. Toleĭkis A. Prashkiavichius A. The role of adenylate kinase in the regulation of the rate and effectiveness of energy transfer from mitochondria to hexokinase in vitro Biokhimiia 1986519749793015265 · pubmed ↗
- 5Lange P.R. Geserick C. Tischendorf G. Zrenner R. Functions of chloroplastic adenylate kinases in Arabidopsis Plant Physiol.200814649250410.1104/pp.107.11470218162585 PMC 2245825 · doi ↗ · pubmed ↗
- 6Regierer B. Fernie A.R. Springer F. Perez-Melis A. Leisse A. Koehl K. Willmitzer L. Geigenberger P. Kossmann J. Starch content and yield increase as a result of altering adenylate pools in transgenic plants Nat. Biotechnol.2002201256126010.1038/nbt 76012426579 · doi ↗ · pubmed ↗
- 7Arora K. Brooks C.L. Large-scale allosteric conformational transitions of adenylate kinase appear to involve a population-shift mechanism Proc. Natl. Acad. Sci. USA 2007104184961850110.1073/pnas.070644310418000050 PMC 2141805 · doi ↗ · pubmed ↗
- 8Maragakis P. Karplus M. Large amplitude conformational change in proteins explored with a plastic network model: Adenylate kinase J. Mol. Biol.200535280782210.1016/j.jmb.2005.07.03116139299 · doi ↗ · pubmed ↗