Genetic Dissection of Plant Height Variation Between the Parental Lines of the Elite Japonica Hybrid Rice ‘Shenyou 26’
Bin Sun, Xiaorui Ding, Kaizhen Xie, Xueqing Zhang, Can Cheng, Yuting Dai, Anpeng Zhang, Jihua Zhou, Fuan Niu, Rongjian Tu, Yue Qiu, Zhizun Feng, Bilian Hu, Chenbing Shao, Hongyu Li, Tianxing Shen, Liming Cao, Huangwei Chu

TL;DR
This study identifies genetic factors influencing plant height in a high-yield rice hybrid, offering insights for improving rice breeding.
Contribution
The study identifies and validates multiple QTLs, including SD1 allelic variants, controlling plant height in the rice hybrid ‘Shenyou 26’.
Findings
Seven QTLs associated with plant height were identified in the DH population of ‘Shenyou 26’.
qPH1.1 co-localizes with SD1, and a rare SD1-EQH allele in ‘Shen 9A’ reduces plant height.
qPH9.1 significantly contributes to plant height variation, with the Shenhui26 allele increasing height.
Abstract
Plant height is a key agronomic trait influencing both seed production and yield in hybrid rice. In the elite japonica hybrid ‘Shenyou 26’, optimal plant height differences between the restorer line (‘Shenhui 26’) and the male sterile line (‘Shen 9A’) are critical for efficient pollination. In this study, we dissected the genetic basis of plant height variation using a doubled haploid (DH) population derived from ‘Shenyou 26’. Multi-environment phenotyping and QTL mapping identified seven QTLs associated with plant height, among which qPH1.1 and qPH9.1 were validated. qPH1.1 co-localized with the semi-dwarf gene SD1, and ‘Shen 9A’ carries a rare SD1-EQH allele that potentially confers reduced height relative to the SD1-EQ allele in ‘Shenhui 26’. qPH9.1 also contributed significantly to plant height variation, with the Shenhui26 allele increasing plant height in backcross validation.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
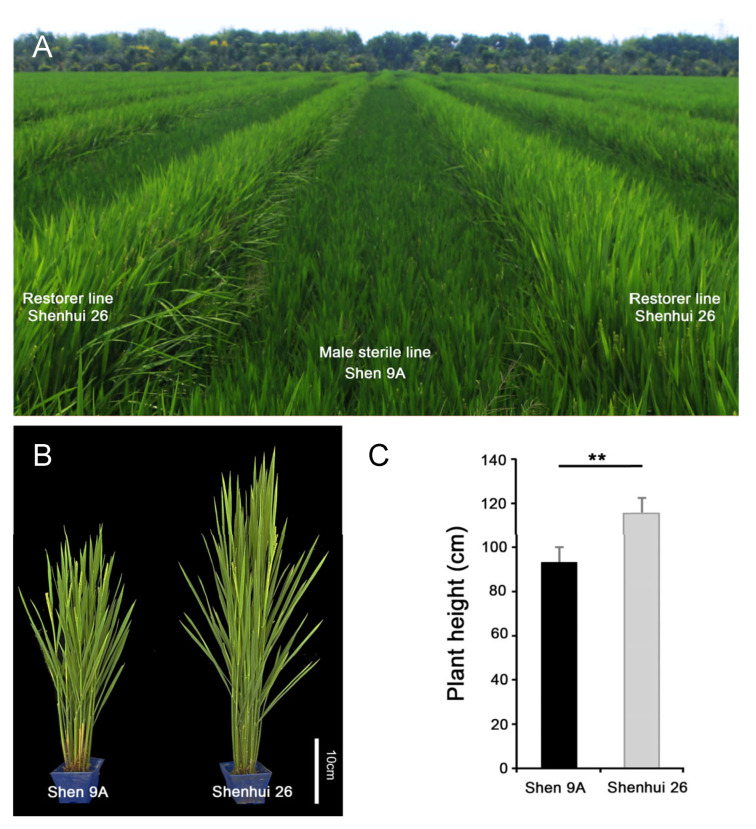 Figure 1
Figure 1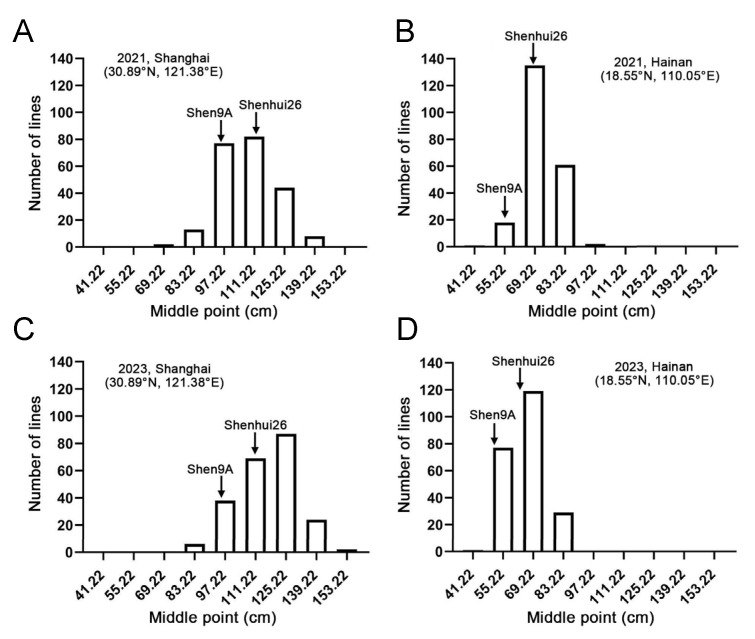 Figure 2
Figure 2 Figure 3
Figure 3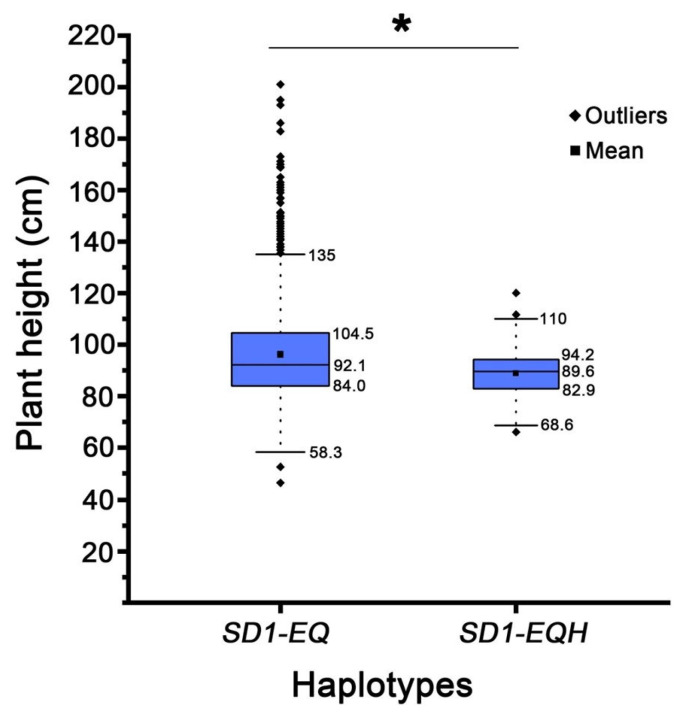 Figure 4
Figure 4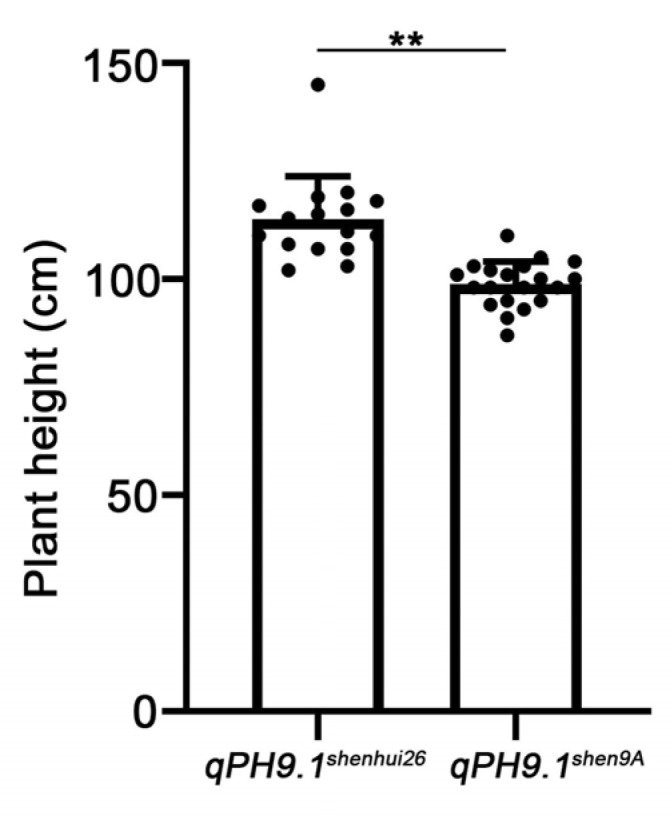 Figure 5
Figure 5- —the Shanghai Science and Technology Innovation Action Plan Project
- —the Shanghai Key Laboratory of Agricultural Genetics and Breeding
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRice Cultivation and Yield Improvement · GABA and Rice Research · Agriculture, Soil, Plant Science
1. Introduction
Plant height is a pivotal agronomic trait that strongly influences rice yield. Excessive plant height increases the risk of lodging and subsequent yield losses, whereas overly short stature can restrict yield potential by limiting biomass accumulation. Thus, optimizing plant height to balance lodging resistance with sufficient biomass production remains a central goal in modern rice breeding [1].
Since the 1960s, the introduction of the semi-dwarf 1 (sd1) gene from the rice variety Dee-Geo-Woo-Gen (DGWG) has dramatically increased rice yields, establishing a cornerstone of the “Green Revolution” in rice breeding [2]. By reducing plant height, sd1 enhanced the harvest index and resolved the trade-off between high yield potential and lodging resistance under high nitrogen fertilizer input [3,4]. Notably, the SD1 locus had already undergone artificial selection prior to the “Green Revolution”. During japonica domestication, two functional nucleotide polymorphisms (FNPs) arose in the japonica SD1 allele (SD1-EQ), reducing GA20ox2 enzyme activity, lowering endogenous gibberellin (GA) levels, and thereby decreasing plant stature. In contrast, most indica landraces and the wild progenitor O. rufipogon retain the strongly functional SD1 allele (SD1-GR), which confers higher GA content and taller growth [5,6,7]. At present, SD1 remains one of the most widely exploited genes in rice improvement, largely due to its abundant natural polymorphism [8,9,10].
However, the widespread deployment of SD1 mutant alleles in rice has also generated unintended consequences. For instance, sustaining high yields in these varieties often requires intensive nitrogen fertilization, which contributes to eutrophication of aquatic ecosystems [11,12]. Moreover, the overreliance on the SD1 mutant alleles not only narrows genetic diversity but also introduces unfavorable traits, including reduced spikelet fertility under low-temperature conditions and heightened sensitivity to drought stress [13,14,15]. To mitigate the drawbacks of excessive reliance on sd1, diversifying the genetic resources for semi-dwarfism through the identification and use of novel, practically valuable genes is essential for sustaining and strengthening the genetic resilience of future rice breeding programs [16,17].
Rice plant height is a complex trait governed by both qualitative and quantitative genes. To date, more than 1000 QTLs associated with plant height have been mapped, and dozens of genes related to dwarfism or semi-dwarfism genes have also been identified from mutants [4,18,19]. However, most of these genes are unsuitable for breeding because they confer undesirable agronomic traits such as excessive dwarfism, reduced fertility, or short grains. Therefore, identifying novel favorable alleles that achieve an optimal balance between plant height and yield remains essential for future rice improvement [20].
In this study, a doubled haploid (DH) population derived from the elite japonica hybrid rice ‘Shenyou 26’ was used to identify QTLs associated with plant height. A total of seven QTLs were detected, including qPH1.1, which may be associated with the SD1 locus, while the genetic effect of qPH9.1 was further validated. These findings contribute to elucidating the genes underlying plant height regulation and provide valuable genetic resources for fine-tuning plant height in rice breeding.
2. Results
2.1. Height Variation Between the Male-Sterile (Shen 9A) and Restorer (Shenhui 26) Lines in the ‘Shenyou 26’ Hybrid System
The three-line japonica hybrid rice ‘Shenyou 26’ was developed by the Shanghai Academy of Agricultural Sciences (SAAS) through using ‘Shen 9A’, a BT-type cytoplasmic male sterile (CMS) line, as the female parent, and the restorer line ‘Shenhui 26’ as the pollen donor [21]. At the heading stage, ‘Shenhui 26’ exhibits slightly taller than ‘Shen 9A’ (Figure 1A–C), ensuring optimal pollination efficiency—a critical factor for high hybrid seed yield. However, the genetic basis underlying this plant height difference remains unclear.
2.2. Genetic Basis of Plant Height Differentiation Between ‘Shen 9A’ and ‘Shenhui 26’
Previously, a doubled haploid (DH) population comprising 232 lines was constructed using young panicles of ‘Shenyou 26’ through anther culture [22]. To investigate the genetic basis of plant height differentiation between ‘Shen 9A’ and ‘Shenhui 26’, plant height was measured and analyzed for the DH population and its parental lines when grown in Shanghai (30.89° N, 121.38° E) and Hainan (18.55° N, 110.05° E) during the 2021 and 2023 growing seasons. Under all four environmental conditions, plant height in the DH population exhibited a continuous variation that approximated a normal distribution. Furthermore, transgressive segregation in both directions was observed (Table 1; Figure 2A–D), indicating that the plant height differentiation between ‘Shen 9A’ and ‘Shenhui 26’ is controlled by multiple quantitative trait loci. Notably, mean plant height for both the DH and parental lines was significantly greater in Shanghai (30.89° N, 121.38° E) than in Hainan (18.55° N, 110.05° E), a trend consistent across both growing seasons (Table 1). These results underscore the profound impact of environmental conditions on rice plant height. Indeed, this environmental responsiveness is characteristic of ‘Shenyou 26’, a photoperiod-sensitive japonica hybrid particularly well-adapted to Shanghai and similar latitudes.
ANOVA analysis was performed to elucidate the sources of plant height variation in the DH population. The resulting variance components and broad-sense heritability (H^2^) are summarized in Table 2. The analysis revealed that the environmental variance (σ^2^E = 662.36) was the largest component and substantially greater than the genotypic variance (σ^2^G = 86.01) and the genotype-by-environment interaction variance (σ^2^GE = 48.21), indicating that environmental variance constitutes the predominant proportion of the total phenotypic variation. This result further reinforces the profound influence of environmental conditions on rice plant height.
Despite the substantial environmental influence, the broad-sense heritability (H^2^) for plant height was 0.88 (Table 2). This high heritability of plant height indicates that, within a given environment, the observed phenotypic variation among the parental and DH lines is largely attributable to genetic factors, confirming that plant height is a highly heritable trait in this population. This conclusion is also supported by the consistently greater plant height of ‘Shenhui 26’ compared to ‘Shen 9A’ in both Shanghai (30.89° N, 121.38° E) and Hainan (18.55° N, 110.05° E) across two growing seasons (Table 1).
2.3. QTL Identified for the Plant Height in the DH Population
Based on a previously constructed genetic linkage map comprising 470 SNP markers for this DH population [22], a QTL analysis was conducted to identify genomic regions associated with plant height. An LOD threshold of 3.0, determined by 1000 permutations, was applied to declare significant QTLs. A total of seven QTLs were identified on chromosomes 1, 2, 5, 6, 9, and 12 (Table 3). Among these, qPH1.1 was consistently detected across all four environments, while qPH5.1 and qPH9.1 were identified in three environments. The remaining four QTLs were environment-specific, each detected in only one environment. The QTLs qPH1.1, qPH9.1, and qPH9.2 exhibited positive additive effects, indicating that the allele from ‘Shenhui 26’ contributed to increased plant height.
2.4. The SD1 Gene Is a Probable Candidate for the qPH1.1
The stable QTL qPH1.1, which was consistently detected across all four environments, was mapped to a region on chromosome 1 that is proximal to the semi-dwarf gene SD1, which is widely recognized as a cornerstone of the ‘Green Revolution’ in rice breeding due to its crucial role in determining plant height. A pronounced reduction in nucleotide diversity has been observed around the SD1 locus in japonica landraces, a pattern not detected in indica landraces or O. rufipogon, indicating that the SD1 gene underwent strong artificial selection during japonica domestication [5]. Compared to the indica variety ‘Kasalath’, two single-base mutations in the SD1 gene of the japonica cultivar ‘Nipponbare’ lead to two amino acid substitutions (G100E and R340Q) in the encoded GA20 oxidase, which significantly reduces plant height [5].
Sequence analysis revealed that the restorer line ‘Shenhui 26’ carries the SD1-EQ allele, identical to that reported in ‘Nipponbare’ [5]. In contrast, the male sterile line ‘Shen 9A’ harbors both the characteristic SD1-EQ mutations and two additional nucleotide substitutions in exon 3 (Figure 3A), resulting in an amino acid change (D349H). Further analysis identified that SD1-EQH in ‘Shen 9A’ is identical to the sd1-r allele derived from ‘Reimei’ [8]. Multiple sequence alignment revealed that the aspartic acid at position 349 is highly conserved in GA20 oxidase across diverse Poaceae species (Figure 3B), suggesting that this residue is likely required for the full activity of GA20 oxidase. Therefore, the D349H substitution may impair enzyme function, thereby contributing to the plant height variation observed in ‘Shen 9A’.
A haplotype analysis of the SD1 gene was performed using the MBKbase database (www.mbkbase.org) [23]. The results revealed that 1343 accessions carried the SD1-EQ allele, which is identical to that in ‘Shenhui 26’, while only 82 accessions possessed the SD1-EQH allele matching that of ‘Shen 9A’. Comparative analysis of plant height between accessions with these two haplotypes showed that those carrying the SD1-EQH allele were significantly shorter than those with the SD1-EQH allele (p < 0.05) (Figure 4), indicating that the D349H substitution may impair the biochemical function of the protein. Taken together, these results all support SD1 as the causal gene underlying the qPH1.1 locus. Furthermore, the SD1-EQH allele represents a valuable genetic resource for fine-tuning plant height in japonica rice breeding programs.
2.5. Genetic Validation for the qPH9.1 Locus
The QTL mapping results revealed that the qPH9.1 locus was consistently detected in three environments (2021 Shanghai, 2021 Hainan, and 2023 Hainan), explaining a considerable proportion of phenotypic variance with contribution rates of 7.42%, 22.72%, and 8.36%, respectively (Table 3). To further validate the effect of the qPH9.1 locus, two flanking InDel markers, C9-16503961 and C9-17927666 (Table S1, Figure S1), were developed to genotype a BC_1_F_2_ population consisting of 96 lines derived from a backcross with ‘Shen 9A’ as the recurrent parent. Among these lines, 17 carried the qPH9.1^Shenhui26^ allele and 21 carried the qPH9.1^Shen9A^ allele. Phenotypic comparison showed that lines with the qPH9.1^Shenhui26^ allele were significantly taller than those with the qPH9.1^Shen9A^ allele (Figure 5). Although minor background effects could not be completely excluded, the significant difference in mean plant height between the two genotypes strongly supports the conclusion that the qPH9.1^Shenhui26^ allele positively regulates plant height. These results confirm that the qPH9.1 locus has potential value for modifying plant height in rice breeding.
3. Discussion
Hybrid rice has made substantial contributions to global food security [24]. The elite japonica three-line hybrid rice ‘Shenyou 26’, developed by the Shanghai Academy of Agricultural Sciences, was derived from a cross between the BT-type male sterile line ‘Shen 9A’ and the restorer line ‘Shenhui 26’. The hybrid was approved by the Shanghai Crop Variety Approval Committee in 2017 and shows good adaptability in the suburban areas of Shanghai, southern Jiangsu, northern Zhejiang, and southern Anhui provinces [21]. From 2019 to 2021, yield evaluations conducted across ten seed production bases in the suburban areas of Shanghai, covering a total of 3334 mu of ‘Shenyou 26’ seed production fields, showed an average seed yield of 245.5 kg per mu. One important factor influencing successful seed production in ‘Shenyou 26’ seed production is the plant height relationship between parental lines. Specifically, having the restorer line ‘Shenhui 26’ slightly taller than the male sterile line ‘Shen9A’ promotes efficient pollination and maximizes seed yield (Figure 1).
In this study, we investigated the genetic basis underlying variation in plant height between the A-line ‘Shen 9A’ and the R-line ‘Shenhui 26’ in the elite japonica hybrid ‘Shenyou 26’. Through multi-environment phenotyping and QTL mapping, we identified seven QTLs associated with plant height. These findings not only deepen our understanding of the genetic architecture of plant height in japonica rice but also provide practically valuable alleles for optimizing plant height in rice breeding.
The QTL qPH1.1, stably detected across all environments, was mapped to the genomic region containing the semi-dwarf gene SD1. Sequence comparison revealed that ‘Shenhui 26’ carries the SD1-EQ allele [5], whereas ‘Shen 9A’ harbors a different allele (SD1-EQH/sd1-r) with a D349H substitution in GA20 oxidase [8,25]. This substitution affects a highly conserved residue across grasses (Figure 3B) and may reduce enzymatic activity, potentially contributing to the reduced height of ‘Shen 9A’. Haplotype analysis in a large germplasm panel further demonstrated that accessions with SD1-EQH were significantly shorter than those carrying SD1-EQ (Figure 4), providing further evidence that SD1 is the causal gene underlying qPH1.1. Recent research showed that replacing the non-functional SD1 allele in the indica cultivar ‘9311’ with the weak allele SD1-EQ led not only to a substantial increase in plant height but also to an increase in yield per plant [7]. Thus, the SD1-EQ and SD1-EQH alleles represent variants with different strengths and may serve as valuable genetic resources for fine-tuning plant height in rice breeding, as they confer different levels of height reduction.
In addition to qPH1.1, we validated the genetic effect of qPH9.1, which explained 7.4181% to 22.7% of the phenotypic variance in various environments. Backcross validation demonstrated that the qPH9.1^Shenhui26^ allele increases plant height relative to the qPH9.1^Shen9A^ allele. The identification of qPH9.1 complements the long-recognized contribution of SD1, indicating that multiple loci contribute to plant height variation in the ‘Shenyou 26’-derived DH population. Such QTLs provide promising new targets for marker-assisted selection and may help overcome the drawbacks of overreliance on sd1 in rice plant height manipulation.
Our analysis also underscored the profound influence of the environment on plant height expression. Although broad-sense heritability was high (H^2^ = 0.88), environmental variance accounted for the largest proportion of phenotypic variation. Consistent with the photoperiod sensitivity of ‘Shenyou 26’, plants grown in Shanghai were significantly taller than those in Hainan across both years (Table 1). These results highlight the importance of considering genotype–environment interactions when manipulating plant height in rice breeding. In particular, when developing varieties adapted to different ecological regions, it is crucial to select appropriate semi-dwarf alleles or plant height-related genetic resources that match the specific environmental conditions of each region.
Together, our results reveal that plant height variation between ‘Shen 9A’ and ‘Shenhui 26’ is influenced by multiple loci, including qPH1.1 (SD1) and qPH9.1. The identification of multiple allelic variants offers new opportunities for fine-tuning plant architecture in rice breeding. Incorporating these loci into breeding programs may help balance plant height and yield potential, reduce lodging risk, and improve pollination efficiency in hybrid seed production. Future work focusing on the cloning and functional characterization of qPH9.1, as well as exploring its interaction with other height-related genes, will further enhance our understanding of plant height regulation and facilitate the development of high-yielding, environmentally adaptable rice varieties.
4. Materials and Methods
4.1. Plant Materials and Field Trials
A doubled haploid (DH) population comprising 232 individual lines derived from the elite japonica hybrid rice ‘Shenyou 26’ was used to map QTLs associated with plant height regulation [22]. ‘Shenyou 26’ was developed from a cross between the BT-type cytoplasmic male sterile (CMS) line ‘Shen 9A’ (used as the female parent) and the restorer line ‘Shenhui 26’ (used as the pollen donor) [21].
The DH population lines and their parental lines were cultivated from May to November in Shanghai (30.89° N, 121.38° E) and from December to April of the following year in Hainan (18.55° N, 110.05° E), China. A randomized complete block design with two replications was implemented at each location. Each plot consisted of six rows, with six plants per row. Field management practices during the growing season followed local farmers’ standard methods. Plant height was measured from ten representative plants located in the central area of each plot, with height defined as the distance from the soil surface to the top of the panicle.
4.2. ANOVA Analysis
Analysis of variance (ANOVA) was conducted using QTL IciMapping software [26]. The linear model used for ANOVA followed the description of Yin et al. [27]:
where y_ijk_ is the phenotypic value of the i-th (i = 1232) genotype in the j-th (j = 14) environment and the k-th (k = 1∼2) replication, μ is the overall mean of the DH population, R_k/j_ is the replication effect within environment j, G_i_ is the genotypic effect, E_j_ is the environmental effect, (GE)ij is the genotype-by-environment interaction, and ε_ijk_ is the random error.
Variance components, including genotypic variance (σ^2^G), environmental variance (σ^2^E), genotype-by-environment interaction variance (σ^2^G×E), and error variance (σ^2^ε), were estimated from the ANOVA results. Broad-sense heritability (H^2^) was calculated as:
where e is the number of environments and r is the number of replications [27].
4.3. QTL Mapping
A genetic linkage map, previously constructed [22], was used for QTL mapping. QTLs associated with rice plant height were identified using Inclusive Composite Interval Mapping (ICIM) implemented in QTL IciMapping software [26]. To determine significant QTLs, an LOD threshold of 3 was applied, which was estimated from 1000 permutations at a genome-wide type I error rate of 0.05. The phenotypic variance explained (PVE) and additive effects of each detected QTL were also calculated to evaluate their contribution to plant height variation. The confidence interval for each QTL was defined as the genomic region corresponding to a 1.5-LOD drop from the peak LOD score, which approximates a 95% confidence interval. QTLs detected in different environments were considered to represent the same locus if their confidence intervals overlapped.
4.4. Validation for the qPH9.1 Locus
To validate the genetic effects of qPH9.1, the hybrid rice ‘Shenyou 26’ was backcrossed with the A-line ‘Shen 9A’. The resultant BC_1_F_1_ plants were genotyped using the flanking InDel markers C9-16503961 and C9-17927666. Heterozygous plants at the qPH9.1 locus were self-pollinated to produce BC_1_F_2_ seeds. In 2025, the BC_1_F_2_ population was planted in Shanghai (30.89° N, 121.38° E) and genotyped with the same InDel markers. Plants homozygous for either qPH9.1^Shenhui26^ or qPH9.1^Shen9A^ were selected, and their plant heights were measured to compare phenotypic differences between the two genotypes.
5. Conclusions
Plant height variation between the parental lines of ‘Shenyou 26’ is controlled by multiple loci, including SD1 allelic variants and qPH9.1. The complementary effects of these alleles enable precise regulation of plant height, providing valuable genetic resources for optimizing plant architecture and balancing the lodging resistance with yield in rice breeding.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Huang Y. Xie Z. Chen D. Chen H. Zeng Y. Dai S. Prediction of Rice Plant Height Using Linear Regression Model by Pyramiding Plant Height-Related Alleles Int. J. Mol. Sci.202526624910.3390/ijms 2613624940650027 PMC 12249533 · doi ↗ · pubmed ↗
- 2Hedden P. The genes of the Green Revolution Trends Genet.2003195910.1016/S 0168-9525(02)00009-412493241 · doi ↗ · pubmed ↗
- 3Peng J. Richards D.E. Hartley N.M. Murphy G.P. Devos K.M. Flintham J.E. Beales J. Fish L.J. Worland A.J. Pelica F. ‘Green revolution’ genes encode mutant gibberellin response modulators Nature 199940025626110.1038/2230710421366 · doi ↗ · pubmed ↗
- 4Cheng X. Huang Y. Tan Y. Tan L. Yin J. Zou G. Potentially Useful Dwarfing or Semi-dwarfing Genes in Rice Breeding in Addition to the sd 1 Gene Rice 2022156610.1186/s 12284-022-00615-y 36542176 PMC 9772376 · doi ↗ · pubmed ↗
- 5Asano K. Yamasaki M. Takuno S. Miura K. Katagiri S. Ito T. Doi K. Wu J. Ebana K. Matsumoto T. Artificial selection for a green revolution gene during japonica rice domestication Proc. Natl. Acad. Sci. USA 2011108110341103910.1073/pnas.101949010821646530 PMC 3131315 · doi ↗ · pubmed ↗
- 6Zhang L. Bian Z. Ma B. Li X. Zou Y. Xie D. Liu J. Ren Y. Zhang C. Wang J. Exploration and selection of elite Sd 1 alleles for rice design breeding Mol. Breed.2020407910.1007/s 11032-020-01161-5 · doi ↗
- 7Yu Y. Hu X. Zhu Y. Mao D. Re-evaluation of the rice ‘Green Revolution’ gene: The weak allele SD 1-EQ from japonica rice may be beneficial for super indica rice breeding in the post-Green Revolution era Mol. Breed.2020408410.1007/s 11032-020-01164-2 · doi ↗
- 8Asano K. Takashi T. Miura K. Qian Q. Kitano H. Matsuoka M. Ashikari M. Genetic and Molecular Analysis of Utility of sd 1 Alleles in Rice Breeding Breed. Sci.200757535810.1270/jsbbs.57.53 · doi ↗