High Proteolytic and Collagenolytic Activity in an Environmental Vibrio Isolate: Insights into Tissue-Degrading Virulence Factors
Monica Salamone, Aldo Nicosia, Giulio Ghersi, Angela Cuttitta, Paola Quatrini, Marcello Tagliavia

TL;DR
A new Vibrio strain from an octopus shows strong tissue-degrading enzyme activity, suggesting potential virulence and biotech uses.
Contribution
Identification of a novel Vibrio strain with high proteolytic and collagenolytic activity linked to virulence.
Findings
The Vibrio isolate shows high proteolytic and collagenolytic activity.
The strain belongs to the Harveyi clade but is distinct from known strains.
The isolate's enzymes could be useful for biotechnological applications.
Abstract
Vibrio is a genus of ubiquitous aquatic bacteria that includes numerous pathogenic species. Their remarkable genomic plasticity and rapid evolution make them of particular interest from both clinical and ecological perspectives. Successful infection by Vibrio species often relies on multiple virulence factors, including secreted enzymes. Here, we report the characterization of a novel environmental Vibrio strain isolated from a wild octopus that developed fulminant septicaemia accompanied by widespread soft tissue lysis. These severe symptoms prompted a detailed investigation into the bacterium’s identity and enzymatic profile, focused on proteases as potential virulence factors. Multi-locus sequence analysis placed the isolate within the Harveyi clade but revealed no perfect match to known strains, supporting its designation as a novel strain. Biochemical assays demonstrated strong…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
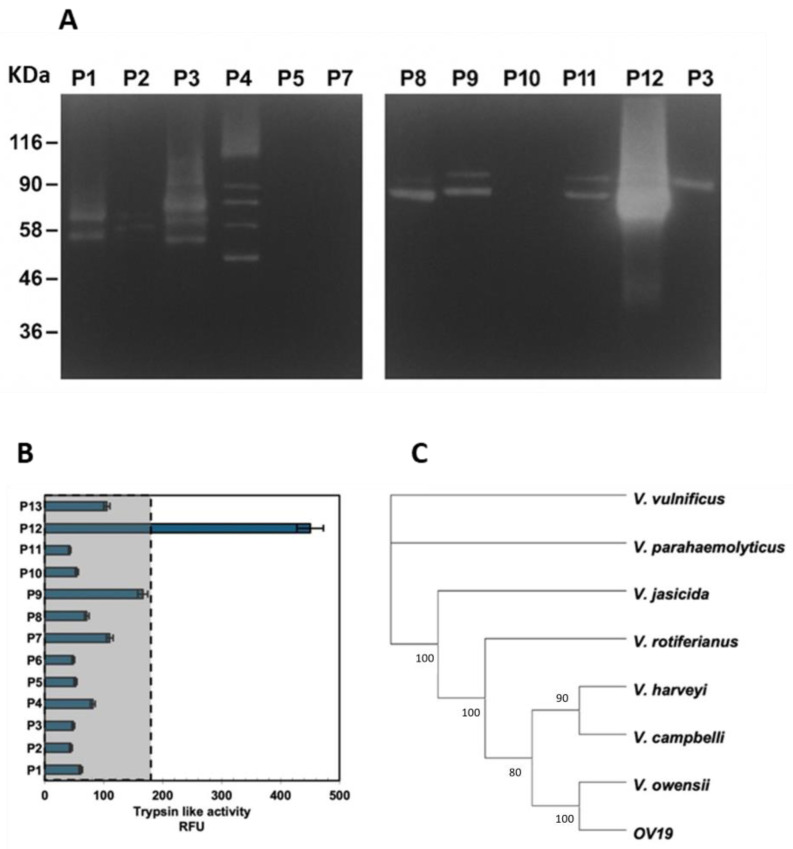 Figure 1
Figure 1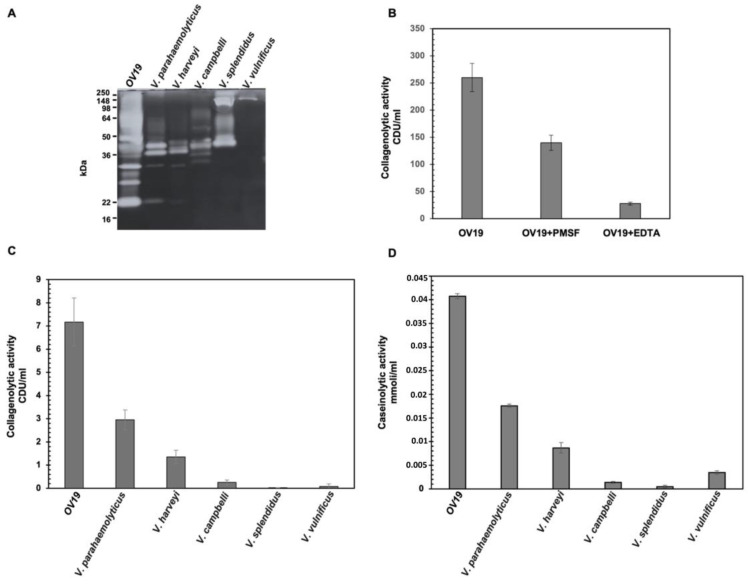 Figure 2
Figure 2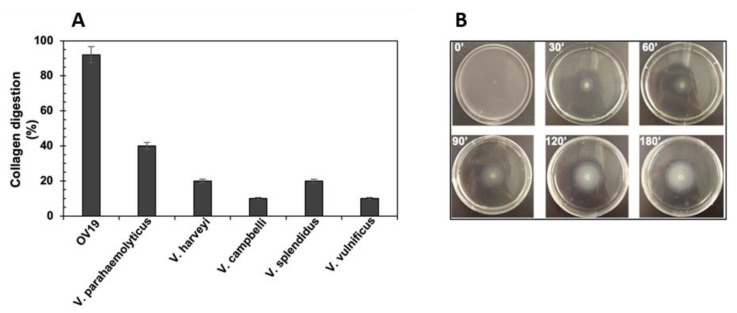 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVibrio bacteria research studies · Aquaculture disease management and microbiota
1. Introduction
Vibrio is a genus of rod-shaped, Gram-negative γ-proteobacteria inhabiting diverse aquatic ecosystems, including marine, estuarine, and brackish waters [1,2]. These bacteria exhibit remarkable ecological versatility and genomic adaptability, supported by a unique genomic architecture comprising two circular chromosomes. Chromosome I typically encodes essential metabolic and housekeeping genes, whereas chromosome II—believed to have evolved from an ancestral megaplasmid—contains genes linked to niche adaptation, environmental sensing, and virulence [3,4].
The genomes of Vibrio species are highly dynamic, undergoing frequent structural rearrangements driven by high rates of horizontal gene transfer (HGT), homologous recombination, and the activity of mobile genetic elements such as transposons, integrons, prophages, plasmids, and genomic islands [5,6,7]. A distinctive feature in several species, most extensively studied in V. cholerae, is the super-integron, a large genetic platform capable of capturing diverse gene cassettes, which facilitates the acquisition of novel functions, thereby accelerating adaptation and evolution [7]. This genomic plasticity allows Vibrio species to rapidly respond to environmental change, colonize novel hosts, and acquire new functional traits, including an expanded enzymatic repertoire [5,6].
Over 100 Vibrio species have been described, with at least 12 recognized as human pathogens [8]. The most studied, V. cholerae, is the etiological agent of cholera, a severe diarrheal disease transmitted via contaminated water. Other clinically relevant species include V. parahaemolyticus, V. alginolyticus, and V. vulnificus, which typically cause vibriosis through ingestion of contaminated seafood or exposure to seawater, particularly in immunocompromised individuals [1,9]. The global incidence of vibriosis is rising, partly due to climate change and associated increases in sea surface temperatures, which are thought to promote proliferation, geographic expansion, and possibly enhanced virulence in Vibrio populations [2,10,11,12,13,14].
Beyond human health, various Vibrio species are significant pathogens in aquaculture and marine wildlife, causing recurrent outbreaks in vertebrate and invertebrate hosts. Species such as V. anguillarum, V. salmonicida, V. harveyi, V. coralliilyticus, and V. crassostreae—most within the Harveyi clade—are associated with diseases in fish, mollusks, and crustaceans, leading to substantial economic losses [11,14].
Vibrio pathogenicity is multifactorial, involving an array of extracellular virulence factors, including hemolysins, enterotoxins, phospholipases, cytotoxins, proteases (such as collagenases and elastases), siderophores, and hemagglutinins; bacterial motility has also been implicated in virulence [15,16,17,18]. Among these factors, proteases play a pivotal role in bacterial invasiveness. Vibrio’s protease repertoire includes zinc metalloproteases (e.g., vibriolysin), serine proteases, cysteine proteases, and neutral elastase-like enzymes, all of which contribute to tissue degradation and invasion, immune evasion, and nutrient acquisition [12]. For example, vibriolysin degrades structural proteins such as collagen, elastin, and fibrinogen, facilitating tissue penetration and inflammatory responses [19]. In fish pathogens like V. anguillarum, proteases are essential for host colonization and mortality [20].
The production of virulence-associated proteases is generally regulated by environmental cues such as iron limitation, temperature, pH, and host-derived signals (e.g., bile salts, tissue contact) [21]. This regulation has been well documented in human pathogens like V. cholerae and V. vulnificus [21,22,23]. Nevertheless, many Vibrio proteases are also expressed under non-host conditions, albeit in a regulated manner, including in natural waters and laboratory culture media [21,22]. Notably, many of these enzymes are halo- and thermos-tolerant, and active across broad pH ranges, supporting roles in general survival strategies such as nutrient utilization and interspecies competition [23,24], in addition to host infection.
Biochemical versatility and diversity of proteases from Vibrio spp. make these bacteria an attractive source of molecules for biotechnological applications spanning medicine (e.g., tissue dissociation for regenerative medicine, wound treatment, etc.), food processing, industrial processes, and environmental remediation [17,25]. Importantly, their high genomic plasticity fosters the emergence of novel strains with unique combinations of enzymatic and pathogenic traits, rendering any isolate a potential source of new bioactive molecules.
Here, we characterize the extracellular proteolytic activities, potentially serving as virulence factors, of a novel Vibrio sp. isolated from a diseased wild octopus. The symptoms made us raise the hypothesis of a major role for proteases in the disease process, prompting molecular identification of the bacterium and detailed enzymatic profiling. The results support classification of the isolate as a strain within the Harveyi clade and reveal distinctive biochemical traits with potential relevance for microbial ecology, pathogenicity, and biotechnological applications, although direct experimental confirmation of its virulence was not undertaken.
2. Results and Discussion
2.1. Bacterial Isolation, Analyses of Enzymes Secreted from Vibrio Isolates and Phylogenetic Analyses
A severely injured wild octopus (Octopus vulgaris), likely attacked by a predator shortly before capture, was found to exhibit symptoms characterized by diffuse, rapidly progressing tissue swelling, disintegration, and necrosis. To investigate whether the pathology was associated with bacterial infection, hemolymph and affected tissues were aseptically collected and cultured on multiple media.
Bacterial loads of approximately 10^5^ cfu/g were detected in both hemolymph and tissues, consistent with systemic infection and peripheral tissue invasion, likely originating from the initial wound.
A preliminary screening aiming to identify putative vibrios was based on growth on thiosulfate–citrate–bile salts–sucrose (TCBS) agar and on PCR amplification using Vibrio-specific primers targeting the rpoD gene [26], which assigned most isolates to the genus Vibrio.
The rapid tissue degradation and the absence of hemolymph clotting, suggestive of pathogenic mechanisms potentially involving potent degradative enzymes, prompted further characterization of the isolates focusing on their proteolytic potential. Sixty-five isolates were randomly selected from Marine Agar plates—including colonies representative of different phenotypes—for proteolytic activity screening. Secreted proteases from liquid cultures were analyzed by gelatin zymography, revealing 13 distinct exoprotease profiles. One profile, corresponding to isolate OV19 (sample P12; Figure 1), displayed particularly strong proteolytic activity compared with the others, and was selected for detailed biochemical characterization.
To compare quantitatively the proteolytic capacity of OV19 with that of representative isolates from each exoprotease profile group (P1–P13), one isolate per group was selected, and the trypsin-like activity in culture supernatants was measured using the specific fluorescent substrate BOC-Gln-Ala-Arg-AMC (Figure 1B).
OV19 exhibited the highest trypsin-like specific activity, with enzyme activity levels nearly fivefold greater than the mean of all other tested isolates. This marked proteolytic capacity reinforced the hypothesis that OV19 could possess higher tissue-destructive potential compared to other isolates, justifying in-depth investigations into its biochemical properties and pathogenic traits.
2.2. Molecular Identification of OV19
Given OV19’s distinctive proteolytic profile, molecular analyses were carried out to enable targeted comparative studies. Since 16S rRNA gene sequencing alone is insufficiently discriminative for Vibrio taxonomy, analyses aiming at identifying the isolate at the species level were carried out firstly based on sequence analysis of pyrH [27], which assigned the isolate to V. owensii. A further analysis, based on rpoA, recA and pyrH sequences considered collectively [28] by BLAST analysis (BLAST+ 2.16.0), showed sequence identity exceeding 99.6% (99.8, 99.6, and 99.7%, respectively) with V. owensii; these percentages were all above the thresholds recommended to assign strains to the same species [28]. To achieve a more robust identification, multilocus sequence analysis (MLSA) was performed on concatenated sequences from seven protein-coding genes (ftsZ, gapA, gyrB, mreB, recA, rpoA, and topA) together with the 16S rRNA gene, using the pipeline described in [29,30].
Each concatenated sequence was first subjected to PSI-BLAST to identify the closest Vibrio matches within a range of 95–100% sequence identity. The resulting dataset was then compared with six Harveyi clade species showing the highest sequence similarity (V. campbellii, V. harveyi, V. rotiferianus, V. jasicida, V. owensii, and V. parahaemolyticus) with V. vulnificus used as an outgroup. Neighbor-joining (NJ) and maximum-parsimony (MP) phylogenetic analyses produced congruent topologies, with V. vulnificus rooting the tree (Figure 1C).
The MLSA placed OV19 within the Harveyi clade, in close association with V. owensii, and at similar pairwise distances to V. campbellii (0.25) and V. harveyi (0.26). Sequence identity analysis revealed values of 98% for ftsZ, 97.4% for gapA, and 100% for rpoA relative to V. harveyi; 99.5% for recA and 97% for mreB relative to V. owensii; and 98% for gyrB and 99% for topA relative to V. campbellii. Therefore, the phylogenetic placement of OV19 appeared intermediate among these closely related type strains, suggesting a mosaic genomic composition. These data partially challenged the earlier identification of V. owensii based on either one or three loci, thereby underscoring the inherent limitations of current molecular approaches for Vibrio identification.
From an evolutionary perspective, these findings highlight the complexity and adaptability of the Vibrio genus, in which frequent horizontal gene transfer facilitates the acquisition of novel genetic material. Such processes may also contribute to shaping the enzymatic arsenal of Vibrio, promoting the emergence of new functional traits, including pathogenic ones [31,32]. Taken together, our data indicate that OV19 might represent a genetically distinct lineage—closely related to V. owensii—within the Harveyi clade. Further information on its genomic structure and functions might be achieved through whole-genome sequencing (WGS) and also from the perspective of biotechnological exploitation of specific genes. Moreover, WGS might help in obtaining insights into the emergence of such a strain, while allowing for a more precise taxonomic resolution. However, it is worth pointing out that taxonomic classification of vibrios is challenging due to their high genomic plasticity and recombination rates. Moreover, the robustness of any bioinformatic analysis strongly depends on the sequence’s availability in GenBank and on criteria employed to assign them at the species level, which is one of the major challenges in Vibrio taxonomy [33].
2.3. Comparison of Collagenolytic Activity of OV19 with Vibrio Type-Strains
To further profile the proteolytic activities of OV19’s secreted enzymes, we focused our analyses on collagenases, since the collagenolytic activity is a common feature of Vibrio strains, particularly of pathogenic and invasive ones [34]. We firstly compared the gelatinolytic activity of OV19 with that of V. vulnificus (well known for its collagenolytic activity) and several Harveyi clade members (V. parahaemolyticus, V. campbellii, V. harveyi, and V. splendidus). Supernatants from overnight cultures grown in appropriate liquid medium were collected, and their gelatinolytic activity was examined by gelatin zymography.
This comparative approach revealed marked differences in the proteolytic profiles of OV19 and the reference type strains (Figure 2A). The tested Vibrio strains produced enzymes spanning an apparent molecular mass range of ~20 to 150 kDa, with band relative intensity varying among species. Notably, the OV19 supernatant displayed a more complex pattern, with numerous bands across different molecular weights, indicative of a diverse and abundant extracellular protease repertoire.
To identify the major classes of proteases contributing to OV19’s activity, zymograms were performed in the presence of either EDTA or PMSF as inhibitors (Figure 2B). PMSF treatment reduced collagenolytic activity by approximately 50%, whereas EDTA inhibited it by more than 90%. These findings suggest that the majority of collagen-digesting proteases in OV19 are cation-dependent metalloproteases, with a significant fraction of activity also attributable to serine and/or cysteine proteases.
To further dissect the contribution of collagenases relative to other proteases, supernatants were tested for their ability to hydrolyze the specific collagenase substrate carbobenzoxy-Gly-Pro-Gly-Gly-Pro-Ala-OH alongside casein to monitor general proteolytic activity. Both assays confirmed that OV19 possesses strong collagenolytic and caseinolytic activities, both outperforming those of reference strains (Figure 2C,D).
Because the assays described above did not strictly distinguish between gelatinolytic and collagenolytic activities, we next investigated whether OV19 could degrade native, insoluble collagen, a defining feature of true collagenases capable of hydrolyzing triple-helix collagen. Such activity is common among invasive Vibrio strains and is known to facilitate tissue dissociation, bacterial invasion, and systemic dissemination [34].
To test this, OV19 and selected reference strains (namely V. vulnificus, V. parahaemolyticus, V. harveyi, V. campbellii, and V. splendidus) were individually inoculated into artificial seawater supplemented with insoluble bovine tendon collagen as the sole carbon and energy source. In this nutrient-limited medium, growth was expected to occur only in strains capable of effective collagen hydrolysis, most likely via secreted collagenases.
Over a 48 h monitoring period, OV19 exhibited clear growth and collagen solubilization, with visible bacterial proliferation emerging after approximately 30 h (Figure 3A). The extended lag phase may reflect the low initial inoculum density (10^5^ cfu/mL) and the requirement for gradual protease accumulation before sufficient soluble peptides could support growth. Once established, bacterial multiplication was presumably accompanied by sustained enzyme secretion, accelerating collagen degradation. The process was likely enhanced by the combined activity of collagenases and other secreted proteases. Notably, OV19’s collagenolytic performance was markedly higher than that of the other pathogenic reference strains tested, including V. vulnificus and V. parahaemolyticus, both well-documented collagenase producers [34], as well as other Harveyi clade members (Figure 3B). These findings are consistent with the severe tissue lysis observed in the infected octopus and support a role for OV19’s collagenolytic arsenal in virulence.
We next evaluated motility, another trait frequently linked to Vibrio pathogenicity [35]. Motility assays conducted under controlled conditions revealed that OV19 displayed pronounced swarming motility within a 3 h observation period (Figure 3B. In Vibrio, motility confers ecological advantages in nutrient-limited aquatic environments, besides playing a critical role in host colonization, tissue invasion, and immune evasion [35].
The combination of potent collagenolytic activity and strong motility suggests that OV19 possesses multiple virulence-associated traits that could act synergistically during infection. In the case of the diseased octopus, these features may have enabled the bacterium to promote rapid tissue destruction, bacterial dissemination, and the fulminant septicemia observed. These results support the positioning of OV19 closer to an actively invasive pathogen than to an opportunistic colonizer.
Given these findings, further genomic investigations, particularly whole-genome sequencing, might help unveil the genetic basis of OV19’s enzymatic repertoire, which is of potential interest for subsequent biotechnological exploitation (e.g., production of recombinant enzymes) and its pathogenic potential, though the latter should be experimentally assessed. Moreover, WGS might allow for a more precise identification of the strain, although it should be pointed out once again that the taxonomic classification of vibrios is challenging, mostly due to their high genomic plasticity and recombination rates [8,33].
3. Materials and Methods
3.1. Specimens’ Collection, Bacterial Isolation and Cultivation
Skin and muscle from a wild octopus (Octopus vulgaris) were sampled by cutting with a scalpel, weighed, homogenated in sterile 2% NaCl, and roughly cleared by sedimentation on ice for 20 min. Haemolymph was sampled from the heart using a sterile syringe. Samples were serially diluted in sterile 2% NaCl, then plated on Marine Agar (Condalab, Madrid, Spain) and mTCBS-Agar (modified TCBS) (Condalab, Madrid, Spain) as described in [26].
Single colonies were selected (also based on colony phenotype), isolated and cultivated in Marine Broth (Condalab, Madrid, Spain).
Vibrio type strains were from DSMZ (Braunschweig, Germany). V. parahaemolyticus (DSM10027), V. vulnificus (DSM10143), V. alginolyticus (DSM2171), V. owensii (DSM2165), V. campbellii (DSM19270), V. harveyi (DSM19623), and V. splendidus (DSM19640) were cultivated in Marine Broth.
3.2. Molecular and Bioinformatic Analyses
DNA for PCR amplifications was prepared from single colonies of isolates grown on Marine Agar using the procedure of fast lysis described in [36]. PCR for Vibrio screening were carried out using the Vibrio-specific primers (rpoD_66M13F/rpoD_1592R) described in [26].
Primers employed to amplify genomic loci from Vibrio for MLSA are described in [28]. The universal primers pair 27F/1492R [37] was used to amplify the 16S rDNA. Amplicons were purified using the Gfx PCR DNA Gel Purification Kit (GE Healthcare, Chicago, IL, USA) and sequenced by the Sanger method by Macrogen Europe (Milano, Italy). Following reads trimming, the obtained sequences underwent concatenation [28] and phylogenetic cluster analyses using MEGA11: Molecular Evolutionary Genetics Analysis software, version 11.0, Pennsylvania State University, USA (https://www.megasoftware.net).
The multilocus sequence analysis (MLSA) approach was carried out by concatenating conserved genetic markers for each strain. Concatenated sequences were compared with reference databases to identify the closest related taxa, and a representative dataset was assembled including related species and an appropriate outgroup. Phylogenetic trees were reconstructed using multiple inference methods to ensure consistency and robustness of the resulting topology [28,29,30].
3.3. Screening of Protease-Producing Bacteria
The trypsin-like activity of the supernatant of different isolated Vibrio strains was assessed using the BOC-Gln-Ala-Arg-AMC- (PeptaNova GmbH, Keplerstr, Andhausen, Germany) specific peptides, at concentrations of 0.025 mM [38]. The analysis was carried out in TES buffer, pH 7.4. Unless otherwise stated, reactions were set up in a total volume of 200 μL in microtiter wells and incubated at 37 °C. Fluorescence was measured for 30 min using wavelengths of 355 nm for excitation and 460 nm for emission, in a Biotek Synergy HT microplate reader (BioTek, Winooski, VT, USA). Enzyme-free reactions were used as a negative control and background fluorescence was subtracted from each value. All experiments were conducted in triplicate.
3.4. SDS-PAGE and Zymography
Supernatants from liquid cultures were analyzed in gelatin zymography. Each sample was separated in 7.5% polyacrylamide gel containing 1 mg/mL bovine gelatin, under non-reducing conditions [39]. After electrophoresis, gelatin zymographies were incubated for 24 h at 37 °C in activation buffer containing 2 mM CaCl_2_, 50 mM Tris-HCl buffer, 1.5% Triton X-100, and 0.02% NaN_3_, pH 7.4. After overnight incubation at 37 °C, gel was stained using Coomassie Brilliant Blue G-250. (Sigma Aldrich, Milan, Italy)
3.5. Proteases and Collagenase Activity
Collagenolityc activity was also evaluated by the enzymatic assay of collagenase using Carbobenzoxy-Gly-Pro-Gly-Gly-Pro-Ala-OH as substrate [22,40]. The synthetic peptide Carbobenzoxy-Gly-Pro-GlyGly-Pro-Ala-OH is specifically hydrolyzed by collagenase in two fragments: the Carbobenzoxy-GlyPro-Gly and Gly-Pro-Ala, which reacts with the ninhydrin and can be quantified by a spectroscopic. In this assay 1 U catalyzes the hydrolysis of 1 micromol of Gly-Pro-Ala from Z-GlyPro-Ala (Fluka, Milan, Italy) in 1 min at pH 7.4 and 37 °C. To investigate the proteases activities, we measured them using a modification of the digestion method in which the enzymes were incubated for 5 h with casein (Sigma-Aldrich, St. Louis, MO, USA) at 37 °C. The casein digestion was determined using the colorimetric ninhydrin process. The amino acids released are expressed as micromoles leucine per milligram dry weight of enzyme. One unit equals one micromole of leucine equivalents released from collagen in 5 h at 37 °C, pH 7.5, under the specified conditions.
3.6. Collagen Degradation Test
Bacteria were grown in Marine Broth at 28 °C with orbital shaking overnight, diluted 1:100 in the same medium, and grown up to 1 OD_600_. Exponentially growing cells were diluted 1:5000 in sterile sea water in the presence of 100 mg of insoluble collagen from bovine Achilles tendon (Sigma Aldrich, Milan, Italy) [41], and grown overnight with shaking at 28 °C. The undigested collagen was recovered by centrifugation (4000× g for 5′), washed twice in ddH_2_O, then with 90% ethanol, and then air-dried and weighted. Thereafter, the percentage of digestion was calculated, considering 100 mg as the initial amount.
3.7. Swarming Motility Test
The motility was assessed following the procedure described in [35] with some modifications. Briefly, 8 mL of Marine Agar (MB + 0.5% agarose) were poured in a 60 mm Petri dish, and a micro-well was formed in the center using a pipet plastic tip. 10 μL of cells grown in MB at 30 °C up to 1.5 OD_600_ were put in the micro-well, kept at 25 °C and monitored over time.
4. Conclusions
This study identified the OV19 isolate as a Vibrio strain, member of the Harveyi clade, closely related to V. owensii but likely distinct from any WGS-sequenced strain present in databases. From an evolutionary standpoint, OV19 exemplifies the genetic variability characteristic of the Vibrio genus, in which high recombination rate and genome plasticity drive the emergence of highly adapted and pathogenic phenotypes, which make taxonomic classification challenging.
Comparative biochemical analyses demonstrated that OV19 possesses high collagenolytic and proteolytic activities capable of inducing extensive tissue lysis through efficient degradation of structural proteins—including collagen—thereby facilitating tissue invasion and damage. However, it is worth noting that no experiments specifically designed to demonstrate the actual pathogenicity or virulence of the OV19 strain were conducted in this study. Consequently, its true pathogenic role remains unproven and would require targeted investigations to be confirmed.
Collectively, our findings further highlight the variability of vibrios and the potential of emergence of novel strains harboring unique features, including the production of molecules with high potential of biotechnological exploitation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Baker-Austin C. Oliver J.D. Alam M. Ali A. Waldor M.K. Qadri F. Martinez-Urtaza J. Vibrio spp. infections Nat. Rev. Dis. Primers 20214119 Erratum in Nat. Rev. Dis. Primers 2021, 7, 1510.1038/s 41572-018-0005-830002421 · doi ↗ · pubmed ↗
- 2Baker-Austin C. Trinanes J. Gonzalez-Escalona N. Martinez-Urtaza J. Non-Cholera Vibrios: The microbial barometer of climate change Trends Microbiol.201725768410.1016/j.tim.2016.09.00827843109 · doi ↗ · pubmed ↗
- 3Heidelberg J.F. Eisen J.A. Nelson W.C. Clayton R.A. Gwinn M.L. Dodson R.J. Haft D.H. Hickey E.K. Peterson J.D. Umayam L. DNA sequence of both chromosomes of the cholera pathogen Vibrio cholerae Nature 200040647748310.1038/3502000010952301 PMC 8288016 · doi ↗ · pubmed ↗
- 4Okada K. Iida T. Kita-Tsukamoto K. Honda T. Vibrios commonly possess two chromosomes J. Bacteriol.200518775275710.1128/JB.187.2.752-757.200515629946 PMC 543535 · doi ↗ · pubmed ↗
- 5Burrus V. Waldor M.K. Shaping bacterial genomes with integrative and conjugative elements Res. Microbiol.200415537638610.1016/j.resmic.2004.01.01215207870 · doi ↗ · pubmed ↗
- 6Pant A. Bag S. Saha B. Verma J. Kumar P. Banerjee S. Kumar B. Kumar Y. Desigamani A. Maiti S. Molecular insights into the genome dynamics and interactions between core and acquired genomes of Vibrio cholerae Proc. Natl. Acad. Sci. USA 2020117237622377310.1073/pnas.200628311732873641 PMC 7519391 · doi ↗ · pubmed ↗
- 7Mazel D. Integrons: Agents of bacterial evolution Nat. Rev. Microbiol.2006460862010.1038/nrmicro 146216845431 · doi ↗ · pubmed ↗
- 8Thompson C.C. Vicente A.C.P. Souza R.C. Vasconcelos A.T.R. Vesth T. Alves N. Ussery D.W. Iida T. Thompson F.L. Genomic taxonomy of vibrios BMC Evol. Biol.2009925810.1186/1471-2148-9-25819860885 PMC 2777879 · doi ↗ · pubmed ↗