A Biofuel Cell for Electricity Generation from Biomass-Derived Cellobiose
Piyanut Pinyou, Peeranat Jatooratthawichot, Luciranon Sribrahma, Salila Pengthaisong, Chamaipon Beagbandee, Kantapat Chansaenpak, Vincent Blay, James R. Ketudat Cairns

TL;DR
Researchers created a biofuel cell that generates electricity from cellobiose using enzymes and found that certain biomass pretreatments improve performance.
Contribution
A novel bioanode design using β-glucosidase and glucose oxidase for efficient electricity generation from cellobiose.
Findings
Alkaline pretreatment of biomass improves biofuel cell performance compared to phosphoric acid.
Adding β-glucosidase enhances current generation even with commercial cellulase.
The bioanode efficiently converts cellobiose to electricity via a two-enzyme cascade.
Abstract
We have developed a new bioanode based on a cascade of reactions catalyzed by two enzymes. A glassy carbon electrode is modified with β-glucosidase and glucose oxidase enzymes entrapped within an osmium redox polymer. Cellobiose, the fuel for the anode, is hydrolyzed by β-glucosidase (TxGH116), yielding two molecules of D-glucose. Glucose is then oxidized by glucose oxidase (GOx) into δ-gluconolactone and produces electrons that are transferred to the electrode mediated by osmium redox polymer. We investigated the kinetic parameters of both enzymes at different temperatures. For GOx, the effect of enzyme loading and enzyme/polymer ratio were also optimized. The proposed bioanode is coupled to a biocathode based on horseradish peroxidase (HRP) in which H2O2, the oxidant, is reduced. We investigated the performance of the biofuel cell on cellobiose and sugarcane hydrolysates subjected to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
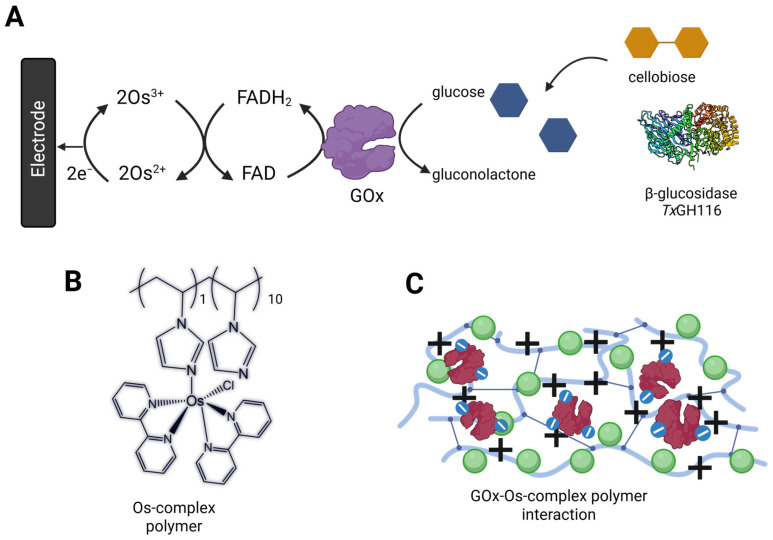 Figure 1
Figure 1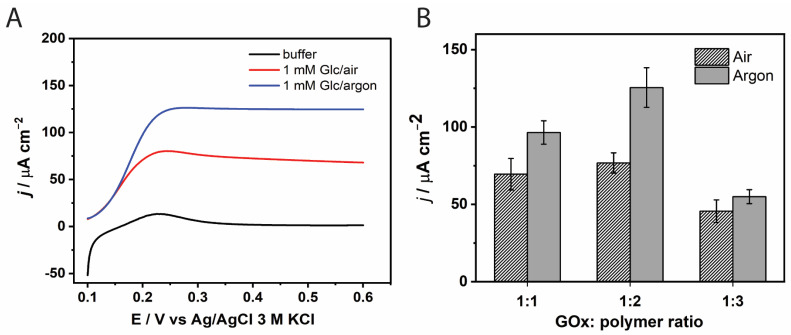 Figure 2
Figure 2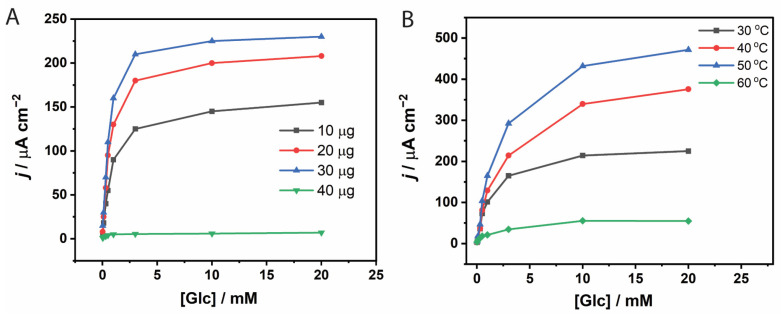 Figure 3
Figure 3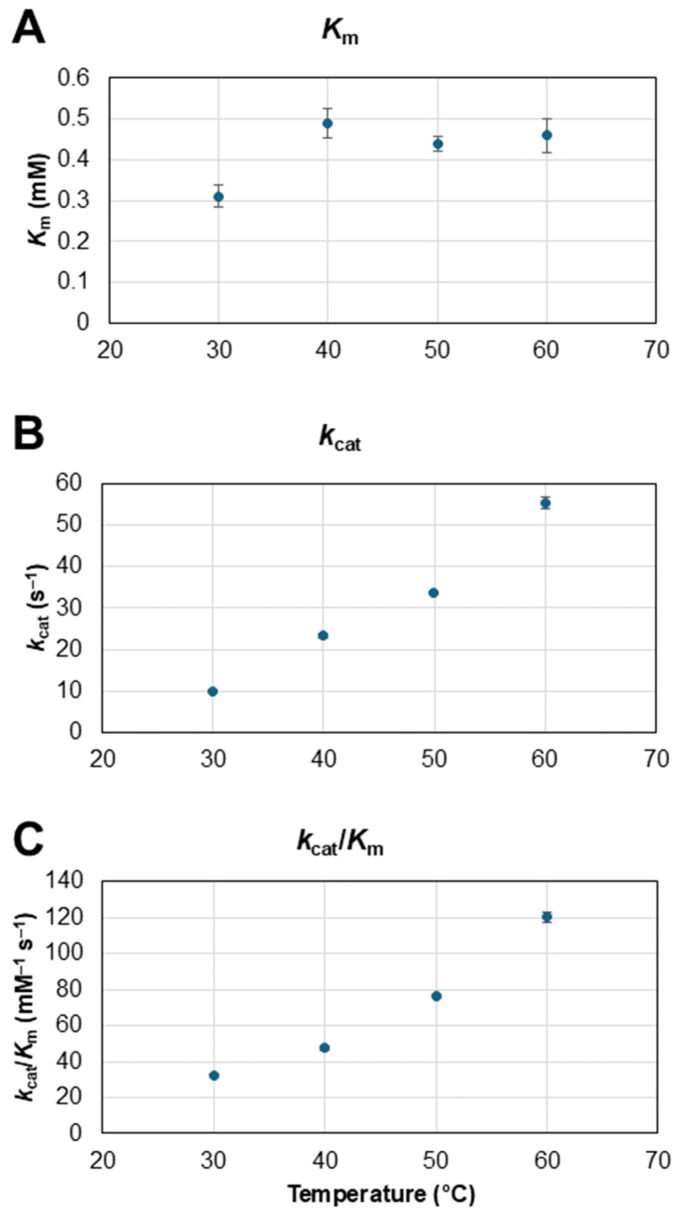 Figure 4
Figure 4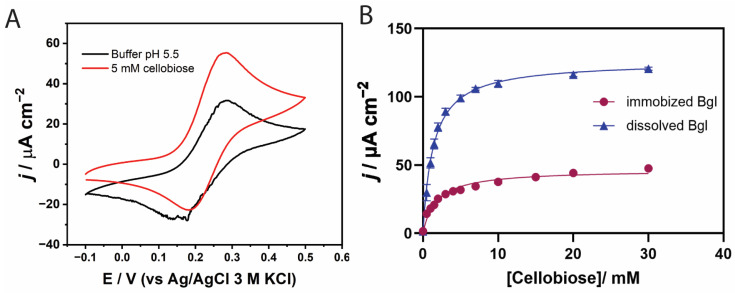 Figure 5
Figure 5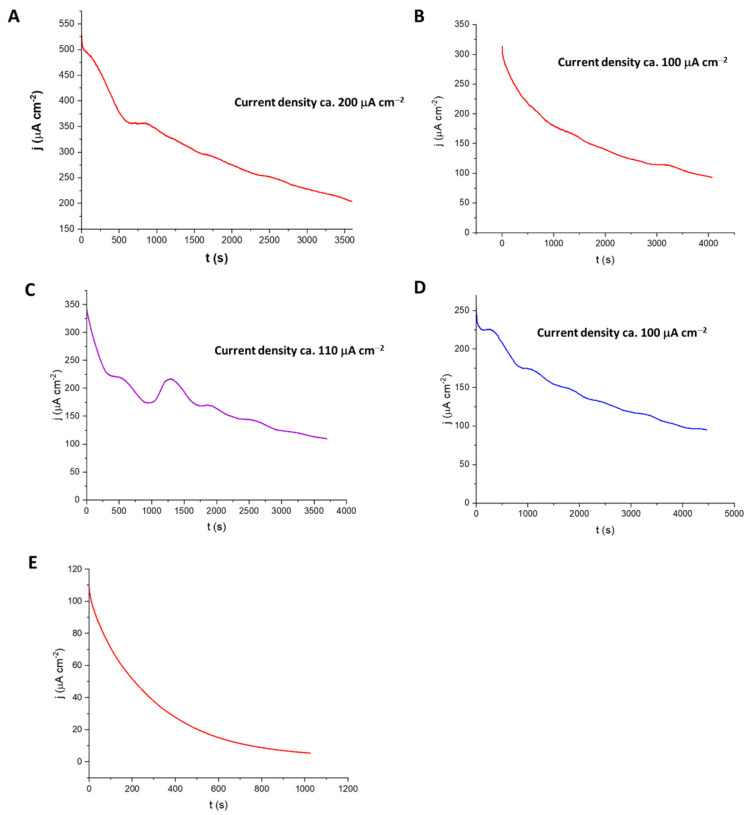 Figure 6
Figure 6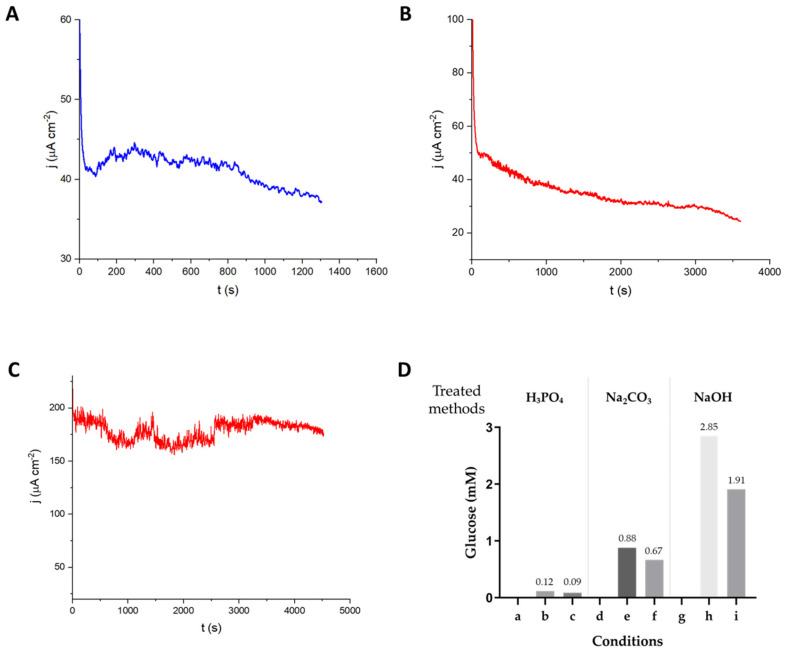 Figure 7
Figure 7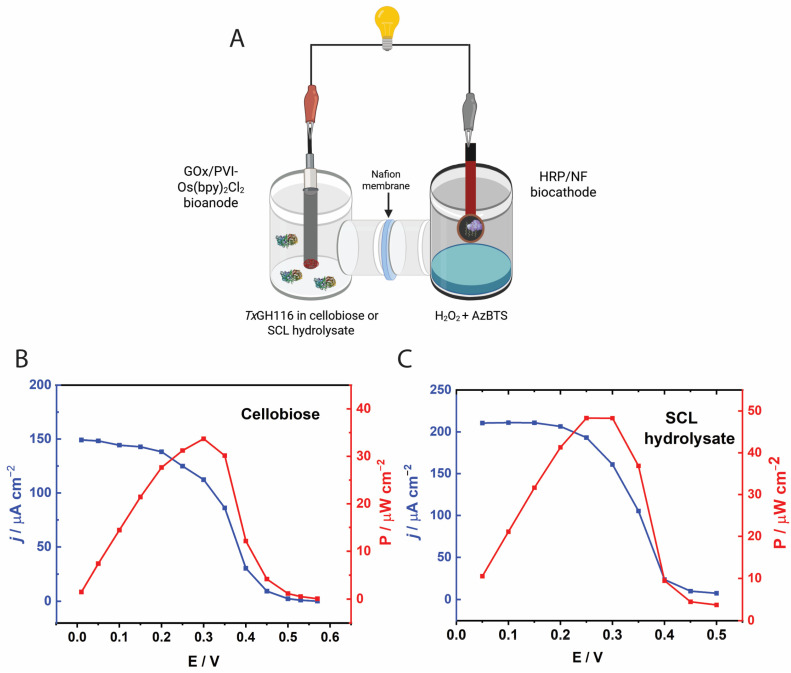 Figure 8
Figure 8- —Suranaree University of Technology (SUT)
- —Thailand Science Research and Innovation
- —National Science, Research and Innovation Fund (NSRF)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Fuel Cells and Bioremediation · Biofuel production and bioconversion · Microbial Metabolic Engineering and Bioproduction
1. Introduction
The pursuit of sustainable and efficient energy solutions has intensified the interest in biofuel cells (BFCs), particularly enzymatic biofuel cells (EBFCs), which convert biochemical energy into electrical energy under mild, environmentally friendly conditions [1]. Multi-enzyme cascades have emerged as a powerful approach to enhance EBFC performance [2]. In these systems, multiple enzymes enable a reaction network that converts complex substrates into valuable products or energy carriers [3,4,5]. In EBFC systems, by enabling a more complete oxidation of fuels through sequential catalytic steps, multi-enzyme cascades offer significantly higher energy conversion efficiency and greater power density than single-enzyme systems [6,7]. For instance, Minteer’s group demonstrated the oxidation of glucose to carbon dioxide through a cascade of six enzymes, achieving the extraction of 24 electrons per glucose molecule. Large biomass-derived molecules, such as cellulose, glycogen, and starch, can also serve as fuels after enzymatic breakdown [8,9]. For example, hydrolases such as cellulases break down lignocellulosic materials into fermentable sugars [10], and oxidoreductases such as glucose oxidase (GOx) [10] or pyranose dehydrogenase (PDH) [11] can oxidize these sugars, releasing electrons that can be captured as electrical current. Recently, Wu et al. developed a biosensor for the real-time monitoring of sucrose uptake in plants. The EBFC utilized a carbon fiber anode modified with a three-enzyme cascade consisting of an invertase (hydrolase), a mutarotase (isomerase), and a glucose oxidase (oxidoreductase) [12]. Sequential multi-enzyme conversions over electrodes provide a direct path to transform abundant biomass into electrical energy [13], supporting the 7th Sustainable Development Goal—affordable and clean energy
β-Glucosidase (EC 3.2.1.21) plays a pivotal role in cellulose-based biomass degradation [13]. It catalyzes the hydrolysis of glucosides such as cellobiose into glucose [14,15]. Nearly all β-glucosidases hydrolyze glucosides by a double-displacement mechanism that leads to retention of chirality at the anomeric carbon [16,17]. Reducing the levels of cellobiose is also desirable as it can inhibit other enzymes used in biomass saccharification [18]. Enzyme cocktails containing β-glucosidase are used in bioethanol production [19], biorefineries [20], and emerging bioelectrochemical systems [11]. β-Glucosidases also have applications in food and beverage industries [16,21].
Although β-glucosidases are particularly abundant in plants, enzymes of microbial origin are often favored for their rapid degradation of cellulosic materials [22,23,24]. Researchers have also devoted significant efforts to engineering β-glucosidases with improved catalytic efficiency, thermostability, and glucose tolerance [25,26,27], and a wide variety of β-glucosidases are available. A GH116 β-glucosidase from the thermophile Thermoanaerobacterium xylanolyticum, TxGH116, has a temperature optimum of 75 °C and a melting temperature of 81 °C [28]. TxGH116 has a known structure, and the roles of catalytic and glucose-binding amino acids have been delineated by mutagenesis [28,29,30]. Catalytic residue mutants of TxGH116 have proven useful for production of glucosylazides [31,32]. Although TxGH116 is relatively sensitive to glucose inhibition with a competitive Ki of 4 mM, its stability and high catalytic rate on oligosaccharides make it attractive for applications, especially at low to moderate glucose concentrations. In addition, it shows much less inhibition by δ-gluconolactone, the product of GOx, than most β-glucosidases, making it appropriate for application in glucose fuel cells [25].
As a primary product of cellulose hydrolysis, cellobiose is a promising renewable fuel for biofuel cells (BFCs). In this work, we designed and evaluated a bioanode platform capable of efficiently converting cellobiose and cellulosic hydrolysates into electrical current through a two-step enzymatic process. In this system, β-glucosidase TxGH116 catalyzes the hydrolysis of cellobiose into D-glucose, which is subsequently oxidized by GOx to δ-gluconolactone, with electrons transferred to the electrode via mediated electron transfer (MET) using an Osmium redox polymer. This configuration enables current generation from a renewable disaccharide as fuel. As proof of concept, the BFC system was evaluated using both cellobiose and hydrolysates derived from sugarcane leaves.
2. Materials and Methods
2.1. Reagents, Enzymes and Solutions
D-Glucose, glucose oxidase (GOx) from Aspergillus niger (≥65 U mg^−1^), and D-(+)-cellobiose were obtained from TCI Chemicals (Japan). Poly(ethylene glycol) diglycidyl ether (PEGDGE), horseradish peroxidase (HRP (≥174 U mg^−1^), 2,2′-azino-bis-(3-ethylbenzothiazoline-6-sulfonic acid) ammonium salt (AzBTS) and Nafion^TM^ 117 were purchased from Sigma-Aldrich (St. Louis, MO, USA). PVI-Os(bpy)2_Cl_2 was prepared according to the established protocol [33]. A bioanode electrode was modified on a surface of a 4 mm glassy carbon disk electrode (Rotating disk electrode, RDE, ALS, Japan). TxGH116 was expressed and purified as previously described with a single immobilized metal affinity chromatography step [28].
2.2. ThCel6A Production and Purification
Based on the bacteria’s tolerance to high temperatures and alkaline pH, a Thermobifida halotolerans glycoside hydrolase family GH6 endoglucanase (NCBI Accession AHN09982, ThCel6A) was selected. A gene optimized for expression in Escherichia coli encoding this protein without its N-terminal signal sequence (residues 31-443) and containing a stop codon (Figure S1) was synthesized and cloned into the NcoI and XhoI sites of pET32a by Gene Universal Corporation. The plasmid was used to transform E. coli strain BL21(DE3) and the protein produced in an 800 mL LB broth culture in a 2 L shake flask induced with 0.2 M IPTG for 18 h at 20 °C shaking at 200 rpm. The cells were centrifuged at 4000× g 20 min at 4 °C to remove the media and the cell pellet frozen at −80 °C, then thawed and lysed in 20 mL lysis buffer containing 50 mM sodium phosphate, pH 7.5, 0.2 mg/mL lysozyme, 1% Triton-X 100, 1 mM phenylmethylsulfonyl fluoride (PMSF), 1 mM 6-aminohexanoic acid, 1 mM benzamidine hydrochloride and 5 µg/mL DNase I for 30 min at 25 °C. The cell debris was removed by centrifugation at 12,000× g for 20 min at 4 °C. The protein was purified over an immobilized metal affinity chromatography resin column (GE Healthcare) bound with CoCl_2_ and equilibrated in 150 mM NaCl, 50 mM sodium phosphate, pH 7.5 (eq buffer). After loading the column, it was washed with 5 column volumes each eq buffer, 5 mM imidazole in eq buffer, and 10 mM imidazole in eq buffer, then eluted with 250 mM imidazole in eq buffer. The fractions containing ThCel6A were pooled and concentrated and the buffer exchanged with 150 mM NaCl, 20 mM Tris, pH 8.0 (enzyme storage buffer) in a centrifugal filter, MWCO 30,000 (Merck Millipore, Darmstadt, Germany). The protein concentration was determined by measuring absorbance at 280 nm on a Nano drop spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA, NanoDrop™ 2000/2000c). Sodium dodecyl sulfate-polyacryamide gel electrophoresis (SDS-PAGE) was performed on 12% acrylamide gels to determine the purity and molecular mass of the protein which was visualized by Coomassie Blue R-250 staining.
2.3. Enzymatic Characterization of ThCel6A
The recombinant ThCel6A was tested for hydrolysis of the soluble β-glucan substrates carboxymethyl cellulose (CMC) and barley β-glucan by the 3,5-dinitrosalicylic acid (DNS) assay for reducing sugars [34]. The enzyme (2 µg) was assayed on 0.5% CMC or barley β-glucan in 50 mM sodium acetate, pH 5.5, at temperatures ranging from 20 to 80 °C for 24 h, followed by boiling 5 min to stop the reactions and DNS assay of released sugars. For the pH optimum, the enzyme was assayed in McIlvaine citrate/phosphate universal buffers over a range of pH 3.5 to pH 9.0 in 0.5 unit intervals at 50 °C for 30 min. To evaluate the temperature stability, the enzyme was incubated at 40, 50, 55 or 60 °C for 1 to 24 h in 50 mM sodium acetate, pH 5.5, then assayed for hydrolysis of 0.5% CMC at 50 °C for 30 min; then the reaction stopped by boiling and the reducing sugar released assayed by the DNS assay. The products of hydrolysis of CMC and barley β-glucan were also assessed by stopping the reactions at various time points by boiling, then evaluating the supernatant by thin-layer chromatography (TLC) on silica gel 60 plates (Merck KGaA, Darmstadt, Germany). The plates were developed in butanol:water:acetic acid (4:2:2, v/v/v). The products were visualized by spreading 10% sulfuric acid in ethanol on the plate and heating to develop gray spots at carbohydrates.
2.4. Enzymatic Hydrolysis of Insoluble Biomass
To prepare biomass, sugarcane leaf was dried in an oven at 65 °C until it was completely dried. Then, it was cut into approximately 1 cm long pieces and was milled in a PULVERISETTE 16 cross beater mill (Fritsch, Germany) and passed through a 500 μm sieve. For alkaline pretreatments, 2 g of milled leaf was pretreated with 50 mL of 2% (w/v) sodium carbonate or sodium hydroxide that was autoclaved at 121 °C 20 min. The pretreated sugarcane leaf was rinsed with distilled water until it reached a pH of about 7 (3–5 times), then dried in the oven at 65 °C overnight. Phosphoric acid pretreatment was performed as described by Wood et al. [35] and Zhang et al. [36].
The pretreated sugarcane leaf biomass was digested in either commercial cellulase (Novozyme Cellic Celltec2) or ThCel6A. 10 mL of a 1% (w/v) suspension of pretreated sugarcane leaf powder in 50 mM sodium acetate buffer pH 5.5 was digested overnight with 1% (w/v) cellulase or 3 mg ThCel6A to produce the hydrolysates for testing in the bioanode.
2.5. Bioanode and Biocathode Preparation
A bioanode was prepared by drop casting 7 μL of a mixture containing 40 μg PVI-Os(bpy)2_Cl_2, 20 μg GOx and 21 μg PEGDGE in DI water on the electrode surface of a 4 mm glassy carbon electrode. The modified electrodes were left to dry in the air at room temperature and subsequently kept in the refrigerator at 4 °C overnight. Prior to measurement, the modified electrodes were rinsed with citrate phosphate buffer pH 5.5 to remove the weakly adsorbed components from the electrode surface. A biocathode was prepared by drop-casting 20 µL of 1-pyrenebutyric acid N-hydroxysuccinimide ester (PBSE) at a concentration of 4 mg/mL in dimethylformamide (DMF) onto the surface of a 9 mm graphene-coated polyimide electrode. The electrode was incubated at room temperature for 30 min to allow π–π stacking interactions between PBSE and the graphene surface. Following incubation, the electrode was gently rinsed with DMF to remove any unbound PBSE and then dried under ambient conditions. Subsequently, 100 µg of 10 mg/mL (10 μL) horseradish peroxidase (HRP) in phosphate buffer was drop-cast onto the PBSE-modified electrode surface and allowed to dry at room temperature. After drying, 2 µL of 5% (w/v) Nafion solution was drop-cast over the enzyme layer to immobilize and protect the biomolecule. The fully assembled biocathode was then stored at 4 °C overnight to ensure proper stabilization of the enzyme and Nafion matrix.
2.6. Electrochemical Characterization
The electrochemical characterization and optimization of both the bioanode and biocathode were performed with a PalmSens potentiostat (PalmSens, Utrecht, The Netherlands). A conventional three-electrode setup was employed, comprising a platinum sheet (1 × 1 cm) as the counter electrode and an Ag/AgCl 3 M KCl reference electrode (Italsens, Italy).
For bioanode experiments, citrate buffer (pH 5.5) was used as the supporting electrolyte. The catalytic performance of the GOx-modified bioanode was assessed by chronoamperometry, monitoring the oxidation current of glucose at a constant applied potential of 0.28 V versus Ag/AgCl 3 M KCl. Prior to measurements, the electrolyte was purged with argon (Air Liquide, Thailand) for at least 30 min to remove dissolved oxygen, and all electrochemical measurements were subsequently carried out under an argon atmosphere.
Temperature control during measurements was achieved with a water-jacketed electrochemical cell equipped with a temperature controller (IKA-Werke GmbH & CO. KG, Staufen im Breisgau, Germany, model ICC basic), ensuring the electrolyte was equilibrated to the desired experimental temperature. For biocathode studies, measurements were conducted under ambient air.
2.7. Biofuel Cell Performance
The performance of the cellobiose/H_2_O_2_ enzymatic biofuel cell (BFC) was evaluated in a two-compartment electrochemical cell separated by a Nafion™ 117 membrane. The glucose oxidase (GOx) bioanode was connected to the working electrode and immersed in 20 mL of 50 mM sodium acetate buffer pH 5.5 containing 30 mM cellobiose or 1% (w/v) sugarcane leaf hydrolysate supplemented with 93 μg of TxGH116.
The horseradish peroxidase (HRP) biocathode was connected to the combined counter and reference electrodes and immersed in 25 mL of electrolyte solution containing 10 mM H_2_O_2_ and 1.0 mM AzBTS. Prior to electrochemical measurements, the anolyte was left to incubate for 15 min after the addition of TxGH116.
Open-circuit voltage (OCV) was recorded until a stable voltage output from the BFC was obtained. Subsequently, multistep amperometry was performed by applying a series of potential steps ranging from 0.60 V to 0.010 V vs. the OCV of the BFC. During operation, the bioanode compartment was maintained under an argon-saturated atmosphere, while the biocathode was exposed to an air-saturated environment. The electrolyte temperature was kept constant at 40 °C throughout the experiment.
3. Results and Discussion
3.1. GOx-Modified Anode Characterization and Optimization
The bioanode was designed to utilize cellobiose as its substrate. This disaccharide is hydrolyzed by β-glucosidase into two molecules of D-glucose, as illustrated in Figure 1. Since β-glucosidase is a hydrolase and does not participate in redox reactions, the electrochemical activity of the bioanode relies on the GOx-catalyzed oxidation of glucose. Therefore, the bioanode was initially optimized using glucose as the substrate. The electrode was modified by entrapping GOx within a poly(1-vinylimidazole)-osmium complex [PVI–Os(bpy)2_Cl_2] matrix.
Redox polymers bearing osmium (Os) complexes have been widely employed for the co-immobilization with GOx, enabling electron transfer from the enzymatic oxidation of glucose to the electrode surface via a mediated electron transfer (MET) pathway through an electron-hopping mechanism [37,38,39]. In addition to facilitating efficient electron shuttling, redox polymers support the entrapment of large quantities of redox enzymes, forming multilayered enzyme assemblies. The resulting redox hydrogel provides a three-dimensional network that promotes rapid substrate diffusion and enhances charge transport across the film [40].
3.1.1. Optimization of GOx to Redox Polymer Mass Ratio
Optimizing the relative amounts of redox polymer and GOx is crucial to ensure efficient electron transfer within the bioanode. A sufficient number of redox relay units, provided by the osmium complex in the polymer is essential to electrically wire the redox-active site of GOx to the electrode surface, thereby enabling effective mediated electron transfer.
The influence of the GOx-to-redox-polymer ratio on the current density was investigated under both oxygenated and deoxygenated (argon-saturated) conditions, as shown in Figure 2B. Previous studies have reported negative effects of oxygen on the current output of GOx-modified electrodes [41,42]. Two mechanisms have been proposed to explain this effect: (i) oxygen can react with the reduced form of GOx, producing hydrogen peroxide (H_2_O_2_), which may degrade the electrode components, and (ii) oxygen competes with the redox mediator in re-oxidizing GOx, thereby hindering efficient electron transfer [41]. Consistent with these reports, our results show that current densities measured in air-equilibrated electrolyte were significantly lower than those obtained in argon-saturated conditions (Figure 2).
Under argon-saturated conditions, the highest current density was observed at a GOx:PVI-Os(bpy)2_Cl_2 mass ratio of 1:2 (Figure 2B). This enhancement can be attributed to the optimal availability of osmium-based redox relays, which facilitates efficient electron transport via the electron-hopping mechanism. However, further increasing the amount of redox polymer (1:3) led to a marked decrease in current density. This reduction may be due to structural changes in the redox hydrogel in the acidic hydrolysate electrolyte. At low pH, the poly(1-vinylimidazole) (PVI) backbone undergoes substantial swelling due to protonation of the imidazole groups [43]. Mao et al. reported that a hydrogel based on a PVI-Os complex can swell to 3.5 times its dry thickness [44,45]. As a result, even a slight increase in polymer loading may increase film thickness and hinder glucose diffusion to the enzyme’s active site.
Based on these findings, a GOx-to-polymer mass ratio of 1:2 was selected, and all subsequent measurements of the GOx-modified electrode were performed in an argon-saturated electrolyte.
3.1.2. Effect of GOx Loading Amount
The GOx loading on the electrode surface was optimized by varying the enzyme amount from 10 to 40 µg (Figure 3A). An increase in current density was observed with increasing GOx from 10 to 30 µg. However, the enhancement in catalytic current between 20 and 30 µg was marginal. This plateau is attributed to the limited number of osmium redox centers in the polymer matrix, which restricts the ability to effectively shuttle electrons from the reduced form of GOx, thereby limiting the current output. Moreover, GOx is a non-conductive protein, and excessive loading may hinder glucose diffusion and further reduce catalytic efficiency. Considering both the current response and enzyme utilization, 20 µg of GOx was selected as the optimal loading for further studies.
3.1.3. Effect of Temperature on Immobilized GOx Kinetics
Amperometric measurements with the GOx-modified electrode were conducted at electrolyte temperatures ranging from 30 °C to 60 °C. Increasing the temperature from 30 °C to 50 °C resulted in a steady rise in current density, reflecting faster enzymatic and electrochemical reaction rates (Figure 3B). However, a sharp decline in current density was observed when the temperature approached 60 °C, likely due to thermal instability or partial detachment of the GOx–polymer film from the electrode surface. Based on these findings, a temperature of 40 °C was selected for subsequent experiments to ensure sufficient catalytic activity while maintaining the structural integrity of the system.
3.2. Effect of Temperature on TxGH116 β-Glucosidase
Although TxGH116 is stable at 60 °C [35], the effect of temperature on its cellobiose hydrolysis efficiency had not been previously assessed. Thus, the kinetics of cellobiose hydrolysis by TxGH116 were assessed at temperatures over the range of 30 to 60 °C. As seen in Figure 4, the catalytic rate (kcat) and specificity constant (kcat/Km) for hydrolysis of cellobiose increased ca. 1.5–2-fold every 10-degree increase in this range, while Km was relatively stable. The relevant parameter at the high cellobiose concentration in the electrode is kcat, which was 24 s^−1^ at the 40 °C temperature used for the electrode. If GOx electrode stability can be improved in the future, we could increase glucose production and subsequent power output by increasing the temperature.
3.3. Evaluation of Bi-Enzymatic GOx/β-Glucosidase Catalysis
While glucose in solution is an equilibrium mixture of 64% β-anomer and 36% α-anomer [46], the overall enzymatic hydrolysis of cellobiose by a retaining β-glucosidase is estimated to yield approximately 82% β-glucose, which will gradually equilibrate by mutarotation to the equilibrium composition. Because GOx selectively oxidizes β-D-glucose, we expect this out-of-equilibrium composition to be advantageous, as long as cellobiose does not inhibit GOx.
Before incorporating cellobiose as a substrate in the bi-enzymatic electrode system, we evaluated the potential inhibitory effect of cellobiose on the catalytic activity of GOx toward glucose oxidation. Amperometric measurements were conducted with the GOx-modified electrode in the presence of a constant 1 mM glucose concentration and cellobiose concentrations from 1.0 to 10 mM. As shown in Figure S2, the catalytic current remained essentially unchanged across the tested range of cellobiose concentrations, indicating that cellobiose does not interfere with or inhibit GOx activity under the given conditions.
We investigated the electrochemical behavior of the enzyme-modified electrode using cyclic voltammetry. The cyclic voltammogram of the co-immobilized GOx/β-glucosidase in the redox hydrogel-modified bioanode is shown in Figure 5A. In the absence of cellobiose, the voltammogram exhibited quasi-reversible redox peaks corresponding to the Os^3+^/Os^2+^ redox couple, with a half-wave potential (E_1_/2) of 0.23 V, consistent with previous reports using the same polymer matrix [47]. Following the addition of cellobiose, the electrolyte was incubated with the modified electrode for 15 min to allow for hydrolysis by β-glucosidase. The resulting voltammogram displayed an increased anodic current associated with glucose oxidation at the electrode surface, accompanied by a cathodic peak shift of approximately 25 mV compared to the cellobiose-free condition. This observation indicates successful electron mediation between the FAD redox centers in GOx and the Os-complex in the polymer matrix.
The catalytic performance of the bi-enzymatic system under two different enzyme configurations. In the first setup, β-glucosidase was added directly to the electrolyte to catalyze the homogeneous hydrolysis of cellobiose, generating glucose in solution prior to its oxidation by the immobilized GOx. In the second configuration, both GOx and β-glucosidase were co-immobilized on the electrode. As shown in Figure 5B, the current density with β-glucosidase in solution (blue) was approximately three times higher than that in the co-immobilized system (purple) with a jmax of 120.5 mA cm^−2^. This enhancement can be attributed to improved substrate accessibility—cellobiose can interact with the active site of free β-glucosidase from all directions, whereas mass transport limitations may hinder access to the immobilized enzyme. In agreement with this, the freely diffusing β-glucosidase system exhibited a lower apparent Michaelis–Menten constant (Km^app^ = 1.37 mM) than the system with β-glucosidase immobilized (Km^app^ = 1.80 mM), indicating saturation at a lower cellobiose concentration. Based on these findings, the homogeneous β-glucosidase system was selected for subsequent experiments.
3.4. Current from Sugarcane Leaf Treated with Cellulase
To assess whether the fuel cell could be used to generate electricity from natural biomass rather than from the model substrate cellobiose, biomass was digested with commercial cellulase and the supernatant added to the biocathode with and without TxGH116 β-glucosidase. Sugarcane leaves were selected as the agricultural waste to be evaluated, and three pretreatment methods were compared.
As seen in Figure 6, treatment with a 2 M sodium carbonate alkali led to the best output for this system. Adding TxGH116 β-glucosidase provided an additional 100–200 μA cm^−2^ of output at each point over a 3500 s time course. A similar but less pronounced effect was seen with sodium hydroxide pretreatment of the sugarcane leaf biomass, although with lower current, while phosphoric acid-treated sugarcane leaf gave much lower current. The current density of ca 200 μA cm^−2^ for the system with commercial cellulase anve d added TxGH116 suggests that this system could generate useful current from sugarcane leaf biomass.
3.5. Current from Sugarcane Leaf Treated with ThCel6A and TxGH116
The presence of a complex mixture of enzymes including β-glucosidase in the commercial cellulase mixture obscures the role of the β-glucosidase added to our bioanode. To better dissect the role of β-glucosidase, we generated a system with a single endoglucanase, ThCel6A, which was previously shown to be thermostable and release cellobiose from CMC [48].
We expressed ThCel6A with an additional thioredoxin fusion tag at the N-terminus and without the signal sequence at high levels and the purified (Figure S3) had high activity over a broad range of pH with a peak around pH 8–8.5 (Figure S4A), in line with the reported pH optimum [48], and 30% maximum activity at pH 5.5. The temperature optimum was 45 °C and it maintained nearly 80% maximal activity up to 65 °C (Figure S4B). The enzyme was stable up to 55 °C, with over 75% activity after 24 h (Figure S5). The good stability over the range of temperatures appropriate to our system and the broad pH range are favorable for use with the TxGH116 β-glucosidase, whose optimal pH is 5.5.
The main ThCel6A hydrolysis products after 24 h digestion of barley β-glucan, CMC, phosphoric acid swollen cellulose (PASC), base-treated cellulose, and base-pretreated rice straw and sugarcane leaves were di-, tri- and tetra-saccharides (Figures S6 and S7), which are appropriate for TxGH116 β-glucosidase digestion. Glucose production was not seen except for a small amount from PASC, so use of ThCel6A could make the contribution of TxGH116 to the bioanode clearer than hydrolysis with commercial cellulase.
Sugarcane leaf powders with three different pretreatments were tested in the bioanode after hydrolysis with ThCel6A. The TxGH116 β-glucosidase was also added to the digest to have glucose available at the start of the current measurement. As seen in Figure 7, pretreatment with phosphoric acid or sodium carbonate gave initial current densities around 40 µA cm^−2^ and maintained the current density above 30 µA cm^−2^ for more than 20 min. Notably, sodium hydroxide pretreatment provided a higher current density with around 180 µA cm^−2^ for 80 min (Figure 7C). Thus, the sodium hydroxide pretreatment gave about 4.5 to 6-fold higher current density than the other two. Over the time measured, the glucose concentration in the anolyte decreased by about one third (Figure 7D), since glucose was mainly released before the digestate was added to the bioanode. The current density was lower than that with hydrolysis with commercial cellulase, likely because the commercial cellulase includes many enzymes that can work together to more effectively break down complex biomass samples to release oligosaccharides for hydrolysis by β-glucosidase. However, the experiment with ThCel6A demonstrated the effectiveness of TxGH116 in converting oligosaccharides released from biomass to glucose for generation of electrons by glucose oxidase, since it was the only β-glucosidase in the mixture.
3.6. BFC Performance with Cellobiose and SCL Hydrolysates
The performance of the biofuel cell (BFC) using cellobiose as the fuel was initially evaluated, as shown in Figure 8A. The polarization curve, obtained in a buffer containing 30 mM cellobiose after a 15 min incubation with TxGH116 prior to electrochemical measurements, displayed a typical decrease in current density with increasing cell voltage, particularly near the open-circuit voltage (OCV). The observed OCV was approximately 0.57 V. The BFC achieved a maximum current density of around 150 µA cm^−2^. The corresponding power density curve peaked at approximately 0.30 V, yielding a maximum power output of about 33.7 µW cm^−2^. Although the need for a 15 min preincubation with β-glucosidase to build a reservoir of glucose may hinder continuous use, it may be less critical for use with commercial cellulase-treated biomass which already contains released glucose. Future improvement in β-glucosidase for more rapid hydrolysis can be used to achieve an appropriate steady-state level of glucose. Nonetheless, this experiment suggests that useful output from biomass is feasible.
Figure 8B presents the BFC performance when the bioanode was operated in 1% SCL hydrolysates derived from either commercial cellulase pretreated with Na_2_CO_3_. The OCV values obtained from both pretreated hydrolysates were approximately 0.50 V, which were lower than the OCV observed during operation with pure cellobiose (0.57 V). This reduction in OCV can be attributed to the increased complexity of the hydrolysate matrices, which may contain insoluble residues and non-fermentable components [49]. These residual substances likely contributed to elevated solution resistance and may have interfered with electron transfer at the bioanode. Additionally, certain compounds (phenolic derivatives) present in the hydrolysates could have exerted inhibitory effects on the enzymatic activity of β-glucosidase and GOx at the bioanode [50,51], thereby lowering the overall electrochemical potential of the BFC system. The BFC utilizing hydrolysate from commercial cellulase showed significantly enhanced performance, attaining a maximum current density of 210 µA cm^−2^ and a peak power density of 48 µW cm^−2^ at 0.30 V. This improvement reflects the efficient enzymatic saccharification of lignocellulosic biomass, resulting in higher glucose concentrations readily oxidized by the GOx-based bioanode.
4. Conclusions
In this study, we successfully demonstrated a cascade enzymatic catalysis strategy in the bioanode of a BFC, employing β-glucosidase and GOx for the hydrolysis and oxidation of cellobiose, a key product from lignocellulosic biomass. To enhance electron transfer efficiency between GOx and the electrode, an osmium-complex-modified polymer was utilized as a redox mediator, facilitating electrical communication between the FAD redox centers and the electrode surface.
Due to the relatively slow kinetics of β-glucosidase, a 15 min incubation was allowed before electrochemical measurements to allow for sufficient cleavage of cellobiose before electrooxidation of glucose catalyzed by GOx. Electrochemical analysis revealed that the bioanode configuration with GOx immobilized on the electrode and β-glucosidase dissolved in the electrolyte exhibited significantly higher current density compared to the co-immobilization of both enzymes on the electrode. This optimized bioanode system was successfully applied to generate current from hydrolysates of sugarcane leaf biomass pretreated under various conditions and with a cellulase cocktail or two β-glucosidases.
The application of this bioanode was further demonstrated by coupling it with a HRP-based biocathode for hydrogen peroxide reduction, with AzBTS serving as the redox mediator. When operated with cellulase hydrolysates of sugarcane leaf pretreated with sodium carbonate, the assembled BFC delivered a maximum power output of 50 µW cm^−2^ at an operating voltage of 0.4 V and an open-circuit voltage (OCV) of 0.5 V. Overall, this work highlights the potential of combining β-glucosidase- and GOx-catalyzed reactions for energy generation from biomass. Furthermore, the development and application of novel β-glucosidases with faster kinetics and high glucose tolerance through enzyme engineering may expand the performance of this system for future bioenergy applications.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Xiao X. Xia H. Wu R. Bai L. Yan L. Magner E. Cosnier S. Lojou E. Zhu Z. Liu A. Tackling the Challenges of Enzymatic (Bio)Fuel Cells Chem. Rev.20191199509955810.1021/acs.chemrev.9b 0011531243999 · doi ↗ · pubmed ↗
- 2Nguyen K.V. Giroud F. Minteer S.D. Improved Bioelectrocatalytic Oxidation of Sucrose in a Biofuel Cell with an Enzyme Cascade Assembled on a DNA Scaffold J. Electrochem. Soc.2014161 H 93010.1149/2.0761414 jes · doi ↗
- 3Tan Z. Cheng H. Chen G. Ju F. Fernández-Lucas J. Zdarta J. Jesionowski T. Bilal M. Designing Multifunctional Biocatalytic Cascade System by Multi-Enzyme Co-Immobilization on Biopolymers and Nanostructured Materials Int. J. Biol. Macromol.202322753555010.1016/j.ijbiomac.2022.12.07436516934 · doi ↗ · pubmed ↗
- 4Megarity C.F. Weald T.R.I. Heath R.S. Turner N.J. Armstrong F.A. A Nanoconfined Four-Enzyme Cascade Simultaneously Driven by Electrical and Chemical Energy, with Built-in Rapid, Confocal Recycling of NADP(H) and ATPACS Catal.2022128811882110.1021/acscatal.2c 0099935966600 PMC 9361290 · doi ↗ · pubmed ↗
- 5Bai J. Li M. Xing F. Wei X. Liu J. Electrically Driven Biocatalysis for Sustainable CO 2-to-Chemicals Transformation Chem Sus Chem 202518 e 20250033410.1002/cssc.20250033440229208 · doi ↗ · pubmed ↗
- 6Macazo F.C. Minteer S.D. Enzyme Cascades in Biofuel Cells Curr. Opin. Electrochem.2017511412010.1016/j.coelec.2017.07.010 · doi ↗
- 7Huang W. Zulkifli M.Y.B. Chai M. Lin R. Wang J. Chen Y. Chen V. Hou J. Recent Advances in Enzymatic Biofuel Cells Enabled by Innovative Materials and Techniques Exploration 202332022014510.1002/EXP.2022014537933234 PMC 10624391 · doi ↗ · pubmed ↗
- 8Shi P. Wu R. Wang J. Ma C. Li Z. Zhu Z. Biomass Sugar-Powered Enzymatic Fuel Cells Based on a Synthetic Enzymatic Pathway Bioelectrochemistry 202214410800810.1016/j.bioelechem.2021.10800834902664 · doi ↗ · pubmed ↗