Allergic Contact Dermatitis: Immunopathology and Potential Therapeutic Strategies
Anders Boutrup Funch, Carsten Geisler, Charlotte Menné Bonefeld

TL;DR
This paper reviews the immune mechanisms and treatment options for rapid-onset allergic contact dermatitis, a skin reaction that occurs quickly after allergen exposure.
Contribution
The paper distinguishes between rapid-onset and delayed-type ACD reactions and explores novel therapeutic strategies targeting immune pathways.
Findings
Rapid-onset ACD occurs within hours at previously sensitized skin sites.
Potential therapies include modulating cytokine signaling and T cell survival.
Current treatments face limitations in targeting redox balance and checkpoint pathways.
Abstract
Allergic contact dermatitis (ACD) is a common inflammatory skin disease induced by exposure of the skin to contact allergens. Classically, ACD is defined as a delayed-type (type IV) hypersensitivity reaction mediated by allergen-specific T cells, with symptoms peaking 48–72 h after exposure to the contact allergen. This delayed response to the contact allergen is seen during patch testing, where allergen-naïve, unaffected skin of allergic individuals is exposed to the contact allergen. However, in daily life and in certain occupational settings, allergic individuals often experience rapid flare-ups/exacerbations with intensely itching erythema, oedema, and often vesicles within hours after re-exposure to the specific contact allergen. These rapid flare-ups only develop at skin sites previously exposed to the contact allergen. Thus, it is important to distinguish between the rapid-onset…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
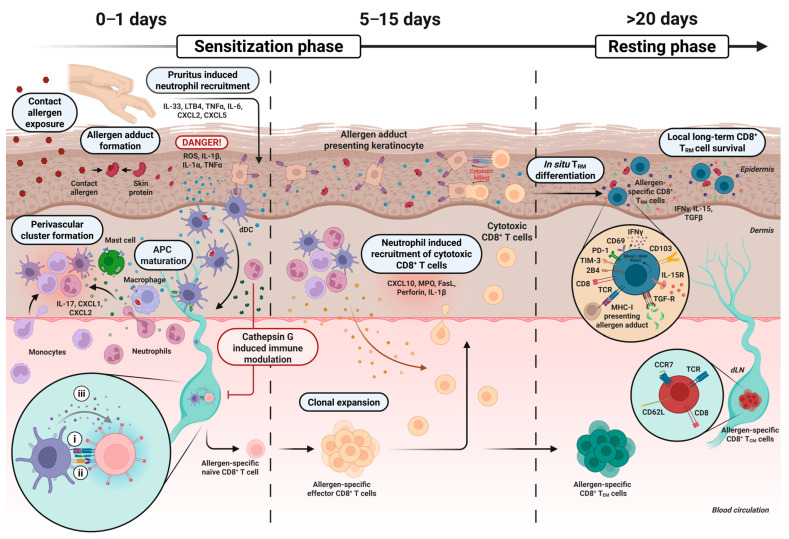 Figure 1
Figure 1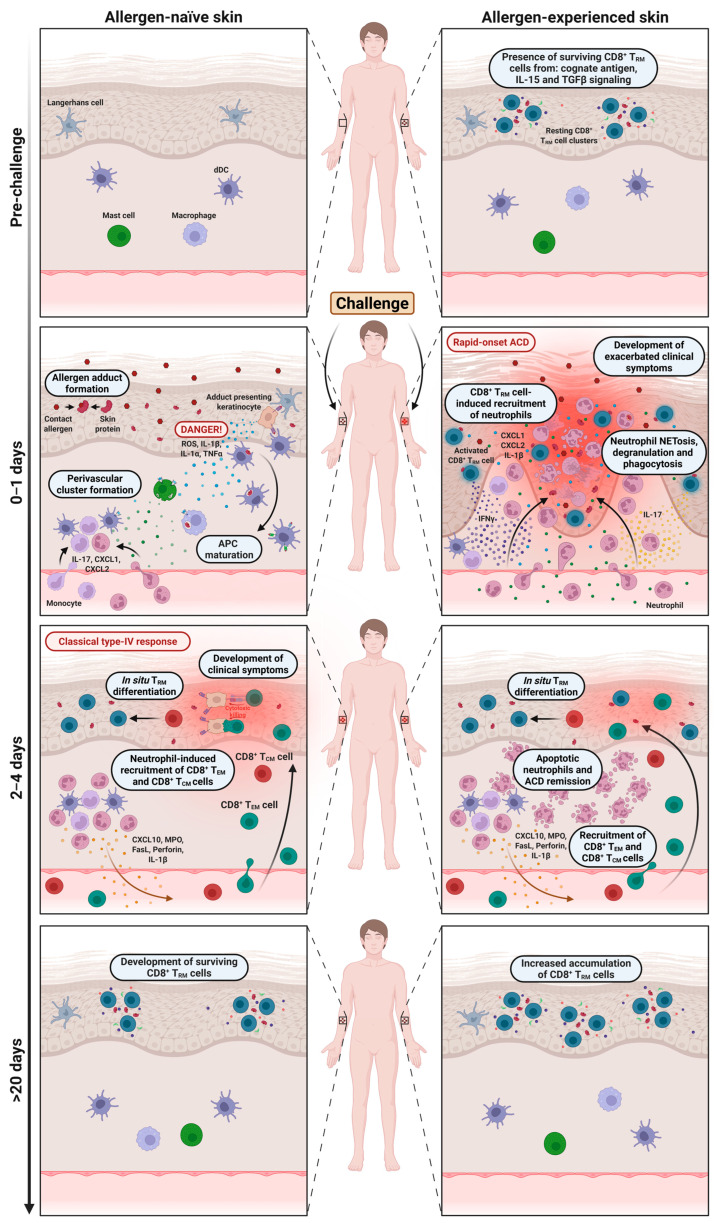 Figure 2
Figure 2- —Carlsberg Foundation
- —LEO Foundation
- —Independent Research Fund Denmark
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsContact Dermatitis and Allergies · Dermatology and Skin Diseases · Allergic Rhinitis and Sensitization
1. Introduction
Allergic contact dermatitis (ACD) is an inflammatory skin disease that occurs in sensitized individuals upon re-exposure to the sensitizing contact allergen [1]. The condition is highly prevalent, affecting approximately 20% of the population [2]. ACD is classically known as a delayed-type or type IV hypersensitivity reaction that is mediated by allergen-specific memory T cells. ACD manifests as local erythematous, pruritic, and occasionally blistering skin lesions that typically emerge within 48–72 h after exposure to the contact allergen, as observed during patch testing [1,3,4]. However, in daily life, patients often experience much faster (<24 h) reactions, here referred to as rapid-onset ACD, when exposed to the contact allergen in allergen-experienced skin areas [5,6,7]. The accelerated ACD response is particularly relevant in occupational settings, where repeated exposure to contact allergens in the same skin area is common. Occupational exposures are recognized as a major risk factor for ACD, with notably high prevalence rates among hairdressers, healthcare workers, and individuals in industries involving reactive chemicals, where complete avoidance of allergens is highly impractical [8,9,10]. Consequently, occupational ACD has considerable socio-economic repercussions, including frequent work absences [11,12]. Given these challenges and the current limited therapeutic options available, it is essential to study the immunopathology and identify potential therapeutic strategies, especially for individuals suffering from rapid-onset ACD.
It is known that CD8^+^ skin-resident memory T (T_RM_) cells develop locally in the skin after exposure to many contact allergens, and recently it was shown that the ability of CD8^+^ T_RM_ cells to recruit massive numbers of neutrophils into the skin is crucial for rapid-onset ACD [7,13,14,15,16]. The vast majority of infiltrating neutrophils disappear from the skin less than 48 h after exposure to the allergen [13]. Furthermore, only a limited number of neutrophils are found after exposure of allergen-naïve skin to contact allergens [7,13,16,17,18], which may explain the hitherto rather limited focus on neutrophils in ACD.
Neutrophils are short-lived immune cells that play a vital role in host defense as they are rapidly recruited to infected peripheral tissues, where they effectively phagocytose pathogens and damaged cells and exert a broad range of cytolytic effector functions. [19,20]. However, when inflammation becomes excessive or misdirected, as seen in rapid-onset ACD, neutrophil recruitment and uncontrolled release of their cytolytic granules can lead to substantial tissue damage [19,20]. In recent years, the interest in the pathological role of neutrophils in both murine models of ACD [13,14,21,22,23,24,25,26,27,28] and in human studies [16,18,29] has grown.
This review summarizes the current knowledge on the interplay between CD8^+^ T cells and neutrophils in both the sensitization and challenge phases of ACD and outlines how this translates into distinct clinical manifestations of rapid-onset versus delayed-type ACD. Furthermore, we highlight the reported limitations of the current therapeutics available, underscoring the need for tailored treatment strategies targeting both classical delayed-type and rapid-onset ACD.
2. The Sensitization Phase and Development of CD8+ TRM Cells
During sensitization, the skin is exposed to a contact allergen, which most commonly is a small (<500 Dalton) electrophilic chemical that readily penetrates the cornified barrier of the epidermis and thus come into contact with live keratinocytes and immune cells [1,30]. Once in the skin, the electrophilic properties of the allergen promote the formation of covalent bonds with the nucleophilic amino acids present in most skin proteins [30]. This results in the generation of highly stable contact allergen–self protein complexes, also referred to as allergen adducts, of which some can be deposited for months in the skin [31,32]. The allergen adducts are recognized by resident skin cells that react by producing pro-inflammatory cytokines, including interleukin (IL)-1β and tumor necrosis factor (TNF)-α, and reactive oxygen species (ROS) [33,34,35,36,37,38]. This activates antigen presenting cells (APCs) (i.e., Langerhans cells and dermal dendritic cells (dDCs)) [38,39,40] which migrate to the local draining lymph nodes (dLNs), where they activate allergen-specific naïve T cells through the following: (i) the presentation of allergen adduct-containing major histocompatibility complexes (MHC I and II) recognized by T cell receptors (TCRs) on the T cells [41,42]; (ii) the expression of the co-stimulatory molecules CD80 and CD86 engaging the CD28 co-receptor on the T cells [43]; and (iii) the release of polarizing cytokines that drive T cell subset differentiation [44,45]. After several days of proliferation (5–7 days in mice and 10–15 days in humans), a high number of allergen-specific effector T cells, with the ability to produce effector cytokines and move into the circulation, are generated in the dLNs [1,46,47]. A large fraction of these allergen-specific effector T cells express skin-homing molecules, including C-X-C motif receptor (CXCR) 3, which enable them to migrate to the affected skin site following a chemokine gradient, including the C-X-C motif ligands (CXCL) 9, 10, and 11 [48,49,50,51,52]. In a highly targeted manner, the T cells then clear keratinocytes presenting allergen adduct-expressing MHC molecules, whereafter most of the effector T cells undergo apoptosis. However, subsets of allergen-specific T cells develop into memory T cells that persist in the skin, circulation, and secondary lymphoid organs [1] (Figure 1).
3. The Role of Neutrophils in the Sensitization Phase
Neutrophils have been described to play an important role during sensitization and challenge to contact allergens. During sensitization, Weber et al. (2015) [21] showed that neutrophils were required for contact allergen-induced release of cytokines, the migration of DCs to the dLNs, and the priming of allergen-specific T cells. The early recruitment of neutrophils to the skin following initial allergen exposure was essential for directing the subsequent infiltration of effector CD8^+^ T cells to the skin [21]. The central role of neutrophils in the sensitization phase is supported by the observation that myeloperoxidase (MPO), a neutrophil-specific mediator of cell damage, is required for the release of IL-1β during sensitization [53]. Neutrophils direct T cells into the skin following exposure to DNFB through expression of FasL and perforin [23] and the-release of the T cell recruiting chemokine interferon gamma (IFN-γ)-induced protein 10 (CXCL10/IP-10) [54]. The rapid entry of neutrophils 3–6 h after exposure to the contact allergen during the sensitization phase depends on IL-1 receptor (IL-1R)-mediated activation of mast cells, dDCs [21], and IL-17-producing γδ T cells in the dermis [55]. Accordingly, IL-1R^−^/^−^ mice exhibit reduced neutrophil and CD8^+^ T cell infiltration in skin exposed to DNFB [56]. Collectively, these findings suggests that infiltration of neutrophils in skin exposed to contact allergens is required to enable subsequent entry of allergen-specific effector T cells during the sensitization phase (Figure 1).
Another important aspect of neutrophil involvement in ACD is the feature of pruritus that often follows after exposure to contact allergens [1]. Transient receptor potential (TRP) ion channels expressed by sensory neurons in the skin enhance ACD responses by stimulating itch and scratching behavior following exposure to contact allergens [57]. The mechanical stress from scratching triggers elevated levels of cytokines (IL-1β, IL-6, TNF-α) and chemokines (CXCL1, CXCL2, CXCL5), leading to increased neutrophil infiltration in the skin [58]. In addition, neutrophil-derived leukotriene B4 (LTB4), released in response to mechanical skin stress (e.g., scratching or tape stripping), further promotes neutrophil recruitment [59]. Notably, both scratching behavior and LTB4-driven neutrophil accumulation were found to be essential for ACD induction [59]. Neutrophil-induced oxidative stress significantly enhances local release of the pruritic mediator IL-33 in the skin, suggesting that neutrophils may also act as a trigger or amplifier of scratching behavior following exposure to contact allergens [60] (Figure 1).
4. The Function of Memory CD8+ T Cells and Neutrophils During the Challenge Phase
Whereas the sensitization phase is typically asymptomatic, the challenge phase elicits the classical clinical symptoms of ACD. Importantly, the dynamics of this phase differ depending on whether the skin has previously been exposed to the contact allergen (Figure 2 and Table 1). Memory T cells are central to the response in both contexts; however, allergen-experienced skin uniquely harbors allergen-specific CD8^+^ T_RM_ cells, whereas allergen-naïve skin does not.
In 1963, Arnason and Waksman were the first to report the existence of local memory to contact allergens at skin sites previously exposed to the given contact allergen, a phenomenon they termed “retest reactions” [61]. This sparked a debate about whether local immunological memory may develop in the skin [62,63]. Yet the critical role of CD8^+^ T_RM_ cells in local immunological memory was not demonstrated until much later. A pivotal study by Gebhardt et al. (2009) identified skin CD8^+^ T_RM_ cells in herpes simplex virus-infected skin that facilitated local protection against reinfections [64]. Later, Gaide et al. (2015) showed that CD8^+^ T_RM_ cells developed and persisted locally in skin exposed to the contact allergen dinitrofluorobenzene (DNFB) [65]. By use of a parabiotic mouse model, they further showed that the presence of skin-resident CD8^+^ T_RM_ cells correlated with an enhanced, rapid challenge response that peaked approximately 24 h after exposure to the allergen. Accordingly, Gadsbøll et al. (2020) showed that the number of epidermal CD8^+^ T_RM_ cells increases with repeated exposures to DNFB and correlates with the magnitude of the challenge response [66].
During the first 24 h after exposure of allergen-naïve skin to the contact allergen, only minimal inflammation is observed. Local innate immune cells recognize the allergen and initiate the processes described in Section 3. Compared to sensitization, the onset of inflammation in allergen-naïve skin is somewhat accelerated due to the presence of allergen-specific CD8^+^ central memory (T_CM_) cells in the draining lymph nodes (dLNs), which proliferate rapidly upon arrival of mature APCs, and the presence of allergen-specific CD8^+^ effector memory (T_EM_) cells in the circulation [65,67,68]. These memory populations infiltrate the challenged skin within 2–4 days, thereby facilitating the classical type IV hypersensitivity response that typically peaks 48–72 h after challenge at an allergen-naïve skin site [4,13,69]. Following resolution, CD8^+^ T_RM_ cells are established at the site, rendering the previously allergen-naïve skin allergen-experienced.
In contrast to allergen-naïve skin, re-exposure of allergen-experienced skin to the sensitizing allergen, as typically occurs in daily life and in some occupational settings, triggers a more rapid and pronounced inflammatory response. In this condition, the epidermis already harbors allergen-specific CD8^+^ T_RM_ cells strategically positioned to respond immediately once the allergen penetrates the skin barrier [7,13,66,70]. Within a few hours, the T_RM_ cells initiate rapid-onset ACD by releasing pro-inflammatory cytokines, most prominently IFN-γ and IL-17 [7,71]. Mechanistically, IFN-γ upregulates MHC I/II and Fas expression on keratinocytes [72,73] and induces secretion of IL-1β, TNF-α, and ROS [71,74], while IL-17, in synergy with IFN-γ, serves as a key upstream driver of neutrophil recruitment [71,75,76]. The critical role of IL-17-producing T cells during the challenge phase has been demonstrated in TCR δ^−^/^−^ and IL-17R^−^/^−^ mice, both of which fail to recruit neutrophils upon DNFB challenge, in contrast to IFN-γR^−^/^−^ mice [55,71]. Accordingly, IL-17 expression and neutrophil accumulation are prominent features of the challenge reaction in nickel-experienced skin of patients allergic to nickel [7,16].
Consistent with these findings, activated T_RM_ cells induce rapid production of neutrophil-recruiting chemokines, including CXCL1 and CXCL2, that drive massive neutrophil infiltration and subsequent tissue damage [13,14,15,16,25,75,76,77]. Both chemokines act through CXCR2, as evidenced by markedly reduced neutrophil infiltration in CXCR2^−^/^−^ mice [22] and following pharmacological inhibition with the CXCR2 antagonist Reparixin [13]. The essential contribution of neutrophils in rapid-onset ACD has further been demonstrated by antibody-mediated depletion of neutrophils [13]. Moreover, neutrophil extracellular trap (NETosis) formation in response to DNFB challenge has recently been identified as an additional effector mechanism amplifying tissue inflammation [78], highlighting the pathological function of neutrophils in the challenge phase.
Whereas the classical type IV response in allergen-naïve skin peaks 48–72 h after challenge, inflammation in allergen-experienced skin has already started to resolve by this time, as skin-infiltrating neutrophils quickly undergo cell death [13]. In parallel, CD8^+^ T_EM_ cells start to infiltrate the site, further promoting the expansion of allergen-specific epidermal CD8^+^ T_RM_ cells that reinforce long-term reactivity to the contact allergen [13,66]. jcm-14-07175-t001_Table 1Table 1Differences between allergen-naïve and -experienced skin upon challenge.FeatureChallenge with Contact Allergen on: Allergen-Naïve SkinAllergen-Experienced SkinRefs.Resulting skin reactionClassical delayed-type (type IV) hypersensitivity reaction. Typically seen when patch testing patients.Rapid-onset and exacerbated reaction. Typically seen in daily life situations where patients are re-exposed at the same skin site repeatedly.[5,6,7,13,15]Presence of allergen-specific T_RM_ cellsAbsent.CD8^+^ T_RM_ cells enriched in epidermis.[7,13,15]KineticsDelayed-type response (peaks ~48–72 h post challenge).Rapid-onset response (<24 h post challenge).[5,6,7,13,15]Memory T cell subsets involved in inflammatory responseCirculating and effector memory T cell subsets (T_CM_ and T_EM_ subsets).CD8^+^ T_RM_ cells (predominantly Tc1 and Tc17 subsets).[7,13,15,79]Cytokines/molecules involvedIL-1β, IL-12, and CXCL10 (late induction); IFN-γ, IL-17, and TNF-α (produced after T cell recruitment).IL-1β, IFN-γ, IL-17, TNF-α, and granzyme B, perforin (produced locally and early); CXCL1 and CXCL2 (recruitment of neutrophils).[7,13,15,22,36,44,49,50,80]
5. Mediators of Long-Term Persistence of Skin-Resident CD8+ TRM Cells
A key feature of T_RM_ cells is their ability to adapt to and persist in the dynamic and energy-constrained environment found in peripheral tissues. In the epidermis, allergen-specific CD8^+^ T_RM_ cells compete with each other and with other T cell subsets for metabolites, survival signals, and the limited anatomical space [66,81,82,83]. IL-15 and transforming growth factor (TGF)-β are essential for the development of CD8^+^ T_RM_ cells [83,84]. IL-15 is constitutively produced by keratinocytes in the hair follicles and appears to play a crucial role in supporting CD8^+^ T_RM_ cell survival in the epidermis [82]. Accordingly, IL-15 signaling has been implicated in CD8^+^ T_RM_ cell-mediated pathology in several autoimmune skin diseases [85], and blockade of the IL-15 receptor has been proposed for treatment of vitiligo [86] and alopecia areata [87], two skin conditions that are driven by pathologic CD8^+^ T_RM_ cells. More recently, TGF-β-induced downregulation of sphingosine-1-phosphate receptor 5 (S1PR5) expression in CD8^+^ T_RM_ cells has been identified as a key mechanism promoting the long-term persistence of CD8^+^ T_RM_ cells in the skin [88]. In contrast, IL-4 impairs CD8^+^ T_RM_ development by inhibiting TGF-β-induced expression of CD103 [89]. Together, these data support that different cytokines, transcription factors, and surface receptors involved in CD8^+^ T_RM_ cell development and persistence could be targets in future treatment of rapid-onset ACD and other CD8^+^ T_RM_ cell-driven skin diseases.
The role of continuous antigen presentation in the long-term persistence of skin CD8^+^ T_RM_ cells has been a subject of debate [31,90,91,92,93]. Most current evidence suggests that while local inflammatory cues can drive effector CD8^+^ T cells to migrate into the skin and transiently adopt a T_RM_ phenotype in the absence of cognate antigen, their long-term persistence depends on the continuous presence of allergen adducts and TCR signaling [31,91,92,93]. It has further been shown that CD8^+^ T_RM_ cells tend to localize near epidermal allergen adducts and that the number and diversity of T cells decline in parallel with decreasing levels of allergen adducts in the epidermis over time [31,70]. Importantly, not all contact allergens are capable of supporting long-term T_RM_ survival [31], which aligns with findings showing that only some contact allergens can propagate the exacerbated neutrophil-mediated reactions observed in rapid-onset ACD [15]. This suggests that the half-life of allergen adducts in the skin influences the persistence of CD8^+^ T_RM_ cells.
6. Potential Therapeutic Targets Against Rapid-Onset ACD
Currently, there is no curative treatment for individuals who have developed contact allergy. Management of ACD primarily focuses on allergen avoidance strategies and topical or systemic corticosteroids to alleviate acute flare-ups [1,94]. Studies in mice have demonstrated that corticosteroid treatments have limited or negligible effects on the development and survival of allergen-specific CD8^+^ T_RM_ cells [95]. There is no current treatment addressing the underlying specific immune response behind rapid-onset nor delayed-type ACD. The recent advances in understanding the immunopathology of rapid-onset ACD have uncovered several promising therapeutic targets aimed at mitigating the immune responses driven by allergen-specific CD8^+^ T_RM_ cells and their recruitment of neutrophils. Here, we wish to review potential therapeutic strategies with a particular focus on targeting rapid-onset ACD. While the following therapeutic approaches are mechanistically intriguing, their clinical applicability in treating ACD remains largely uncertain. Each of the proposed strategies presents specific limitations and potential side effects. A summary of these potential therapeutic strategies and their limitations is provided in Table 2.
6.1. Targeting the IL-1R-CXCR2 Neutrophil Axis
IL-1β acts as a local danger signal during sensitization, and IL-1α drives dermal macrophage production of CXCL2, promoting perivascular immune-cell cluster formation and augmenting CD8^+^ T cell activation and expansion in the skin [33,96]. Accordingly, preclinical mouse studies indicate that blockade of IL-1R [33,97] or CXCR2 [13,22] reduces neutrophil recruitment and downstream T cell-dependent pathology and therefore represents an attractive therapeutic strategy.
However, translation of these therapies to humans is at an early stage. An interventional trial testing IL-1 blockade (Anakinra) in ACD is registered (NCT05498467), and CXCR2 antagonists have been evaluated in several clinical trials against neutrophil-driven implications (for example COPD/bronchiectasis and oncology studies) [98,99,100,101,102], but, to our knowledge, not yet in randomized trials for ACD. Importantly, CXCR2 inhibitors produce reversible reductions in the number of circulating and tissue neutrophils within 24 h in humans [99], demonstrating the pharmacodynamic feasibility of targeting this pathway; however, the clinical efficacy and safety of their use in ACD are still not clear [98,99,100,101,102]. A practical limitation for IL-1R–CXCR2 targeting in rapid-onset ACD is the need for near-immediate administration following allergen re-exposure to prevent neutrophil extravasation. Thus, timing of treatment may limit practical applicability and argues for exploring short-acting topical/locally delivered formulations or rapid-access treatment pathways in future trials for ACD. Moreover, as neutrophils are critical for first-line host defense and wound healing, therapeutic inhibition of neutrophil recruitment may increase susceptibility to infections and impair tissue repair, as reported in clinical studies of systemic CXCR2 blockade [101,102].
6.2. Neutralization Inflammatory Cytokines (IL-17/IFN-γ)
CD8^+^ T_RM_ cells contribute to early neutrophil recruitment in rapid-onset ACD by secreting IL-17 and IFN-γ, which in turn induce the release of CXCL1 and CXCL2 [7,13,16,55,71]. In preclinical models, blockade of IL-17 or IFN-γ signaling has demonstrated efficacy, and several monoclonal antibodies targeting these cytokines, such as Secukinumab (anti-IL-17) [103] and Emapalumab (anti-IFN-γ), exist [104]. Furthermore, neutralization of IL-17 has shown strong efficacy in the treatment of psoriasis [105,106,107]. Given the proposed role of IL-17 in neutrophil recruitment during rapid-onset ACD [7,13,16], it indicates that IL-17 blockade may also be beneficial for ACD patients.
Yet a trial of Secukinumab in nickel-allergic patients demonstrated little-to-no clinical effect after patch testing [108], suggesting that IL-17 is not a major driver of the classical type IV response to nickel, which of course does not rule out that IL-17 plays an important role in rapid-onset ACD. However, a key concern especially with systemic IL-17 blockade is safety as multiple clinical trials (e.g., in psoriasis) showed a significantly increased risk of mucocutaneous candidiasis (oral, cutaneous, and oropharyngeal) with IL-17 inhibitors compared with placebo or non-IL-17 biologics [109,110]. Again, short-acting topical/locally delivered formulations would be preferrable.
6.3. JAK Inhibitors
IL-15, IL-17, and IFN-γ all signal through pathways involving Janus kinases (JAKs). JAK inhibitors, such as Tofacitinib and Ruxolitinib, have shown promise in the treatment of CD8^+^ T_RM_-driven inflammatory skin disorders, including vitiligo [111] and alopecia areata [112,113], and both systemic agents (Abrocitinib, Upadacitinib, Baricitinib) and topical Ruxolitinib cream are approved for the treatment of atopic dermatitis [111,114]. Their ability to suppress cytokine signaling downstream of IFN-γ, IL-4, and IL-17 makes JAK inhibitors attractive candidates for ACD, especially when applied topically, as systemic JAK inhibition carries recognized risks, including thromboembolic events, cytopenia, and serious infections [115,116]. Topical formulations may reduce some of these risks, but their efficacy and safety in ACD remain largely unexplored. Considering the efficacy shown in other local and CD8^+^ T_RM_-driven skin diseases, topically applied JAK inhibitors represent a promising treatment strategy for rapid-onset ACD.
6.4. Modulation of CD8+ TRM Cell Survival and Function
The persistence of CD8^+^ T_RM_ cells in the epidermis depends on IL-15 and TGF-β signaling [83]. Local inhibition of these pathways, for example via monoclonal antibodies targeting IL-15/CD122 signaling [117] or TGF-β [118], may effectively reduce T_RM_ cell survival. Additionally, recombinant cytokines such as IL-4, which has been shown to counteract CD8^+^ T_RM_ cell survival [89], could offer an alternative strategy to inhibit T_RM_ cell survival. IL-4 downregulates the expression of CD103, CD49a, and TGF-β receptor II on CD8^+^ T cells, thus impairing T_RM_ formation in skin, and in atopic dermatitis lesions, IL-4 expression correlates with reduced T_RM_ formation [89].
Despite encouraging preclinical data, no clinical trials have yet evaluated IL-15, TGF-β, or IL-4-based interventions in ACD. Translating these approaches into ACD is challenging because both IL-15 and TGF-β are pleiotropic cytokines with essential roles in immune homeostasis and tissue repair. Blockade of IL-15 signaling risks impairing antiviral and antitumor immunity, while TGF-β inhibition may cause widespread immune dysregulation and promote autoimmune or inflammatory pathology. Recombinant IL-4, while potentially useful for modulating T_RM_ cell survival, could exacerbate type 2 inflammation and worsen conditions such as atopic dermatitis or asthma. Yet, to our knowledge, specific adverse effects in humans with ACD or other immune skin defects have not been studied. Thus, while modulation of T_RM_ survival through IL-15, TGF-β, or IL-4 pathways represents a conceptual curative strategy for rapid-onset ACD, its clinical feasibility and safety remain uncertain and require cautious investigation in human trials.
6.5. Stimulation of Inhibitory Checkpoint Receptor Signaling
PD-1 is an inhibitory receptor expressed on T cells that, upon engagement with its ligands PD-L1 and PD-L2, downregulates T cell activation and promotes immune tolerance [119]. Experimental models have shown promise in enhancing the expression or signaling of inhibitory checkpoint receptors, e.g., by treatment with PD-1 agonists [120]. This may raise the activation threshold of CD8^+^ T_RM_ cells, thereby suppressing T_RM_-induced inflammation and preventing the development of dermatitis [70]. Similarly, cytotoxic T-lymphocyte-associated protein 4 (CTLA-4), another inhibitory receptor, has been shown to suppress pathogenic T cells in an atopic dermatitis mouse model [121]. However, several limitations and potential risks exist with this approach. Systemic administration of checkpoint agonists may cause profound immunosuppression, increasing the susceptibility to infections and malignancies. However, new drugs that can be topically applied and enhance inhibitory checkpoint receptor signaling might hold promise in the treatment of ACD.
6.6. Targeting Metabolic Pathways
Metabolic reprogramming via the AhR and mTOR signaling pathways has regulated CD8^+^ T_RM_ cell development, function, and survival in preclinical mouse studies [66,122,123,124]. In addition, targeting AhR with modulators such as Tapinarof has been shown to downregulate IL-17 responses in both mice and humans [125]. Thus, modulating these pathways may offer an effective strategy to alter both CD8^+^ T_RM_ cell activation and persistence in the skin. Tapinarof has been proven effective in clinical trials for psoriasis [126,127,128] and atopic dermatitis [129,130], with adverse events reported including folliculitis, nasopharyngitis, upper respiratory tract infection, headache, and contact dermatitis [127]. mTOR inhibitors like rapamycin modulate T cell responses, alleviating autoimmune diseases in mice [131,132] and humans [133]. Systemic use of rapamycin has several well-documented adverse effects including enhanced risk of infections, dyslipidemia, mouth ulcers, and occasionally renal or metabolic abnormalities [133,134], so careful monitoring is required in clinical use. To our knowledge, AhR and mTOR pathway modulation in ACD has not been studied. However, topical targeting of metabolic pathways may be an avenue for ACD therapy, but further studies are needed to establish its safety and efficacy in humans.
6.7. Anti-Pruritic Strategies
As scratching exacerbates skin inflammation and allergen penetration, targeting itch-related pathways, such as TRPA1, IL-33, LTB4, or histamine receptors, could indirectly reduce the severity of ACD by preventing mechanical skin barrier disruption and neutrophil recruitment [57,58,59,60].
While leukotriene receptor antagonists (LTRAs) are clinically used for other pruritic or inflammatory skin conditions [135,136], no trials have specifically tested these in ACD. Similarly, TRPA1 antagonists (Montelukast) have shown efficacy in reducing itch/pain [137], but no dermatology-specific clinical trials have been completed to our knowledge. Biologics targeting IL-33 (e.g., Etokimab, Tozorakimab) have entered testing in atopic dermatitis, with early proof-of-concept data suggesting reduced disease activity and pruritus, including a significant reduction in neutrophil skin infiltrates [138]. Subsequent phase 2 trials have reported mixed efficacy [139]. In contrast, H_1_-antihistamines remain the primary agent of symptomatic anti-pruritic therapy in dermatology, particularly for urticaria and other histamine-dependent pruritic dermatoses [140], although their efficacy in non-histaminergic itch seem limited, suggesting no or minor efficacy in ACD.
Potential adverse effects also need to be considered when targeting itch pathways. H_1_-antihistamines are generally well tolerated, but drowsiness, fatigue, and impaired concentration and memory are reported due to their ability to cross the blood–brain barrier. Yet second-generation antihistamines, including loratadine, seem less likely to cause sedation, but may still have mild central nervous system effects [141,142]. IL-33 antagonists have shown a favorable safety profile in clinical trials for atopic dermatitis, although a single adverse event of venous thrombosis was reported [139]. However, larger studies are needed to fully assess their long-term safety. TRPA1 antagonists have demonstrated safety in early-phase clinical trials, with no significant adverse events reported [143]. Nonetheless, due to the broad expression of TRPA1 in various tissues, potential off-target effects warrant further investigation.
6.8. Redox Modulation and Allergen Metabolism
Variations in allergen persistence seem to influence the ability of CD8^+^ T_RM_ cells to survive [31,69,144]. The Nrf2 transcription factor pathway plays a key role in the detection and detoxification of cytosolic allergen adducts [28,145]. Thus, enhancing Keap1-Nrf2 signaling to regulate the skin’s redox balance and facilitate allergen clearance could represent a strategy to reduce allergen adducts and ROS-driven skin inflammation. In this context, antioxidants, including sulforaphane and curcumin, are potent activators of the Nrf2 pathway that have been shown to modulate oxidative stress and inflammatory responses [146]. While preclinical studies support these effects, clinical trials in ACD are lacking, and systemic administration of Nrf2 activators may carry off-target effects, again highlighting the need for further research to establish efficacy for ACD and safety in humans. jcm-14-07175-t002_Table 2Table 2Potential therapeutic strategies for rapid-onset ACD.Therapeutic StrategyMechanismLimitationsReported/Potential Side EffectsRefs.Anti-IL-1and anti-IL-1R blockade (e.g., Anakinra, CXCR2 inhibitors)Reduces neutrophil recruitment and downstream CD8^+^ T_RM_ activationRequires near-immediate administration after exposure; limited data in ACDNeutropenia, increased infection risk; may transiently impair host defense[13,22,33,96,97,98,99,100,101,102]Cytokine neutralization (anti-IL-17/anti-IFN-γ) (e.g., Secukinumab, Emapalumab)Blocks CD8^+^ T_RM_-induced inflammation and downstream release of neutrophil-recruiting chemokinesSecukinumab showed limited efficacy in a nickel allergy trial; unknown efficacy against rapid-onset ACDIL-17 inhibitors: increased risk of mucocutaneous candidiasis; IFN-γ blockade: increased infection risk[7,71,103,104,105,106,107,108,109,110]JAK inhibitors (e.g., Tofacitinib, Ruxolitinib, Abrocitinib, Upadacitinib, Baricitinib)Broad inhibition of cytokine signaling (IL-15, IL-17, IFN-γ, IL-4) and inflammationSystemic use limited by safety concerns; efficacy in ACD unknown; topical efficacy in ACD is unexplored Systemic: thromboembolism, cytopenia, and serious infections; topical: local irritation and unknown long-term safety[111,112,113,114,115,116]CD8^+^ T_RM_ modulation (e.g., anti–IL-15, anti–TGF-β, IL-4 administration)Reduces CD8^+^ T_RM_ survival and persistenceNo ACD trials; IL-15 and TGF-β are pleiotropic; IL-4 may worsen or cause development of type 2 autoimmunity.IL-15 blockade may impair antiviral/antitumor immunity; TGF-β blockade increases risk of autoimmunity/inflammation; IL-4 may exacerbate AD/asthma[83,89,117,118,147]Checkpoint receptor agonists (e.g., PD-1, CTLA-4 agonists)Inhibit T cell activation and dampen T_RM_-driven inflammationNo clinical trials in skin disease; systemic immunosuppression risk; topical formulations not availableIncreased infection and malignancy risk with systemic administration[70,119,120,121]Metabolic pathway modulation (AhR modulators e.g., Tapinarof; mTOR inhibitors, e.g., rapamycin)Alters T_RM_ activation/inflammation and survival; Tapinarof is shown to reduce IL-17 expressionNo studies in ACD; paradoxical of contact dermatitis with Tapinarof; systemic rapamycin toxicityTapinarof: folliculitis, nasopharyngitis, and contact dermatitis; rapamycin: infections, dyslipidemia, mouth ulcers, and renal/metabolic toxicity[66,122,123,124,125,126,127,128,129,130,131,132,133,134,148] Anti-pruritic strategies (e.g., H_1_-antihistamines, TRPA1 antagonists, LTRAs, IL-33 blockade)Reduce scratching-induced barrier damage and neutrophil infiltrationLimited efficacy in non-histaminergic itch; mixed efficacy in AD (IL-33); no ACD trialsH_1_-antihistamines: sedation, fatigue, and cognitive impairment; IL-33 blockade: rare thrombosis; TRPA1 antagonists: possible off-target effects; LTRAs: safe in asthma, but untested in ACD[57,58,59,60,135,136,137,138,139,140,141,142,143]Redox modulation/allergen metabolism (e.g., Nrf2 activators: sulforaphane, curcumin)Enhances detoxification of allergen adducts and reduces oxidative stressOnly preclinical data; systemic administration may cause off-target effectsSafety in ACD not established [28,31,69,144,145,146]
7. Conclusions and Perspectives
Importantly, by distinguishing rapid-onset ACD from the classical delayed-type reaction, this review highlights a clinically distinct manifestation that is highly relevant for patients who experience sudden flare-ups in daily life. Recognizing this difference is crucial for accurate diagnosis and the development of therapeutic strategies tailored to rapid-onset ACD, which is currently underrepresented in the literature. Collectively, this review extends current knowledge by providing a comparative perspective on rapid-onset and delayed-type ACD. Whereas delayed-type ACD is dominated by T cell responses peaking after 48–72 h, rapid-onset ACD involves immediate CD8^+^ T_RM_ cell activation and neutrophil-driven inflammation, leading to acute clinical flare-ups. This contrast not only clarifies discrepancies between experimental models, patch testing, and daily patient experiences, but also carries translational implications for therapy, emphasizing the need to specifically target mechanisms unique to rapid-onset disease. The recognition of CD8^+^ T_RM_ cells and neutrophils as central players in rapid-onset ACD has opened new avenues for therapeutic interventions regarding ACD. Approaches aimed at modulating T_RM_ cell survival, function, and activation thresholds including targeting central cytokine pathways, altering the metabolic and transcriptional environment, and disrupting neutrophil recruitment are proposed in this review.
While the therapeutic strategies discussed here highlight the potential for mechanism-based treatment of ACD, their translation to clinical practice faces significant challenges. Most remain untested in ACD patients, and safety profiles may be unfavorable for a condition often managed effectively by allergen avoidance. The accelerated kinetics of rapid-onset ACD further complicates intervention, as patients are frequently unaware of recent allergen exposure at the time symptoms develop. Therefore, avoidance strategies will likely continue to be the main strategy for contact allergic patients. Nonetheless, for individuals at a high risk of repeated exposure, such as those in certain occupations, a novel, fast-acting, local/topical, and easy-to-administer therapy capable of halting or attenuating rapid-onset ACD could be highly beneficial. Such interventions could reduce acute inflammation, prevent disruption of skin barrier function, and limit the downstream recruitment of pathogenic immune cells, thereby preserving work ability and lowering the current cost for society. Future studies should focus on evaluating these strategies in well-designed preclinical and clinical trials, with careful attention paid to their safety and feasibility and the unique kinetics of rapid-onset versus delayed-type ACD.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Scheinman P.L. Vocanson M. Thyssen J.P. Johansen J.D. Nixon R.L. Dear K. Botto N.C. Morot J. Goldminz A.M. Contact Dermatitis Nat. Rev. Dis. Prim.202173810.1038/s 41572-021-00271-434045488 · doi ↗ · pubmed ↗
- 2Svensson A. Ofenloch R.F. Bruze M. Naldi L. Cazzaniga S. Elsner P. Goncalo M. Schuttelaar M.-L.A. Diepgen T.L. Prevalence of Skin Disease in a Population-Based Sample of Adults from Five European Countries Br. J. Dermatol.20181781111111810.1111/bjd.1624829247509 · doi ↗ · pubmed ↗
- 3Tramontana M. Hansel K. Bianchi L. Sensini C. Malatesta N. Stingeni L. Advancing the Understanding of Allergic Contact Dermatitis: From Pathophysiology to Novel Therapeutic Approaches Front. Med.202310118428910.3389/fmed.2023.1184289 PMC 1023992837283623 · doi ↗ · pubmed ↗
- 4Johansen J.D. Aalto-Korte K. Agner T. Andersen K.E. Bircher A. Bruze M. CannavõA. Giménez-Arnau A. Gonçalo M. Goossens A. European Society of Contact Dermatitis Guideline for Diagnostic Patch Testing—Recommendations on Best Practice Contact Dermat.20157319522110.1111/cod.1243226179009 · doi ↗ · pubmed ↗
- 5Ahlström M.G. MennéT. Thyssen J.P. Johansen J.D. Nickel Allergy in a Danish Population 25 Years after the First Nickel Regulation Contact Dermat.20177632533210.1111/cod.1278228386993 · doi ↗ · pubmed ↗
- 6Hindsén M. Bruze M. Christensen O.B. The Significance of Previous Allergic Contact Dermatitis for Elicitation of Delayed Hypersensitivity to Nickel Contact Dermat.19973710110610.1111/j.1600-0536.1997.tb 00312.x 9330814 · doi ↗ · pubmed ↗
- 7Schmidt J.D. Ahlström M.G. Johansen J.D. Dyring-Andersen B. Agerbeck C. Nielsen M.M. Poulsen S.S. Woetmann A. Ødum N. Thomsen A.R. Rapid Allergen-induced Interleukin-17 and Interferon-γ Secretion by Skin-resident Memory CD 8 + T Cells Contact Dermat.20177621822710.1111/cod.1271527873334 · doi ↗ · pubmed ↗
- 8Johansen J.D. Bonefeld C.M. Schwensen J.F.B. Thyssen J.P. Uter W. Novel Insights into Contact Dermatitis J. Allergy Clin. Immunol.20221491162117110.1016/j.jaci.2022.02.00235183605 · doi ↗ · pubmed ↗