RNAi of vATPasea Affects Survival and Larval-Pupal Development in Plutella xylostella
Xuetao Yu, Jinhua Luo, Lin Lu, Li Zhu, Siyuan Wang, Kang Yang, Xia Wan, Yuhua Wu, Boboev Akmal, Gang Wu, Xiaohong Yan, Chenhui Shen

TL;DR
This study shows that silencing a specific gene in diamondback moths using RNAi reduces their survival and development, suggesting a new pest control method.
Contribution
The study identifies PxvATPasea as a potential RNAi target gene in Plutella xylostella with dose-dependent effects.
Findings
Injecting dsPxvATPasea reduced mRNA levels and caused larval mortality in a dose-dependent manner.
PxvATPasea silencing significantly decreased larval fresh weight and pupation rates.
The gene is abundantly expressed in hindgut and Malpighian tubules of the diamondback moth.
Abstract
Relatively little attention has been given to genes encoding V0 subcomplex subunits. In the study, we identified PxvATPasea specifically and evaluated expression profiles of PxvATPasea across developmental stages and among tissues. By injecting two doses of dsPxvATPasea (800 ng or 1200 ng) in Plutella xylostella, we showed that PxvATPasea is a potential molecular target gene and RNAi efficiencies worked in a dose-dependent way. The diamondback moth, Plutella xylostella, is a major pest of brassica vegetables and oilseed crops, posing a serious threat to China’s grain and oil production. RNA interference (RNAi) has been developed as an efficient strategy to control pests. In this study, the effects of RNAi on P. xylostella were evaluated by injecting two doses of synthesized dsPxvATPasea. The transcripts of PxvATPasea were widely transcribed during different developmental stages from…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
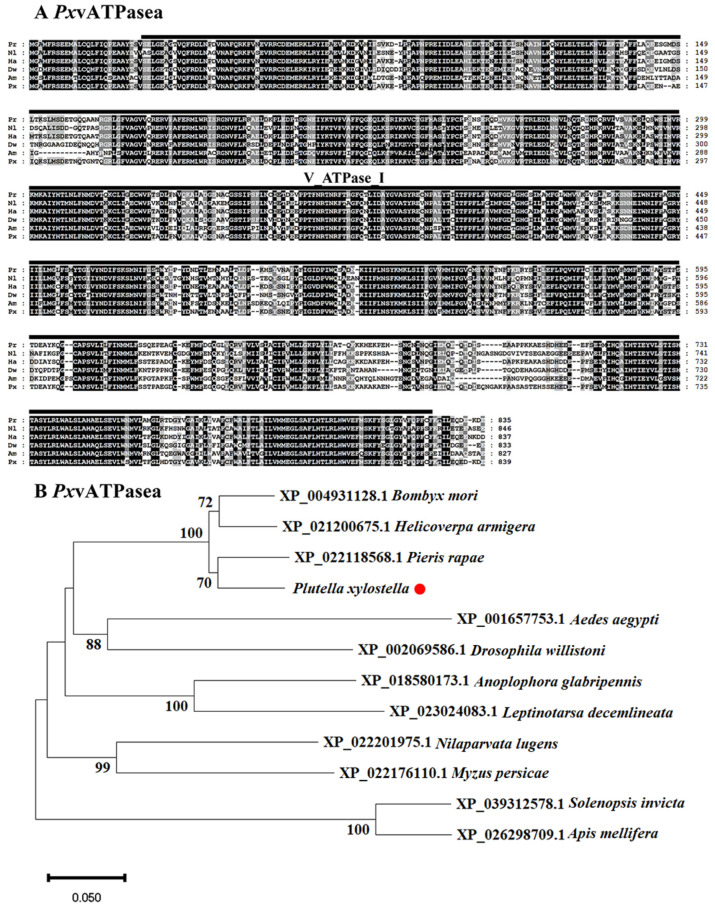 Figure 1
Figure 1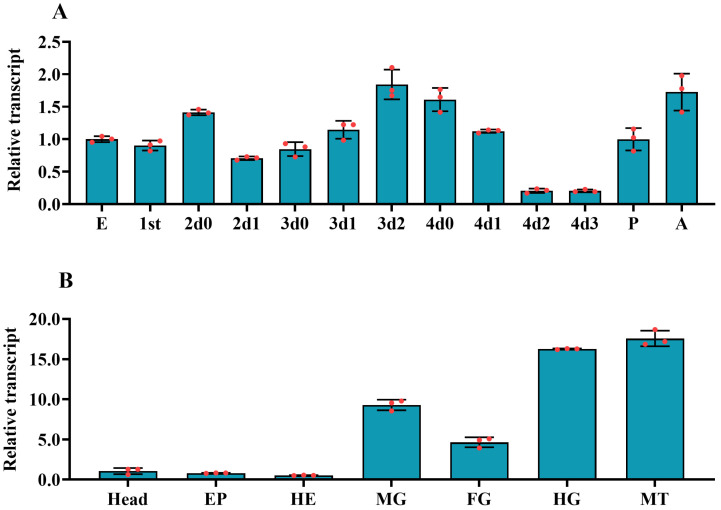 Figure 2
Figure 2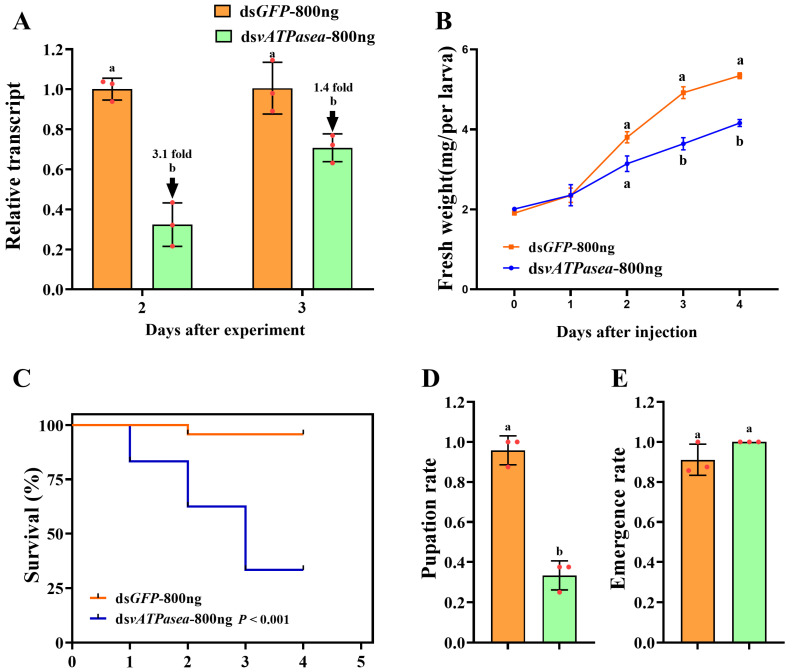 Figure 3
Figure 3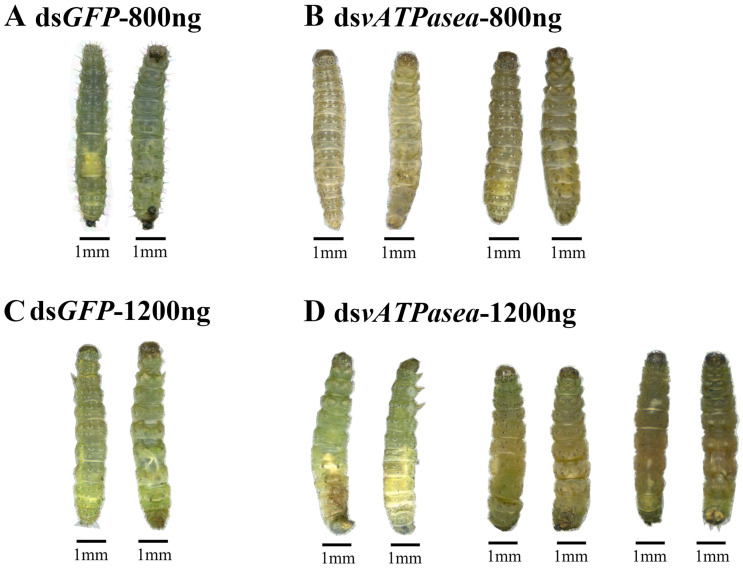 Figure 4
Figure 4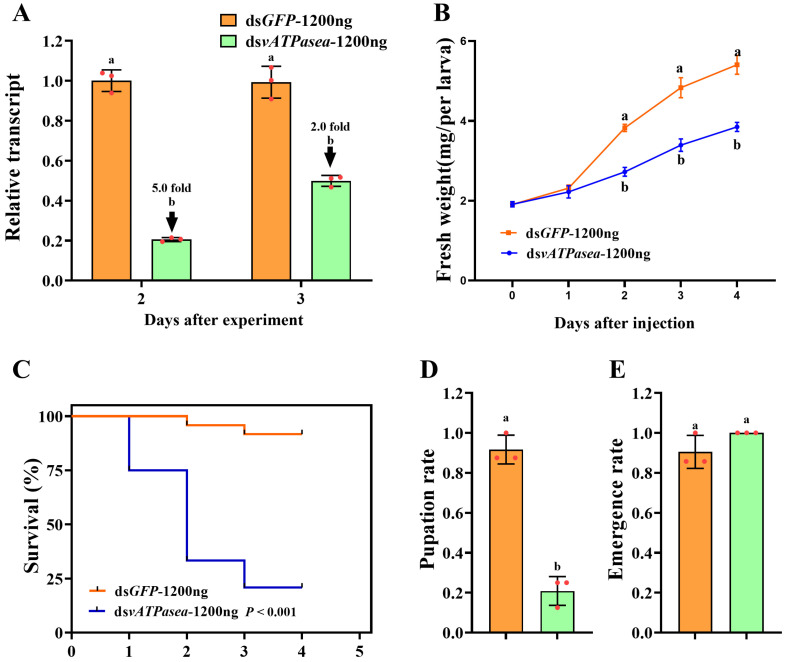 Figure 5
Figure 5- —National Key Research and Development Program of China
- —Outstanding Young Talent Cultivation Project of Oil Crops Research Institute of Chinese Academy of Agricultural Sciences
- —Innovation Project of Chinese Academy of Agricultural Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect Resistance and Genetics · CRISPR and Genetic Engineering · Advanced biosensing and bioanalysis techniques
1. Introduction
The diamondback moth, Plutella xylostella, is one of the primary lepidopteran pests attacking brassica vegetables and oilseed crops, causing monetary losses of around US $4–5 billion worldwide [1,2]. At present, chemical pesticides are applied mainly to manage this pest, such as chlorantraniliprole, chlorpyrifos, and indoxacarb [3]. However, owing to the rapid development of resistance, it is more and more difficult to control its field populations based on insecticides [2]. Thus, it seems much more urgent and necessary to develop an efficient strategy to manage the population of P. xylostella.
RNA interference (RNAi) is an efficient molecular mechanism for suppressing the expression of post-transcriptional target genes triggered by double-stranded RNA (dsRNA), which can be used to develop an environmentally friendly and reduced-risk insect pest management approach [4,5,6,7]. In recent years, numerous research have demonstrated that RNAi-based integrated pest management works well in various insects [7,8,9,10,11,12,13]. For example, use plant-mediated RNA interference strategy to control Frankliniella occidentalis [9] and Apolygus lucorum [10]; RNAi silencing protease genes can affect mortality in Helicoverpa armigera [12]; continuous ingestion of dsLdRan can kill the larvae and adults of Leptinotarsa decemlineata [7]; knockdown of HvUAP impairs larval growth in Henosepilachna vigintioctopunctata [13]. Currently, silencing target genes PxPiwi [14] and PxTH [15] in P. xylostella affected larval development. Thus, it seems practicable to apply RNAi-based methods to manage population of P. xylostella.
Vacuolar-type H^+^-ATPase (vATPase) is a proton translocating pump localized in the internal membranes of eukaryotes, functioning by hydrolyzing ATP to ADP and phosphate to pump protons across membranes [16,17,18,19]. The structures of vATPases are evolutionarily conserved in eukaryotes, consisting of two subcomplexes, V_1_ and V_0_ [20,21]. The V_1_ subcomplex contains eight different subunits (A through H), with stoichiometry of A_3_B_3_CDE_3_FG_3_H in Manduca sexta and Saccharomyces cerevisiae, which is related to ATP hydrolysis. The V_0_ subcomplex consists of five different subunits (a_1_d_1_c_4–5_c′_1_c″1) and accessory proteins, which can drive transmembrane proton movement [21,22,23,24,25,26].
Currently, relatively little attention has been given to genes encoding V_0_ subcomplex subunits, and only two research have verified that depletion of vATPasea was lethal [27,28]. In Drosophila melanogaster, P-element insertions in vha100-2 are lethal [27]. In H. vigintioctopunctata, knockdown of HvvATPasea in the third-instar larvae caused 100% larval lethality [28]. It is vital to determine whether silencing genes encoding V_0_ subcomplex subunits results in severe defective phenotypes in other insects. In addition, compared to Bt-transgenic Brassica napus, expression of CHS1 hpRNA in transgenic B. napus shortened the mortality time in P. xylostella [1]. Therefore, RNAi-based control of Plutella xylostella populations by feeding them dsRNA targeting PxvATPasea holds promise for future development.
In the present study, PxvATPasea was chosen as the target gene to assess the possibility of management methods for P. xylostella. Specifically, we (i) cloned, aligned, and phylogenetically analyzed the vATPasea gene in P. xylostella; (ii) evaluated expression profiles of PxvATPasea across developmental stages and among tissues; (iii) assessed the RNAi efficiency by measuring the suppression of PxvATPasea expression after treatment with synthesized dsPxvATPasea in vitro; and (iv) examined the effects on larval weight, survival rate, pupation rate, and emergence rate following injection of dsPxvATPasea.
2. Materials and Methods
2.1. Insect Rearing
P. xylostella was collected from B. napus in Haidong City, Qinghai Province, China. The larvae of P. xylostella were fed on fresh brassica leaves at 26 ± 1 °C, 16 h:8 h photoperiod, and 60% ± 5% relative humidity. The adults were reared using 10% honey solution as food.
2.2. Molecular Cloning
The putative PxvATPasea was obtained from the genome and transcriptome data of P. xylostella [29,30]. The correctness of the sequence was proven by polymerase chain reaction (PCR) using primers in Table S1 and Figure S1. The accession number of sequenced cDNA was XP_048479200.1.
Phylogenetic analysis of PxvATPasea and other vATPasea sequences downloaded from NCBI (https://www.ncbi.nlm.nih.gov/, accessed on 18 April 2024) was conducted using MEGA 5.0 software (https://sourceforge.net/projects/mega5/, accessed on 18 April 2024) and the neighbor-joining method, with 1000 bootstrap replications.
2.3. Synthesis of dsRNA Molecules
The two cDNA fragments targeting PxvATPasea and enhanced green fluorescent protein were amplified by PCR using special primers containing the T7 promoter sequence (Table S1, Figure S1). To ensure the specificity and efficacy of the dsRNA, the target sequences by BioEdit 7.0 software were subjected to BLASTN analysis against the P. xylostella transcriptome and evaluated using a web-based prediction tool (https://www.dsrna-engineer.cn, accessed on 18 April 2024) for on-target and off-target sites. Regions with a high density of on-target sites were selected while avoiding potential off-target sequences, thereby excluding any segments with ≥20 bp identical matches that could lead to off-target effects (Figure S2). dsRNA was then synthesized using the MEGAscript T7 High Yield Transcription Kit (Ambion, Austin, TX, USA), according to the manufacturer’s instructions. The reaction mixture contained 2 μL of T7 Express Enzyme Mix (Promega, Beijing, China), 10 μL of RiboMAX™ Express T7 2× Buffer (Promega, Beijing, China), and 1 μg of DNA template and was incubated at 37 °C for 2–6 h, followed by 70 °C for 10 min, and slowly cooled to room temperature (approximately 20 min) for dsRNA annealing. The DNA template and single-stranded RNA were removed by treatment with DNase and RNase A, respectively, and the dsRNA product was purified using a gel extraction kit (Omega Bio-tek, Norcross, GA, USA). The yield of dsRNA was determined by measuring absorbance at 260 nm with a Nanodrop 1000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA), and its integrity was verified by agarose gel electrophoresis. The purified dsRNA was aliquoted and stored at −80 °C until use.
2.4. Introduction of dsRNA
The same method as previously described was used to inject dsRNA [31,32,33]. Briefly, newly molted fourth-instar larvae were selected for microinjection. Borosilicate microcapillaries were pulled using a PC-10 puller (Narishige, Tokyo, Japan). A volume of 200 nL containing one of two dsRNA doses (800 ng or 1200 ng in total, based on references [34]) was delivered into the larval hemolymph using a microinjection system from World Precision Instruments (Sarasota, FL, USA). Larvae injected with dsGFP served as the negative control. A group of eight injected larvae constituted one replicate, and each dsRNA treatment was repeated six times. After injection, larvae were transferred to individual plastic rearing boxes and fed with fresh rape leaves. For sampling, three replicates were collected 2 and 3 days post-injection for qRT-PCR analysis to assess RNAi efficiency. Another three replicates were maintained for a 3-week observation period to monitor defective phenotypes, changes in larval weight, survival rate, pupation rate, and emergence rate.
2.5. Real-Time Quantitative PCR (qRT-PCR)
RNA templates of samples were collected to analyze the temporal expression profiles and tissue transcription patterns. Moreover, for testing the effects of the treatments, total RNA was extracted from treated larvae by the TRIzol reagent (YiFeiXue Tech, Nanjing, China). Each sample contained eight individuals and was repeated three times. Quantitative real-time PCRs (qRT-PCR) were conducted to assess the transcript expression levels according to the previously described method [35]. The qRT-PCR was performed with three biological and technical replicates. The relative mRNA expression levels were analyzed using 2^−ΔΔCt^ method.
2.6. Data Analysis
We used The SPSS Statistics 27 software for Windows (Chicago, IL, USA) for the statistical analyses. The averages (±SE) were used to analyze variance with the Tukey–Kramer test. Survival curves were analyzed using a log-rank test (Mantel–Cox, 95% CI) in GraphPad Prism software version 8.0.
3. Results
3.1. Identification of PxvATPasea
A putative full-length cDNA encoding PxvATPasea was obtained in P. xylostella by mining transcriptome data. The cDNA was comprised of a 2520 bp complete open reading frame encoding 839 amino acid residues (Figure S1).
The sequences of vATPasea proteins in different insects were highly conserved. It contained V_ATPase_I structural domain (Figure 1A). The phylogenetic dendrogram of vATPasea proteins from 12 species was built to assess the evolutionary relationships (Figure 1B). These sequences were from four Lepidoptera, Pieris rapae, H. armigera, Bombyx mori, and P. xylostella; two Hymenoptera, Apis mellifera and Solenopsis Invicta; two Diptera, Aedes aegypti and Drosophila willistoni; two Hemiptera, Nilaparvata lugens and Myzus persicae; and two Coleoptera, Leptinotarsa decemlineata and Anoplophora glabripennis. The unrooted tree revealed that the vATPasea-like proteins of species from the same order were clustered together. Apparently, vATPasea from P. xylostella belonged to Lepidoptera subclade (Figure 1B).
3.2. The Expression Profiles of PxvATPasea
We used qRT-PCR to test the temporal expression pattern of PxvATPasea. The mRNA levels of PxvATPasea were widely transcribed during different developmental stages from embryo (egg) to adult. Its mRNA levels rose to a peak at day 2 of third-instar larval period; however, the lowest PxvATPasea level was detectable at the late fourth-instar larvae (day 2 and day 3). It was clearly observed that the mRNA levels of PxvATPasea regularly aggrandized during ecdysis period (Figure 2A).
The spatial expression profiles of PxvATPasea were evaluated in all tested tissues, including head, foregut, midgut, hindgut, hemolymph, Malpighian tubules, and epidermis. The mRNA levels of PxvATPasea were high in the hindgut and Malpighian tubules, intermediate in the foregut and midgut, and low in the hemolymph, epidermis, and head (Figure 2B).
3.3. Effects of RNAi for 800 ng of dsPxvATPasea at the Fourth-Instar Larvae
To evaluate the effects of knockdown of PxvATPasea in P. xylostella, we silenced PxvATPasea by injection of dsRNA. Introduction of 800 ng dsPxvATPasea in the newly molted fourth-instar larvae significantly decreased the PxvATPasea mRNA level by 3.1 and 1.4 times at 2 d and 3 d, respectively (Figure 3A).
Knockdown of PxvATPasea significantly restrained the development of the larvae, with fresh weights reduced by 26.0% and 22.1% at day 3 and 4, respectively, compared to the dsGFP control (Figure 3B). RNAi of PxvATPasea caused a significant mortality of the larvae. After the bioassay, mortality was 16.7%, 37.5%, and 66.6% at day 2, 3, and 4, respectively, in this treatment (Figure 3C). The remaining 33.4% larvae successfully pupated (Figure 3D). There were no significant differences in emergence rate among the two groups (Figure 3E). The dsPxvATPasea-treated larvae possessed small and misshapen body size. Subsequently, they slowly withered, blackened, and, lastly, died (Figure 4A vs. Figure 4B).
3.4. Impacts on Introduction of 1200 ng dsPxvATPasea at the Fourth-Instar Larvae
Introduction of 1200 ng dsPxvATPasea by microinjection at the newly molted fourth-instar larvae successfully reduced the mRNA level of PxvATPasea by 5.0 and 2.0 times at 2 d and 3 d, respectively, compared to the dsGFP-treated group (Figure 5A).
RNAi of PxvATPasea significantly inhibited the larval growth (Figure 5B), with fresh weights decreased by 28.0%, 30.0%, and 28.8% on day 2, 3, and 4, respectively, compared to the dsGFP-injected group (Figure 5B). In addition, silencing PxvATPasea seriously led to larval lethality, with mortality at 25.0%, 66.6%, and 79.2%, at day 2, 3, and 4, respectively (Figure 5C). The remaining 20.8% larvae successfully pupated (Figure 5D), and the emergence rate showed no significant differences (Figure 5E). The PxvATPasea RNAi larvae failed to molt their cuticle to become pupae. Their body size was small and abnormal, compared with the dsGFP-injected group. Subsequently, these stunting PxvATPasea hypomorphs became withered, dried, and blackened slowly and eventually died (Figure 4C vs. Figure 4D).
4. Discussion
Currently, relatively little attention has been given to genes encoding V0 subcomplex subunits. In the present study, we identified a vATPase subunit a gene in P. xylostella. We uncovered that silencing PxvATPasea affects the survival and growth of larvae.
4.1. RNAi Is a Powerful Tool to Manage P. xylostella
RNAi is one of the most important technologies in the last century [36], which is used as a powerful tool to study the functions of target genes or screen lethal genes for pest management in environmentally friendly and species-specific way [5,37,38,39,40]. Moreover, the development and application of biopesticides based on RNAi is a popular subject in pest management [41]. Up to date, great strides have been made in using genetically modified crops to control virous pests, including A. lucorum [10], L. decemlineata [42], Adelphocoris suturalis [43], Sitobion avenae [44], M. persicae [45], and so on. This evidence has strongly indicated that an RNAi-based strategy is viable to control pests.
The first step is to select suitable candidate genes for the development of RNAi-mediated control methods. Numerous studies have demonstrated that housekeeping genes such as actins [46]; cellular function genes like vacuolar ATPase [47]; energy metabolism genes, including eukaryotic translation initiation factor 5A [48]; and 20E signaling network genes like ecdysone receptors (EcR) [49,50,51] are known to be necessary for the growth of larvae. They can be developed as dsRNA insecticides or transgenic crops expressing dsRNA to control pests.
At present, RNAi of PxPiwi [14] and PxTH [15] in P. xylostella affected larval growth. Therefore, it seems very feasible to apply RNAi technology to control population of this pest.
4.2. PxvATPasea Is a Potential Candidate Gene to Control the Larvae of P. xylostella
In short, our results indicated that dsPxvATPasea is a potential candidate gene to control the larvae of P. xylostella. Firstly, vATPasea is a highly conserved evolutionarily ancient enzyme with the conserved domain of V_ATPase_I (Figure 1), which performs a wide range of functions in eukaryotic species [16,17,18,22,52].
Secondly, PxvATPasea was abundantly transcribed during different developmental stages, from embryo (egg) to adult. The expression of PxvATPasea was high at day 2 of the third-instar larval period and was low in the late fourth-instar larvae. Consistent with our results, the mRNA level of HvvATPasea was high in the third-instar larvae in H. vigintioctopunctata.
Thirdly, vATPases have an important role in nutrient absorption in the gut of various insects [19,53,54]. Our results showed that PxvATPasea was abundantly transcribed in the hindgut and Malpighian tubules in P. xylostella. Likewise, in H. vigintioctopunctata, the expression levels of HvvATPasea were highly expressed in hindgut [28].
Lastly, we uncovered that the introduction of dsPxvATPasea by injecting two different doses of dsRNA at the fourth-instar larvae significantly decreased the mRNA levels of PxvATPasea at different times, severely affected larval development, and killed 66.6% and 79.2% of larvae, respectively, under 800 ng and 1200 ng conditions (Figure 3 and Figure 4). In line with our result, in D. melanogaster, P-element insertions in vha100-2 are lethal [27]; RNAi of HvvATPasea in the third-instar larvae of H. vigintioctopunctata led to 100% larval lethality [28].
Consequently, PxvATPasea is a potential candidate gene to manage by RNAi-based management strategies in P. xylostella.
Current functional studies on insect vATPases have primarily focused on subunits of the V1 complex, while research on V0 complex subunits remains relatively limited. Silencing of vATPase subunit a in the migratory locust (Locusta migratoria) specifically affects the midgut epithelium and causes high mortality, mainly by reducing the number of columnar epithelial cells during midgut development, which subsequently lowers nutrient transport efficiency and leads to reduced body weight [55]. Similarly, RNAi-mediated knockdown of the vATPase-a2 gene in the Sogatella furcifera impacts feeding behavior, resulting in significant changes in mortality, phenotypic defect incidence, and adult eclosion rate [56]. In Lepidoptera, research on the vATPase a subunit remains largely unexplored. Current evidence demonstrates that knockdown of vATPasea in the diamondback moth reduces body weight and causes high mortality, consistent with findings from RNAi targeting vATPasea subunits in other insects. However, the precise mechanisms through which it influences vital physiological processes require further investigation.
4.3. Enhancing RNAi Efficiency by Stabilizing dsRNA in Insects
Multiple factors influence RNAi efficiency, with dsRNA stability being a critical determinant. Naked dsRNA introduced into insects via injection or feeding is susceptible to degradation by nucleases present in saliva, hemolymph, and the gut, which partially accounts for the low RNAi efficiency observed in certain insect species. Particularly in Lepidoptera, nucleases identified as RNAi efficiency-related nucleases (REases) have been recognized [57]. Studies demonstrate that insect hemolymph and gut fluid contain double-stranded ribonucleases (dsRNases), which are considered a major limiting factor for RNAi efficiency due to their impact on dsRNA stability in bodily fluids. In insects, reducing dsRNase activity helps minimize intracellular dsRNA degradation and enhances RNAi efficacy [58,59].
Regarding dsRNA degradation prevention, encapsulating dsRNA with protective materials effectively preserves its integrity. The use of nanoparticle complexes to encapsulate dsRNA represents a novel delivery strategy for RNAi-based applications. These diverse complexes facilitate improved cellular uptake of dsRNA due to their high transduction efficiency and low cytotoxicity, while also reducing the risk of nuclease degradation under environmental conditions, such as temperature variations and medium changes [60]. The most extensively tested nanocarriers include chitosan, liposomes, star polycations (SPcs), layered double hydroxides (LDH), and guanylated polymers (GNP). The control efficacy of different vATPase subunits in S. furcifera was evaluated using star polymer (SPc) nanoparticles via spray-induced and nanoparticle-delivered gene silencing (SI-NDGS), which successfully reduced target mRNA levels and vATPase enzyme activity, while also assessing the environmental safety of nanoparticle-encapsulated dsRNA [56]. Three distinct nanoparticle-encapsulated dsRNA complexes significantly knocked down multiple genes, including VATPase, in Earias vittella [61]. Loading dsRNA of the VATPaseA gene onto layered double hydroxide (LDH) enhanced its environmental stability, leading to mortality in Holotrichia parallela larvae and disruption of their cuticle and midgut structures [62]. Therefore, further research on RNAi approaches targeting dsPxvATPasea will represent a promising strategy for managing diamondback moth populations.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Deng P. Peng Y. Sheng Z. Li W. Liu Y. RN Ai silencing CHS 1 gene shortens the mortality time of Plutella xylostella feeding Bt-transgenic Brassica napus Pest Manag. Sci.2024802610261810.1002/ps.796838252693 · doi ↗ · pubmed ↗
- 2Shehzad M. Bodlah I. Siddiqui J.A. Bodlah M.A. Fareen A.G.E. Islam W. Recent insights into pesticide resistance mechanisms in Plutella xylostella and possible management strategies Environ. Sci. Pollut. Res. Int.202330952969531110.1007/s 11356-023-29271-537606784 · doi ↗ · pubmed ↗
- 3Wang M. Zhu B. Zhang L. Xiao Y. Liang P. Wu K. Influence of seasonal migration on evolution of insecticide resistance in Plutella xylostella Insect Sci.20222949650410.1111/1744-7917.1298734873833 · doi ↗ · pubmed ↗
- 4Christiaens O. Whyard S. Vélez A.M. Smagghe G. Double-stranded RNA technology to control insect pests: Current status and challenges Front. Plant Sci.20201145110.3389/fpls.2020.0045132373146 PMC 7187958 · doi ↗ · pubmed ↗
- 5Liu S. Jaouannet M. Dempsey D.A. Imani J. Coustau C. Kogel K.H. RNA-based technologies for insect control in plant production Biotechnol. Adv.20203910746310.1016/j.biotechadv.2019.10746331678220 · doi ↗ · pubmed ↗
- 6Zhu K.Y. Palli S.R. Mechanisms, applications, and challenges of insect RNA interference Annu. Rev. Entomol.20206529331110.1146/annurev-ento-011019-02522431610134 PMC 9939233 · doi ↗ · pubmed ↗
- 7Shen C.H. Jin L. Fu K.Y. Guo W.C. Li G.Q. RNA interference targeting Ras GT Pase gene Ran causes larval and adult lethality in Leptinotarsa decemlineata Pest Manag. Sci.2022783849385810.1002/ps.682235104039 · doi ↗ · pubmed ↗
- 8Zhang J. Khan S.A. Hasse C. Ruf S. Heckel D.G. Bock R. Full crop protection from an insect pest by expression of long double-stranded RN As in plastids Science 201534799199410.1126/science.126168025722411 · doi ↗ · pubmed ↗