Saucerneol D Suppresses the Growth of Helicobacter pylori and Their Virulence Factors
Su Man Kim, Hyun Jun Woo, Zhongduo Yang, Tiankun Zhao, Ji Yeong Yang, Sa-Hyun Kim

TL;DR
Saucerneol D, a natural compound, inhibits the growth and harmful effects of Helicobacter pylori bacteria.
Contribution
This study is the first to report the antibacterial and anti-virulence effects of saucerneol D against H. pylori.
Findings
Saucerneol D suppresses H. pylori growth by reducing dnaN and polA gene expression.
It decreases the secretion of CagA toxin by downregulating type IV secretion system proteins.
Saucerneol D reduces urease production and motility-related gene flaB expression.
Abstract
Helicobacter pylori infects the human stomach and causes various gastrointestinal diseases. Saucerneol D is a type of lignan, which is a polyphenol compound that exists naturally in plants, and it is abundant in flaxseed, sesame seeds, whole grains, vegetables, and fruits. Saucerneol D is found in Saurus chinensis extract and has been reported to exert a variety of effects, such as antioxidant and anti-inflammatory abilities. However, its antibacterial effect against H. pylori has not been reported; therefore, we analyzed the effect of saucerneol D on H. pylori in the present study. Changes in the expression of pathogenic factors and gene transcription in H. pylori were observed after treatment with saucerneol D using Western blotting and RT-PCR. It was confirmed that saucerneol D suppressed the growth of H. pylori by decreasing the expression of the genes dnaN and polA, which are…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
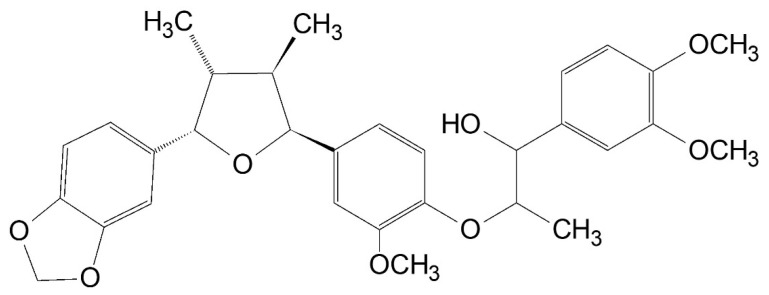 Figure 1
Figure 1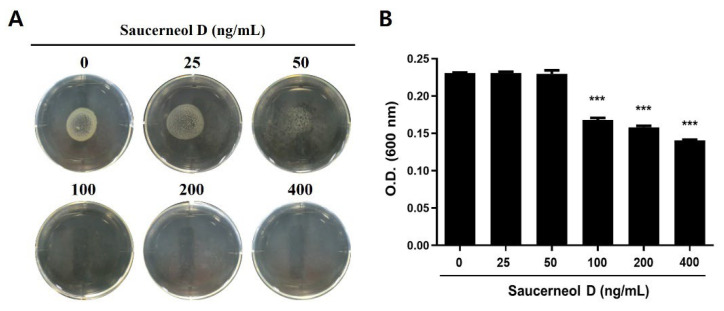 Figure 2
Figure 2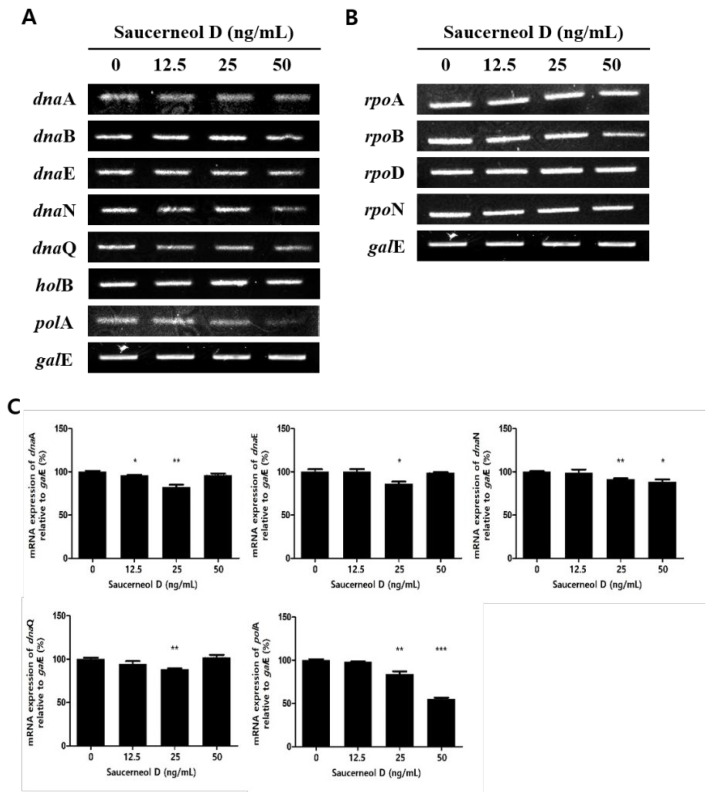 Figure 3
Figure 3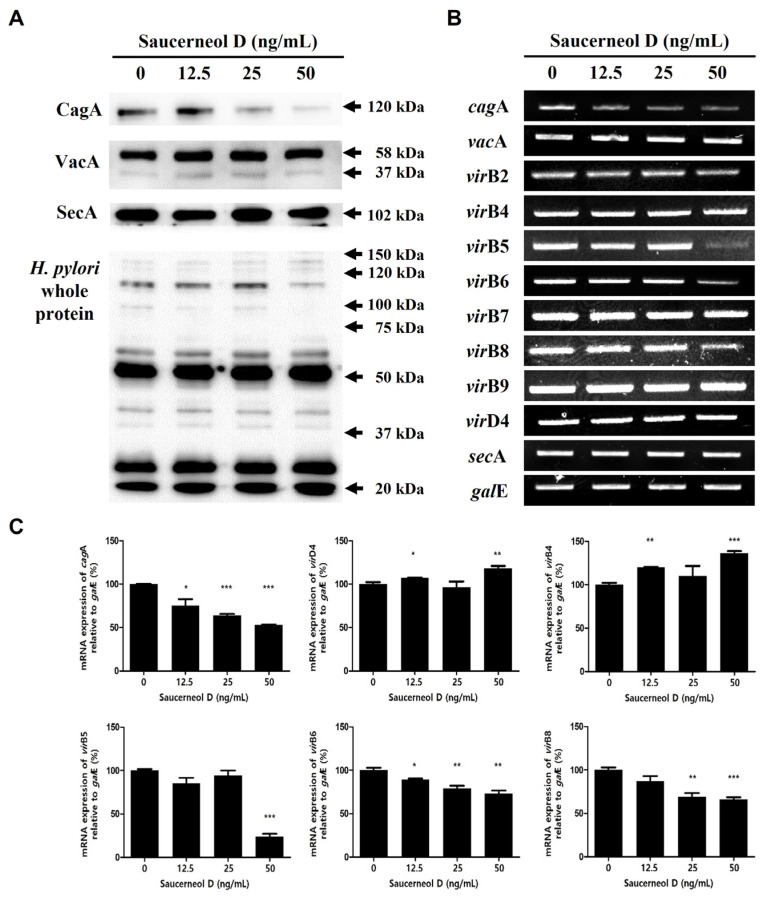 Figure 4
Figure 4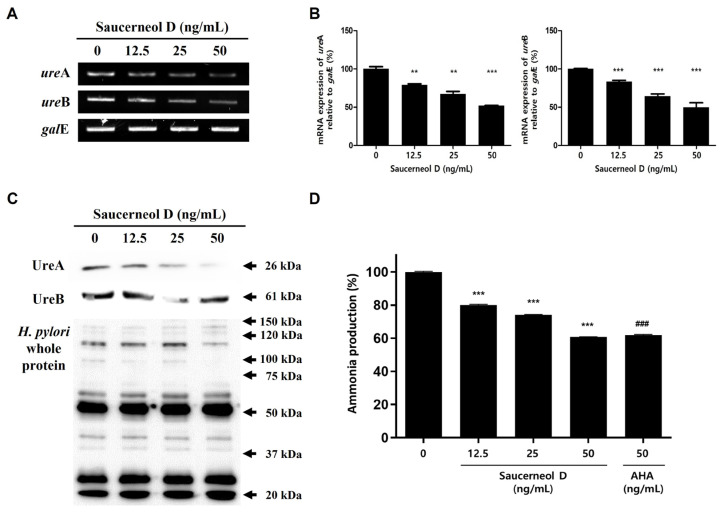 Figure 5
Figure 5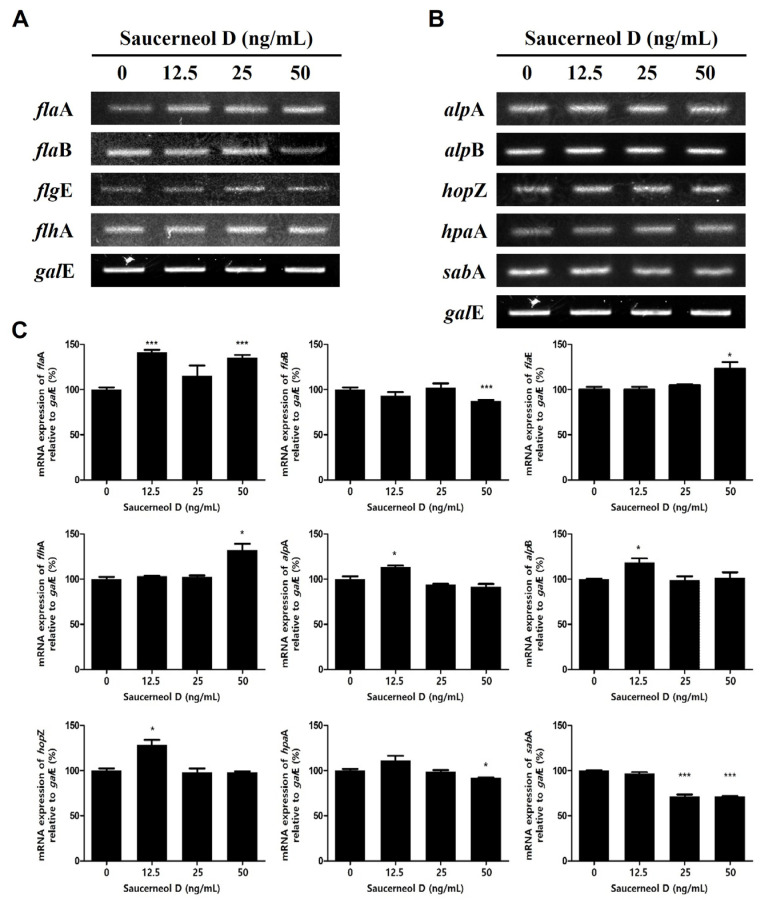 Figure 6
Figure 6 Figure 7
Figure 7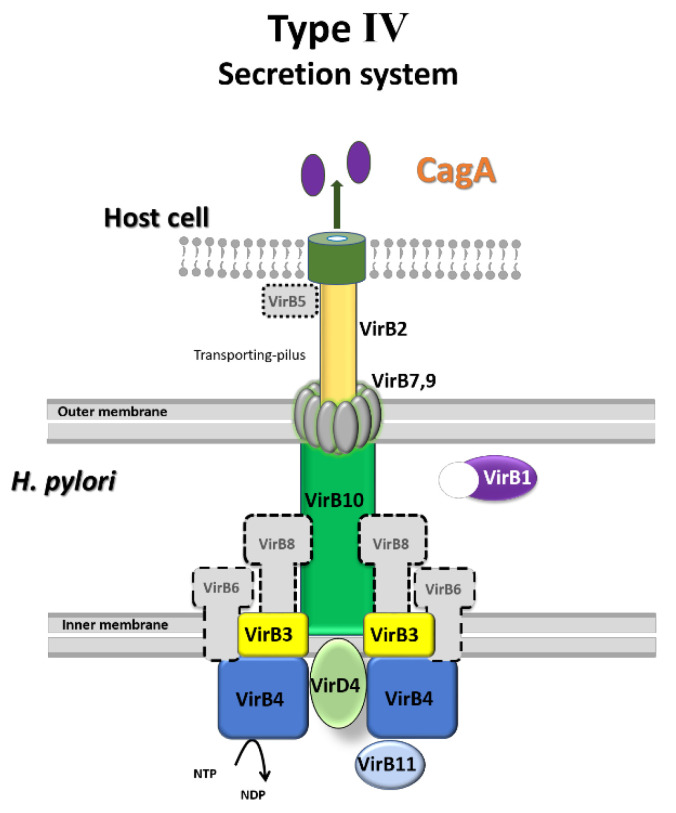 Figure 8
Figure 8- —Ministry of Education
- —Korea government (MSIT)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHelicobacter pylori-related gastroenterology studies · Galectins and Cancer Biology · Gastrointestinal disorders and treatments
1. Introduction
Helicobacter pylori is a Gram-negative, spiral, and microaerophilic bacterium that is a significant pathogen responsible for various gastrointestinal diseases, including superficial gastritis, chronic gastritis, peptic ulcer, gastric mucosa-associated lymphoid tissue (MALT) lymphoma, intestinal metaplasia, and gastric cancer [1]. Thus, the World Health Organization classified H. pylori as a class I carcinogen in 1994 [2]. In recent years, the recommended primary treatment for H. pylori infection in Korea has been a triple-therapy regimen consisting of proton pump inhibitors (PPIs), clarithromycin, and amoxicillin for 10–14 days [3,4]. If the primary treatment fails, a secondary treatment using a four-drug regimen of PPIs, metronidazole, bismuth, and tetracycline can be employed [5]. Over the past decade, the rate of resistance to clarithromycin has increased, and the WHO has prioritized research and development of antibiotics to treat clarithromycin-resistant H. pylori [6,7]. Additionally, the tolerance rate to metronidazole has been reported to be over 15% worldwide [7]. These reports indicate that the antibiotic resistance rate of H. pylori is gradually increasing, necessitating the search for new treatments.
Cytotoxin-associated antigen A (CagA), the most important pathogenic factor in H. pylori, is encoded by the cag pathogenicity island (PAI) [8]. CagA is injected into gastric epithelial cells by the type IV secretion system (T4SS), which is also encoded by cagPAI [9]. It has been suggested that CagA significantly affects the transcriptional activation of interleukin-8 (IL-8) through the activation of mitogen-activated protein kinase (MAPK) and the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and induces a cell membrane modification termed the hummingbird phenotype [10,11].
Another crucial H. pylori pathogenic factor, vacuolating cytotoxin A (VacA), is secreted by all H. pylori strains through the type Va secretion system (T5aSS) [12,13]. The secretion system subunit protein A (SecA) is an essential protein in T5aSS, providing the energy required for VacA to be secreted outside the bacterial membrane [14]. Once secreted, VacA binds to surface receptors on host cells to form anion channels, enters cells through clathrin-independent endocytosis, induces necrosis or apoptosis, and results in VacA-dependent vacuole formation [15,16].
Urease is an enzyme necessary for H. pylori to colonize the highly acidic environment of the stomach. Urease catalyzes the conversion of urea in the stomach into carbon dioxide (CO_2_) and ammonia (NH_3_). The ammonia produced neutralizes the gastric environment by increasing the pH [17]. Therefore, the urease enzyme is a known therapeutic target for the eradication of H. pylori.
Saururus chinensis (Lour.) Baill., a perennial herb native to East Asia, is distributed on Jeju island and mountain Jiri in Korea [18]. Extracts of S. chinensis have been studied for their antioxidant activity, hepatocellular protection, and antibacterial properties. Saucerneol D, a lignan found in the roots of Saururus chinensis, has been used to treat several inflammatory conditions, including jaundice, pneumonia, edema, and fever [18,19]. Saucerneol D, one of the lignans of S. chinensis, has the chemical structure shown in Figure 1 [20]. However, there is no report of saucerneol D having antibacterial effects. Therefore, this research investigated the inhibitory effect and mechanism of saucerneol D on H. pylori, as well as its impact on the reduction in various virulence factors of H. pylori in vitro.
2. Materials and Methods
2.1. Bacterial Culture
The H. pylori reference strain of ATCC 49503 was purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). H. pylori were grown on Brucella agar plates (BD Biosciences, Franklin Lakes, NJ, USA), with selectively supplemented H. pylori (Oxoid Limited, Hampshire, UK) and heat-inactivated 10% bovine serum (Gibco, Grand Island, NE, USA) placed under microlender microaerophilic and 100% humidity conditions at 37 °C and inspected after three to four days.
2.2. Determination of Minimum Inhibitory Concentration (MIC)
For the dilution test, H. pylori ATCC 49503 strain colonies grown on Brucella agar plates were collected and resuspended in sterile saline. Saucerceol D was obtained from the Natural Products Bank, National Institute for Korean Medicine Development (Gyeongsan, Gyeongsangbuk, Republic of Korea), and was dissolved using DMSO (dimethyl sulfoxide) as a solvent. To test the agar dilution, the number of bacterial particles in the H. pylori suspension was set to McFarland 3.3. An amount of 10 µL of the bacterial suspension was placed on Mueller–Hinton agar plates with heat-inactivated 10% bovine serum including indicated concentrations of saucerneol D. For liquid dilution experiments, the number of bacterial particles in the H. pylori suspension was set to a McFarland scale of 0.5. Bacteria were cultured for 72 h in broths treated with various concentrations of saucerneol D. All the solutions were prepared in such a manner that the final DMSO concentration was the same in all treatments. The final optical density (600 nm) of the bacterial suspension was measured by spectrophotometry.
2.3. RNA Extraction and Reverse Transcriptase Polymerase Chain Reaction (RT-PCR)
Cultured H. pylori were washed twice with sterile saline, and total RNA was extracted using easy-BLUE™ Total RNA Extraction Kit (iNtRON Biotechnology, Seongnam, Republic of Korea) as described in the manufacturer’s instructions. The reverse transcriptase polymerase chain reaction (RT-PCR) primer sequences used in this study are listed in Table 1. GalE (UDP-Galactose-4-Epimerase) was used as an internal control. Gel images were taken using the Molecular Image Gel DocTM XR+ system (Bio-Rad, Hercules, CA, USA). The band intensity of PCR product was analyzed with the ImageLab 6.1 software (BioRad, Hercules, CA, USA).
2.4. Anti-H. pylori SecA or Anti-H. pylori Polyclonal Antibody Production
We produced rabbit anti-SecA or rabbit anti-H. pylori (ATCC 49503) polyclonal antibodies following a method described previously [25]. New Zealand White rabbits (8~10 weeks) were purchased from Central Lab Animal Inc. (Seoul, Republic of Korea), and the SecA polyclonal antiserum was produced by injection of 500 μg of 22 mer peptides (SecA: GTERHESRRIDNQLRGRSGRQG; nucleotide 518–539) through ear vein. The H. pylori polyclonal antibodies were generated by immunization with intravenous injection with formaldehyde-fixed 1 × 10^8^/mL concentration of H. pylori (ATCC 49503) bacteria every week for a total of six weeks. After six weeks, about 100 mL of whole blood was collected from the heart and then allowed to clot in a refrigerator overnight, and the serum was isolated after centrifugation. Pre-immune serum was harvested prior to immunization. Antibodies were purified using a protein A column. The rabbit anti-SecA antibodies were used at a 1:5000 dilution in 5% skim milk blocking solution. The rabbit anti-H. pylori (ATCC 49503) polyclonal antibodies were used at a 1:10,000 dilution.
2.5. Protein Extraction and Western Blot
The bacterial cell lysate was prepared to detect SecA molecules. Bacteria were lysed with a radio immunoprecipitation assay lysis buffer (Biosesang, Seongnam, Republic of Korea) containing a protease inhibitor cocktail, it was then incubated on ice for 30 min. To lyse the bacterial cells completely, the mixture was sonicated for 2 min in 10 s intervals (Sonicator XL-2020, Heat Systems Ultrasonics, Pittsburgh, PA, USA). The cell lysates were then centrifuged at 14,000 rpm at 4 °C for 10 min, and the supernatants were collected. Proteins were quantified by Lowry Protein assay (Bio-Rad) and mixed with ×5 sodium dodecyl sulfate (SDS) sample loading buffer. Protein samples were separated by SDS-polyacrylamide gel electrophoresis for 90 min at 120 voltage and transferred to a nitrocellulose membrane for 90 min at 400 mA. The bacterial culture supernatant was also collected to detect CagA VacA or urease, and then it was concentrated 10-fold using Amicon Centricon centrifugal filters (3 kDa cut-off) (Millipore, Billerica, MA, USA) at 3000 rpm for 2 h at 4 °C. Polyclonal antibodies against whole H. pylori ATCC 49503 proteins were used as an internal control.
2.6. Urease Activity Test
H. pylori ATCC 49503 strain was grown in urea broth containing 0.1% urea with the indicated concentrations of saucerneol D (12.5, 25, and 50 ng/mL) and acetohydroxamic acid (AHA) (50 ng/mL). After 1 h, centrifugation was performed at 3000 rpm for 10 min and then 1 mL of supernatant was extracted. Urease activity was confirmed by measuring the amount of ammonia using an Asan Set Ammonia kit (Asan Pharmaceutical, Seoul, Republic of Korea) according to the manufacturer’s instructions. Ammonia concentrations in the specimens were calculated using the standard curve.
2.7. Statistical Analysis
Data in the bar graphs are presented as mean ± standard error of mean (SEM). All the statistical analyses were performed using GraphPad Prism 7.0 software (GraphPad Software, San Diego, CA, USA). All the data were analyzed using an unpaired Student’s t-test, and p < 0.05 was considered statistically significant (* p < 0.05, ** p < 0.01, and *** p < 0.001). Every experiment was repeated at least three times to verify the results and perform statistical analysis.
3. Results
3.1. Determination of the MIC
The agar dilution test was performed to determine the minimum inhibitory concentration (MIC) of saucerneol D against H. pylori. Mueller–Hinton agar containing heat-inactivated 10% bovine serum and varying concentrations of saucerneol D (25, 50, 100, 200, and 400 ng/mL) was prepared, and H. pylori were grown on the agar plates for 72 h. According to the agar dilution test, the MIC of saucerneol D against H. pylori was found to be 100 ng/mL, indicating that even low concentrations were sufficient to suppress bacterial growth (Figure 2A). The MIC was also confirmed by a broth dilution test, which showed bacterial growth inhibition at concentrations above 100 ng/mL of saucerneol D, consistent with the agar dilution test results (Figure 2B). Therefore, to avoid bactericidal conditions and better assess mechanistic effects, subsequent experiments were performed with saucerneol D concentrations below 50 ng/mL.
3.2. Effect on Replication and Transcription Genes in Saucerneol D-Treated H. pylori
To investigate the mechanism behind the inhibitory effect of saucerneol D on H. pylori, the bacteria were treated with sub-MICs of saucerneol D (12.5, 25, and 50 ng/mL). The results showed that the mRNA expression levels of dnaA, dnaN, dnaQ, and polA, among the replication genes of H. pylori, were decreased in saucerneol D-treated H. pylori (Figure 3A,C). In contrast, saucerneol D did not affect the transcription genes rpoA, rpoB, rpoD, and rpoN (Figure 3B,C).
3.3. Effect on CagA, VacA, and Their Secretion Systems in Saucerneol D-Treated H. pylori
H. pylori were treated with saucerneol D (12.5, 25, and 50 ng/mL) for 72 h in Mueller–Hinton broth containing heat-inactivated 10% bovine serum. After harvesting the protein and mRNA, Western blotting and RT-PCR analyses were performed. Saucerneol D decreased cagA expression and CagA protein secretion in a concentration-dependent manner, but the mRNA and protein secretions of VacA were not affected by saucerneol D (Figure 4A–C). Among the components of T4SS, which is the CagA secretory pathway, the expressions of virB5, virB6, and virB8 mRNA were decreased by saucerneol D, whereas those of virB4 and virD4 were increased somewhat (Figure 4B,C). The mRNA and protein expressions of SecA, a regulating component of the T5aSS for VacA secretion, were not affected by saucerneol D (Figure 4A–C).
3.4. Downregulation of Urease in Saucerneol D-Treated H. pylori
H. pylori’s urease enzyme is vital for its survival and colonization of the gastric mucosa [12]. It is composed of two subunits, UreA and UreB, encoded by ureA and ureB, respectively [26]; both of these were inhibited after treatment with saucerneol D (Figure 5A–C). To confirm the effect of reduced UreA and UreB expression on urease activity, the amount of ammonia produced by urease was measured. H. pylori were grown in brucella broth containing 0.1% urea with the indicated concentrations of saucerneol D (12.5, 25, and 50 ng/mL) and AHA (50 ng/mL). After 1 h of incubation, the ammonia produced was measured. Ammonia levels decreased in a concentration-dependent manner after treatment with saucerneol D, reaching similar levels as those treated with AHA at the same concentration (Figure 5D).
3.5. Effect on Flagella and Adhesion in Saucerneol D-Treated H. pylori
The movement and adhesion of H. pylori are critical for the successful infection of gastric epithelial cells. H. pylori’s movement is facilitated by flagella [27]. This study aimed to determine the effects of saucerneol D on the expression of flagella genes (flaA*, flaB, flgE,* and flhA) and adhesion genes (alpA*, alpB, hopZ, hpaA,* and sabA). Saucerneol D increased the expression of flaA*, flgE,* and flhA among the flagella genes while decreasing the expression of flaB (Figure 6A,C). These findings suggest that saucerneol D may interfere with the balance of flagellar components, potentially altering motility. Additionally, among the adhesion genes, only the expression of sabA was decreased by saucerneol D (Figure 6B,C). Together, these results indicate that saucerneol D not only affects bacterial motility genes but may also weaken the adhesion capacity of H. pylori.
4. Discussion
H. pylori is a major cause of gastric cancer, accounting for 75% of all cases [28,29]. However, the increasing resistance to clarithromycin and the limitations of current primary therapies have been widely reported [6,30,31]. This highlights the need for research into new therapeutic agents or adjuvants to inhibit H. pylori. Although the antibacterial activity of saucerneol D, which is a lignan, has not been studied, the antibacterial activity of extracts from Saururus chinensis (Lour.) Baill. has been documented [18,19]. This study investigated the inhibitory effects of saucerneol D, a component of the extract, on H. pylori and its pathogenic factors.
Extracts from S. chinensis (Lour.) Baill. are known to contain quercetin glycosides such as quercitrin and rutin, as well as lignans such as saucerneol D, manassantin A, manassantin B, and saucernetin, and their structural formulas are displayed in Figure 7 [20]. It appears that, along with other constituents of the crude extract, saucerneol D contributes to anti-H. pylori activity. Saucerneol D inhibited H. pylori growth by downregulating essential replication machinery genes (Figure 2 and Figure 3). Specifically, it reduced the mRNA expression of dnaN and polA (Figure 3). DNA polymerase III, which is crucial for bacterial replication, comprises core polymerases (DnaE, DnaQ, and HolE), a sliding clamp (DnaN), and a multiprotein clamp loader (DnaX, HolA, HolB, HolC, and HolD) [32]. DNA polymerase I, encoded by the polA gene, plays a key role in connecting Okazaki fragments, repairing DNA, and replacing RNA primers with DNA [33]. Since DnaN and PolA are indispensable for bacterial replication, this result suggests that saucerneol D inhibits H. pylori growth by targeting these replication components.
Saucerneol D not only inhibited the synthesis of CagA, a critical virulence factor of H. pylori, but it also increased the mRNA expression of virB4, virB9, and virD4 of the T4SS, which injects CagA into host cells, while decreasing virB5, virB6, and virB8 (Figure 4). VirB4 and VirD4 are required for substrate recruitment and transport to T4SS, VirB6 stabilizes VirB5, and VirB8 is involved in assembling the inner membrane complex [9,34,35]. VirB9 forms a transmembrane pore complex, essential for transferring the matrix through the peri-membrane space and anchoring to the membrane [36]. VirB5 links external pilus recipients and donor cells and is a component of the T4SS external pilus [9,37]. Overall, the reduction in pathogenic factors and key secretory pathway components suggests that CagA translocation into host cells might be hindered. The preferential effect on CagA–T4SS over VacA pathways suggests possible selectivity, which warrants further mechanistic investigation.
Urease secretion is also regulated by T5aSS [25]. SecA, an important protein constituting the T5aSS, provides the energy for urease or VacA secretion outside the cell membrane [14,25,38]. Although there were no changes in the transcription of secA or expression of SecA after saucerneol D treatment (Figure 4A,B), the reason for the decrease in urease secretion is thought to be due to the decrease in the expression of ureA and ureB (Figure 5A). Urease-negative mutant H. pylori cannot colonize the stomachs of nude mice or gnotobiotic piglets, indicating that urease activity is essential for colonization [39,40]. If saucerneol D caused a reduction in urease secretion, as suggested in Figure 5, this might deteriorate H. pylori’s colonization and viability in the acidic environment of the stomach. The decrease in UreA and UreB synthesis led to reduced ammonia production (Figure 5D). Ammonia not only neutralizes stomach acid but also directly damages gastric epithelial cells and induces inflammation [41,42]. Additionally, the CO_2_ produced by urease protects H. pylori from oxidative damage by inhibiting peroxynitrite, thus enhancing viability [43,44]. Overall, saucerneol D might reduce urease synthesis in H. pylori, preventing gastric environment neutralization, decreasing colonization and survival rates, and potentially reducing the inflammatory response of gastric epithelial cells.
Saucerneol D also downregulated flaB but upregulated flaA, flgE, and flhA among the H. pylori flagella genes (Figure 6A). FlaA and FlaB are flagellins, making up the flagellar filament [44,45]. FlgE anchors flagellar filaments to the export domain of the basal body, while FlhA is part of the export dome and regulates flagellin protein release [46,47]. Although flaB decreased, the increase in the other flagella genes suggests that H. pylori, when exposed to saucerneol D, might have a compensatory mechanism to escape saucerneol D stress.
Saucerneol D also suppressed the expression of sabA, an adhesion gene in H. pylori (Figure 6B). H. pylori adhesion is a key mechanism in inducing chronic gastritis and gastric cancer, with chronic infection increasing inflammation and sialyl-Lewis X expression [48,49,50]. SabA mediates H. pylori binding to inflamed gastric mucosa by recognizing sialyl-Lewis A and sialyl-Lewis X antigens [51]. Thus, the inhibition of sabA expression by saucerneol D may reduce H. pylori adhesion, chronic infection, and the inflammatory response.
Further experiments using human gastrointestinal cell models are needed to confirm the effects of saucerneol D on CagA and VacA translocation, host cell adherence, and motility. Moreover, in vivo experiments using animal infection models are needed to substantiate these findings, and the bioavailability and gastric stability of saucerneol D remain unknown. Future studies should evaluate these properties in simulated gastric fluids and in vivo models. In addition, the potential toxicity and selectivity of saucerneol D toward host cells should be evaluated in future studies to assess its suitability as a therapeutic candidate.
This study demonstrated that saucerneol D inhibits the growth of H. pylori and reduces its virulence factors. To summarize, saucerneol D decreased the expression of CagA virulence factors and related secretion systems, including T4SS. Saucerneol D also downregulated urease subunit proteins, thereby reducing urease activity.
5. Conclusions
In conclusion, we found that saucerneol D acts as a potent inhibitor of H. pylori multiplication and of virulence factors—CagA, urease, and sabA—involved in H. pylori pathogenesis. H. pylori growth was inhibited through the downregulation of its replication machinery when exposed to saucerneol D (Figure 3). In addition, saucerneol D decreased the CagA secretion of H. pylori by downregulating the T4SS-comprising molecules VirB5, VirB6, and VirB8 (Figure 8). Saucerneol D also downregulated the expression of H. pylori urease subunit proteins, thereby reducing urease activity. The expression of an adhesion molecule gene, sabA, was also reduced when exposed to saucerneol D. Following these results, we suggest saucerneol D as a potential H. pylori inhibitor. We have plans to re-examine the anti-H. pylori effect of saucerneol D in the future using a gastric cell line infection model or a Mongolian gerbil infection model.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Santacroce L. Topi S. Bottalico L. Charitos I.A. Jirillo E. Current Knowledge about Gastric Microbiota with Special Emphasis on Helicobacter pylori-Related Gastric Conditions Curr. Issues Mol. Biol.2024464991500910.3390/cimb 4605029938785567 PMC 11119845 · doi ↗ · pubmed ↗
- 2Møller H. Heseltine E. Vainio H. Working group report on schistosomes, liver flukes and Helicobacter pylori Int. J. Cancer 19956058758910.1002/ijc.29106005027860130 · doi ↗ · pubmed ↗
- 3Chey W.D. Leontiadis G.I. Howden C.W. Moss S.F. ACG Clinical Guideline: Treatment of Helicobacter pylori Infection Am. J. Gastroenterol.201711221223910.1038/ajg.2016.56328071659 · doi ↗ · pubmed ↗
- 4Jung H.K. Kang S.J. Lee Y.C. Yang H.J. Park S.Y. Shin C.M. Kim S.E. Lim H.C. Kim J.H. Nam S.Y. Evidence based guidelines for the treatment of Helicobacter pylori infection in Korea 2020 Korean J. Intern. Med.20213680783810.3904/kjim.2020.70134092054 PMC 8273819 · doi ↗ · pubmed ↗
- 5Valada P. Mata A. Brito R.M.M. Gonçalves T. Medeiros J.A. Nogueira C. Detection of Helicobacter pylori and the Genotypes of Resistance to Clarithromycin, Fluoroquinolones, and Metronidazole in Gastric Biopsies: An In Silico Analysis to Help Understand Antibiotic Resistance Curr. Issues Mol. Biol.20254718710.3390/cimb 4703018740136441 PMC 11940985 · doi ↗ · pubmed ↗
- 6Ansari S. Yamaoka Y. Helicobacter pylori Infection, Its Laboratory Diagnosis, and Antimicrobial Resistance: A Perspective of Clinical Relevance Clin. Microbiol. Rev.202235 e 002582110.1128/cmr.00258-2135404105 PMC 9491184 · doi ↗ · pubmed ↗
- 7Savoldi A. Carrara E. Graham D.Y. Conti M. Tacconelli E. Prevalence of Antibiotic Resistance in Helicobacter pylori: A Systematic Review and Meta-analysis in World Health Organization Regions Gastroenterology 201815513721382.e 1710.1053/j.gastro.2018.07.00729990487 PMC 6905086 · doi ↗ · pubmed ↗
- 8Censini S. Lange C. Xiang Z. Crabtree J.E. Ghiara P. Borodovsky M. Rappuoli R. Covacci A. cag, a pathogenicity island of Helicobacter pylori, encodes type I-specific and disease-associated virulence factors Proc. Natl. Acad. Sci. USA 199693146481465310.1073/pnas.93.25.146488962108 PMC 26189 · doi ↗ · pubmed ↗