Comprehensive Genome-Wide Analysis of Dmrt Transcription Factors Reveals Their Roles in Sexual Dimorphism in Scolopendra mutilans
Zhiqiang Li, Jingjing Guo, Ghulam Nabi, Zheng Wang, Buddhi Dayananda, Lin Zhang

TL;DR
This study identifies eight Dmrt transcription factors in the centipede Scolopendra mutilans and explores their roles in sexual dimorphism and sex determination.
Contribution
The study reveals the structural and functional diversity of Dmrt genes and their regulatory mechanisms in a myriapod species.
Findings
Eight Dmrt transcription factors were identified, including five Dsx-related genes and three Dmrt-related genes.
Dsx2 showed tandem duplication on chromosome 4, and Dsx5 was female-biased while Dsx2, Dsx3, and Dsx4 were male-biased.
Sxl and Fem-1C were linked to the regulation of Dsx isoforms, indicating a conserved upstream regulatory cascade.
Abstract
In this study, we identified eight Dmrt transcription factors in the genome of the centipede Scolopendra mutilans. Phylogenetic analysis revealed evolutionary conservation across arthropods, and structural analysis confirmed conserved DM domains and sex-specific motifs, with tan-dem duplication of Dsx2 on chromosome 4. Correlation analysis linked the expression of Sxl and Fem-1C to the regulation of Dsx isoforms, suggesting the presence of a conserved upstream regulatory cascade for sex-specific splicing. The double-sex and mab-3-related transcription factors (Dmrt) are widely distributed in the animal kingdom and play a crucial role in sex determination and differentiation. In this study, we identified eight Dmrt transcription factors in the genome of the centipede Scolopendra mutilans, including five Dsx-related genes (as Dsx1, Dsx2 (five splice variants), Dsx3, Dsx4 and Dsx5) and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
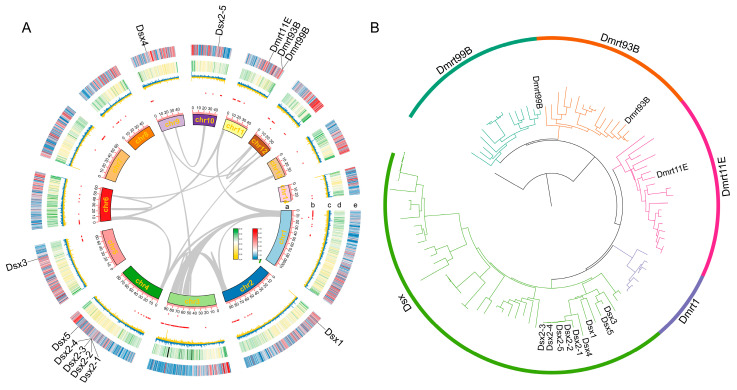 Figure 1
Figure 1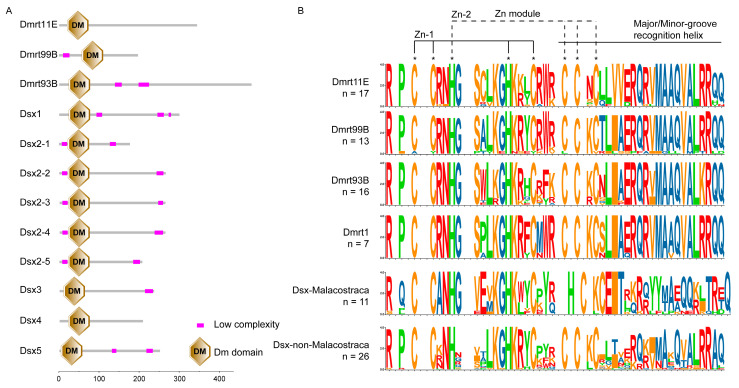 Figure 2
Figure 2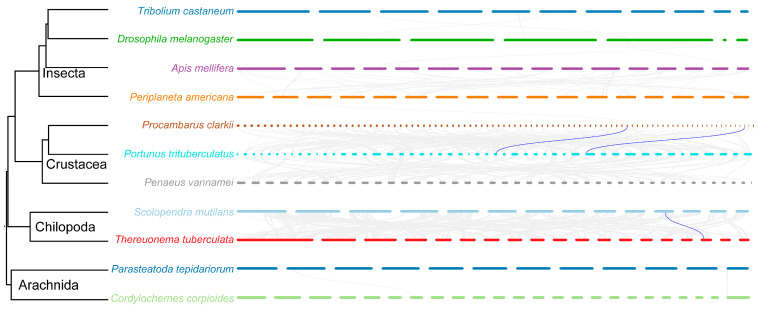 Figure 3
Figure 3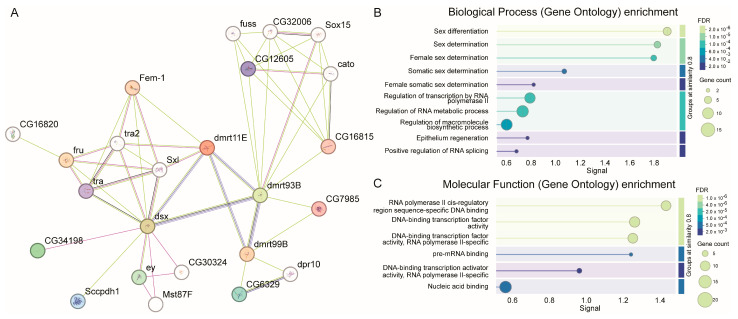 Figure 4
Figure 4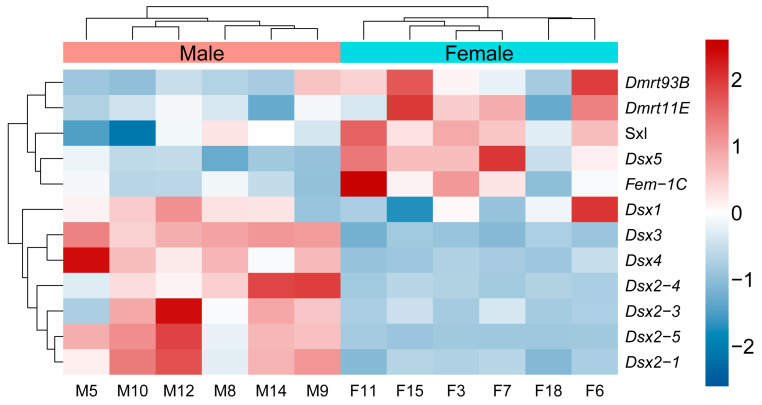 Figure 5
Figure 5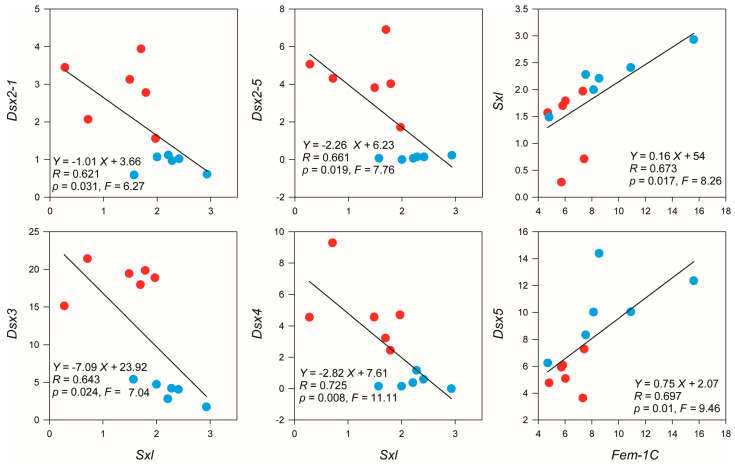 Figure 6
Figure 6- —National Natural Science Foundation of China
- —Key Project of Chinese Medicine Research of Hubei Province
- —Innovation Team of Chinese Medicine Research of Hubei Province
- —Excellent Young and Middle-Aged Science and Technology Innovation Team in Hubei Universities
- —High-Level Talent Project of Hubei University of Chinese Medicine
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic and Clinical Aspects of Sex Determination and Chromosomal Abnormalities · Animal Behavior and Reproduction · Sperm and Testicular Function
1. Introduction
Sexual reproduction is a fundamental phenomenon across the animal kingdom. During reproduction, sex determination is the result of a complex mechanisms primarily regulated by genetic sex determination (GSD), environmental sex determination (ESD), or a combination of both [1]. In ESD species, the environmental factors such as temperature determine the sex during embryonic development [2]. In contrast, GSD species rely on a master sex-determining gene to direct gonadal development into either testes or ovaries [3]. Recent studies have led to the discovery of multiple genes implicated in sexual development, such as double-sex and mab-3-related transcription factor (Dmrt), feminizer (Fem-1), Forkhead box (Fox), SRY-related HMG-box (Sox), signal transducer and activator of the transcription factor (STAT), Sex-lethal (Sxl), transforming growth factor-β family genes (TGF-β), and wingless-type mmtv integration site family (Wnt). Many of these genes demonstrate sexually dimorphic expression patterns within gonadal tissue, underscoring their essential functions in sex determination, gonadal differentiation, and maintenance [4]. In vertebrates, sex differentiation is governed by the female pathway (Rspo1/Wnt/β-catenin/Foxl2) and male pathway (Sry/Sox9/Fgf9/Dmrt1) [5]. In Eumetazoa, Dmrt are evolutionarily conserved components of the sex determination cascade. In Nematoda and Arthropoda, Fem-1 has been functionally linked to regulatory pathways involving Dmrt homologs. In insects, tra (transformer) has been described as the critical regulator of Dmrt sex-specific splicing [6]. In Drosophila melanogaster, the presence of Sxl protein activates tra, generating DsxF, which then produces female traits. In contrast, the absence of Sxl protein generates DsxM, producing male traits [7,8,9].
Dmrt (doublesex and mab-3-related) proteins are a class of transcription factor encoded by homologous genes with Doublesex (Dsx) in D. melanogaster and mab-3 in Caenorhabditis elegans. These proteins are characterized by a conserved double-sex and mab-3 domain, and were first discovered in D. melanogaster [10]. Dmrt are considered master regulators of sex determination and sexual differentiation across the animal kingdom [11,12,13]. The number of Dmrt genes varies across among taxon. In humans, there are eight members (Dmrt1–Dmrt8) [14], in mice, seven (Dmrt1–Dmrt7) [15], and in fish, seven as well (Dmrt1–Dmrt6 plus the paralog Dmrt2b) [16]. Notably, two mammalian-specific Dmrt genes have been identified and designated as Dmrt7 and Dmrt8 [17], highlighting the dynamic evolution of this transcription factor. Phylogenetic analyses of the Dmrt have revealed strongly conserved orthologous groups spanning diverse metazoan lineages. The mammalian Dmrt2/2a [18,19], originally identified in human [20], shares homologous relationships with fish Dmrt2b/terra [21,22], bivalve Dmrt [23], and arthropod Dmrt11E [24]; these orthologous were clustered into the Dmrt2 group. Similarly, the vertebrate Dmrt4/5 [25], arthropod Dmrt99B [26], and nematode Dmd5 [27] were grouped into the Dmrt4/5 group. Interestingly, the invertebrate Dmrt93B [28], vertebrate Dmrt3 [29], and nematode Dmd4 [30] were clustered into the Dmrt93B group. The high conservation of these clusters suggests strong selective pressure to preserve their developmental and reproductive functions [13]. Compared to invertebrates, the members of Dmrt as Dmrt1, Dmrt7 and Dmrt8 are unique to vertebrates [24], whereas Dsx is only restricted to arthropods [31]. The number of Dsx genes varies significantly among arthropod species.
In the teleost fish Oryzias latipes, the Y chromosome-linked gene Dmrt1bY (also known as DMY), a paralogue of Dmrt1, functions as the primary sex-determining factor governing male differentiation, fulfilling a role functionally analogous to that of mammalian Sry [32]. In Xenopus laevis, DMW, the W chromosome-linked paralogue of Dmrt1, acts as a dominant-negative inhibitor of masculinization, suppressing testis differentiation and promoting ovarian development [33]. In pan-arthropod, Dmrt genes are clustered into four groups, designated as Dsx, Dmrt11E, Dmrt93B and Dmrt99B [29]. Dsx-like is the Dsx homolog identified in Eriocheir sinensis [34]. The absence of the DMA domain in a gene, despite its homology to Drosophila Dmrt99B and vertebrate Dmrt5, led to its designation as a Dmrt-like gene [35]. iDMY is the Y-linked Dmrt gene identified in Sagmariasus verreauxi [36].
Scolopendra mutilans Koch (Scolopendromorpha: scolopendridae) is widely distributed across China [37], and represents the most common and largest centipede species in the middle and lower reaches of the Yangtze River. It plays a crucial role in stabilizing the local soil ecosystem [38]. As a medically important invertebrate in China, most studies on S. mutilans have focused on its captive breeding and medicinal properties, including peptide toxin [39,40,41]. Notably, S. mutilans exhibits significant sexual dimorphism (unpublished data). Females generally possess a larger body size (including body length and body width) and display faster growth rates compared to males. Venom production correlates positively with body size in S. subspinipes and S. polymorpha [42], suggesting that female S. mutilans likely holds greater economic and medicinal value.
Beyond their canonical role in sex determination and differentiation, the pleiotropic functions of the evolutionarily conserved Dmrt genes encompass essential roles in embryonic development across invertebrates and vertebrates. These genes regulate a wide range of biological processes during embryo development and sexual differentiation. We hypothesized that differential expression of Dmrt is associated with sexual dimorphism in S. mutilans, potentially leading to transcriptional differences between adult males and females. To test this, we conducted a genome-wide analysis to identify the Dmrt transcription factors in S. mutilans and examined their physicochemical properties, structure features, and phylogenetic relationships. Additionally, we compared the expression profiles of Dmrt transcription factors between the two sexes. This study provides a genomic framework for understanding Dmrt transcription factor in S. mutilans and establishes a foundation for further research on sexual differentiation mechanisms in Myriapoda.
2. Materials and Methods
2.1. Identification and Characterization of Dmrt Transcription Factor in S. mutilans
To identify the potential Dmrt transcription factor members in S. mutilans, we employed the advanced HMMER search (version 3.2.1) using the Hidden Markov Model (HMM) of Dmrt (PF00751) and Basic Local Alignment Search Tool for Proteins (BLASTp) analysis with published Dmrt protein sequences from Bombyx mori, D. melanogaster, Daphnia magna, E. sinensis, Euperipatoides kanangrensis, Glomeris marginata, Procambarus clarkii, Tribolium castaneum, and X. laevis as query sequences (with e-value ≤ 1 × 10^−5^). These analyses were conducted in TBtools v2.142 [43] based on the local S. mutilans genome database [44]. These candidates’ Dmrt transcription factors were further validated by screening conserved domains and motifs using NCBI’s Conserved Domain Database (https://www.ncbi.nlm.nih.gov/cdd, accessed on 23 December 2024), the Simple Modular Architecture Research Tool (http://smart.embl-heidelberg.de/, accessed on 26 December 2024) and the Multiple EM for Motif Elicitation suite (http://meme-suite.org/tools/meme, accessed on 24 December 2024). Subsequently, those conversed domains and motifs were visualized using the Gene Structure View (Advanced) and multiple amino acid sequence alignments functions in TBtools (version 2.357).
To gain insight into the conservation and variation in the DM domain across different species and groups including Dmrt1, Dmrt99B, Dmrt93B, Dmrt11E, Dsx-Malacostraca and Dsx-non-Malacostraca from different species, three-dimensional structures of the DM domains were generated by homology modeling employing SWISS-MODEL server (https://swissmodel.expasy.org/, accessed on 16 Match 2025). The structural alignment and the root mean square deviation (RMSD) calculations were carried out with PyMOL (version 3.1), and all protein molecules’ 3D structural figures were visualized using the same software.
In addition, we determined the several physicochemical properties of all Dmrt transcription factor proteins, including amino acid sequence lengths, molecular weight (MW), isoelectric point (pI), and grand average of hydropathy, employing ProtParam (https://web.expasy.org/protparam/, accessed on 27 December 2024) in TBtools (version 2.357). Then, we employed ProtComp 9.0 (http://www.softberry.com/, accessed on 27 December 2024) to predict the subcellular location of all Dmrt transcription factors.
2.2. Molecular Phylogenetic Analysis
A phylogenetic tree was reconstructed using the Dmrt transcription factor sequences from S. mutilans and others 37 other species, including D. melanogaster, D. magna, and G. marginata (for details, see Supplementary Materials). L-INS-Ioption (the most accurate model) was selected and the MAFFT software (version 7.526) was employed to perform the multiple-sequence alignment and manually trimmed. Then, the maximum-likelihood (ML) phylogenetic tree was conducted employing the IQ-TREE software (version 2.40) with 1000 ultrafast bootstrap replicates to assess branch support [45]. The best-fit substitution model (VT+F+R5) was automatically selected using the “-MPF” option implemented in IQ-TREE. Finally, phylogenetic consensus tree visualization and annotation were carried out in iTOL version 7.2.2 (https://itol.embl.de, accessed on 27 December 2024) and further refined using the itol.toolkit package (version 1.1.7) in R (version 4.3.1) [46].
2.3. Chromosome Locations and Synteny Analysis
The chromosomal location information of the Dmrt transcription factors was derived from the genomic annotation data of S. mutilans. The visualization of the chromosomal distribution pattern of the Dmrt transcription factors was accomplished using TBtools. MCSanX (version 1.0.0, the minimum match size as 5, max gap-size as 25, gap penalty as −1, e-value as 1 × 10^−10^) was employed to carry out collinearity analyses of all Dmrt transcription factors across 11 species genomes, including Apis mellifera, Cordylochernes scorpioides, D. melanogaster, Parasteatoda tepidariorum, Penaeus vannamei, Periplaneta americana, Portunus trituberculatus, P. clarkii, Thereuonema tuberculata, T. castaneum, and S. mutilans [47].
2.4. The PPI Networks of Dmrt Transcription Factors
To generate the protein–protein interaction (PPI) network, we submitted the 12 Dmrt transcription factor sequences to the STRING database (version 12.0, https://string-db.org/ accessed on 21 March 2025), selecting D. melanogaster as the reference organism. Subsequently, functional enrichment analysis was performed within STRING 12.0 to identify the gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway by default parameters.
2.5. The Expression Pattern of Dmrt Transcription Factors in Males and Females
Expression profiles of the Dmrt transcription factors were analyzed using RNA-seq data derived from six male and six female individuals. All reads were aligned to the S. mutilans genome, and gene expression levels were quantified as FPKM values using Cufflinks (version 2.2.1, http://cole-trapnell-lab.github.io/cufflinks, accessed on 12 April 2025). To identify sex-biased expression, we compared transcript levels between the male and female groups. The expression patterns were visualized in the form of a heatmap generated with the pheatmap package (version 1.0.12) in R (version 4.3.1), based on Z-score normalized values across genes.
3. Results
3.1. Structure and Physicochemical Traits of the Dmrt Transcription Factors
In this present study, 12 putative proteins of the Dmrt transcription factors were identified in S. mutilans, including Dmrt11E, Dmrt93B, Dmrt99B, Dsx1, Dsx2 (containing Dsx2-1, Dsx2-2, Dsx2-3, Dsx2-4 and Dsx2-5), Dsx3, Dsx4 and Dsx5 (Figure 1A). All Dmrt transcription factors contained a DM structural domain and a different number of low complexity (LC), such as Dsx1 with 3 LCs, Dsx2 and Dsx5 with 2 LCs (but the location of LCs in Dsxe5 was different from Dsx2), Dsx3 with one LC, and no LCs in Dsx4 (Figure 2A).
The DMRT DM motif was a cysteine-rich DNA-binding domain that included interwoven CCHC and HCCC Zn2+ binding sites, as well as a putative nNLS composed of KGHKK/R (Figure 2B). Protein motif analysis showed that Motif1 and Motif2 existed in all proteins, Motif6 existed only in Dmrt93B, Motif5 and Motif4 existed only in Dmrt11E, Motif7 existed only in Dsx, and Motif9 existed only in Dsx2 and Dsx5 (Figure S1). The 3D structure of the Dmrts in S. mutilans DM domains was found to be highly consistent in spatial folding (Figure S2). The RMSD values among these aligned structures vary between 0.007 and 0.041 (Figures S2 and S3), further supporting the high degree of structural conservation within the Dmrt protein family throughout evolution.
The number of amino acids encoded by the Dmrt ranged from 177 (Dsx2 to 1) to 480 (Dmrt93B), with molecular weights ranging from 19.77 (Dsx2-1) to 52.59 (Dmrt93B) kDa, and the predicted PI ranging from 7.55 (Dsx2-4) to 9.69 (Dmrt11E) (Table 1). All Dmrt members exhibited high hydrophilicity in S. mutilans. These proteins were predicted to localize in the nucleus, except for Dsx5 and Dsx3, which were predicted to be membrane-bound extracellular locations (Table 1).
3.2. Chromosome Locations and Synteny
The Dmrt transcription factors were distributed across chromosomes (Figure 1A), including Dmrt11E, Dmrt99B and Dmrt93B on chromosome 11, but Dsx on different chromosomes, such as Dsx1 on chromosome 2, Dsx2 (expect Dsx2-5 on chromosome 9) and Dsx5 on chromosome 4, Dsx3 on chromosome 5, and Dsx4 on chromosome 10. We employed synteny analysis among 11 arthropods. However, for Dmrt transcription factors, there were only Dmrt11E gene pairs between T. tuberculata and S. mutilans, with a Dmrt93B gene pair between P. clarkii and P. trituberculatus (Figure 3). Furthermore, there were no gene pairs within S. mutilans, and tandem duplications only occurred in Dsx2, except for Dsx2-5 in S. mutilans (Figure 1A).
3.3. Phylogenetic Analysis
To elucidate the phylogenetic relationships among the Dmrt family, we reconstructed a phylogenetic tree using the ML method (Figure 1B). Dmrt99B and Dmrt93B formed a clade whereas Dsx and Dmrt1 formed another clade, which was subsequently associated with Dmrt11E to form a sister clade. The Dsx clade was further divided into four subclades, including the Arachnida Dsx clade, Insecta Dsx clade, Chilopoda Dsx clade, and Crustacea Dsx clade. S. mutilans and G. marginata were clustered with T. tuberculata in Dmrt99B. Dmrt93B from G. marginata, E. kanangrensis, P. chinensis, S. mutilans and T. tuberculata formed a subclade. In the Dmrt11e clade, centipedes, including S. mutilans and T. tuberculata, were grouped together, highlighting lineage-specific conservation.
3.4. The PPI Networks of Dmrt Transcription Factors
We examined 12 putative Dmrt transcription factor sequences in the STRING database and identified 12 matched proteins, which were classified into four protein groups. The PPI network comprised 24 nodes and 54 edges with an average node degree of 4.5, and an ap-value of PPI enrichment of 5.96 × 10^−10^ (Figure 4A). GO enrichment analysis indicated that these proteins were primarily associated with biological processes such as sex differentiation (GO:0007548), sex determination (GO:0007530), female sex determination (GO:0030237), somatic sex determination (GO:0018993) and female somatic sex determination (GO:0019101) (Figure 4B). For molecular function, the proteins were enriched in DNA-binding transcription activator activity, RNA polymerase II-specific (GO:0001228), RNA polymerase II cis-regulatory region sequence-specific DNA binding (GO:0000978), DNA-binding transcription factor activity (GO:0003700), DNA-binding transcription factor activity, RNA polymerase II-specific (GO:0000981), pre-mRNA binding (GO:0036002), and nucleic acid binding (GO:0003676) for molecular function (Figure 4C). Additionally, the male semi-fertile phenotype (FBcv:0000006) in Drosophila is associated with Dmrt11E and Dsx, highlight the functional relevance of these transcription factors in sex determination.
3.5. Expression Profile of Dmrt Transcription Factors in Individuals of Both Sexes
Dsx99B in S. milantus demonstrated no expression (PFKM = 0). However, Dsx5 exhibited a substantially higher expression level in females compared to males (p < 0.01, LogFC = −0.90), while Dsx2-1 (p < 0.01, LogFC = 1.65), Dsx2-3 (p < 0.05, LogFC = 3.30), Dsx2-4 (p < 0.01, LogFC = 4.14), Dsx2-5 (p < 0.001, LogFC = 5.31), Dsx3 (p < 0.001, LogFC = 2.30), and Dsx4 (p < 0.001, LogFC = 3.55) showed a pronounced increase in expression levels in males relative to females (Figure 5). The expression level indicated a negative correlation between sxl and Dsx2-1 (F = 6.265, R = 0.621, p < 0.05; Figure 6), between sxl and Dsx2-5 (F = 7.757, R = 0.661, p < 0.05; Figure 6), between Sxl and Dsx3 (F1,10 = 7.039, R = 0.643, p < 0.05; Figure 6), and between sxl and Dsx4 (F1,10 = 11.11, R = 0.725, p < 0.01; Figure 6). The expression of Fem-1C and Dsx5 was positively related (F1,10 = 9.462, R = 0.697, p < 0.05; Figure 6), and a similar trend was reported between the expression of Fem-1C and Sxl (F1,10 = 8.261, R = 0.673, p < 0.05; Figure 6).
4. Discussion
We identified eight members among the Dmrt transcription factors members in S. mutilans (Figure 1A), located on six chromosomes, including a remarkable tandem duplicate of the Dsx2 gene (Dsx2-1 to Dsx2-5) on chromosome 4, except for Dsx2-5. We reported the Dmrt11E gene pair between T. tuberculata and S. mutilans; however, no gene pairs were found within S. mutilans. The expression analysis uncovered sexually dimorphic patterns for several key genes: Dmrt11E was female-biased, while multiple Dsx isoforms exhibited either male- (Dsx2, Dsx3, Dsx4) or female-biased (Dsx5) expression.
The Dmrt93B cluster includes orthologs such as vertebrate Dmrt3 [29] and Dmd-4 [30]. Dmrt3 is typically associated with testicular development and embryogenesis [16,48,49,50,51,52]. However, in S. mutilans, the Dmrt93B transcript levels were strikingly low in both sexes, suggesting a divergent regulatory role compared to other species. Sexual dimorphism is also observed in the Dmrt99B gene. While it is testis-biased in Scylla paramamosain and Macrobrachium rosenbergii, consistent with the typical role of Dmrt99B genes in male development [26,53], it is female-specific in Daphnia magna and Bombyx mori [54,55]. Intriguingly, Dmrt99B was undetectable in S. mutilans transcriptome data, raising questions about whether this reflects species-specific gene loss or functional replacement by another gene, which warrants further investigation.
Dmrt11E is an important gene in gonadal maturation; however, the expression profile of the Dmrt11E gene is complex. The function of Dmrt11E is similar to Dmrt2 [24], playing a key role in the male reproductive physiology of many species. In S. mutilans, Dmrt11E was highly expressed in females compared to males, a pattern similar to D. magna [56] but opposite to that observed in M. rosenbergii [26] and M. nipponense [57]. This striking divergence suggests that the functional role of Dmrt11E is not conserved across the arthropod phylogeny and may have been co-opted for female-specific functions in certain lineages, including myriapods and branchiopods.
Dsx undergoes sex-specific alternative splicing to produce male (DsxM) and female (DsxF) isoforms, which play a crucial role in controlling sexual dimorphism. While this core mechanism is conserved, its regulation and complexity exhibit remarkable lineage-specific variation. The number of alternative transcripts differs among insects; the default splicing pattern in the absence of regulators can be male in D. melanogaster or female in Antheraea mylitta and Bombyx mori [58,59,60,61]. Insect and crustacean studies suggest that upstream regulators like Sxl and Tra-2 may also interact with Dmrt genes in a species-specific manner [62,63,64,65,66,67]. Given the evolutionary context, where the core function of Dsx is conserved but its regulatory inputs and molecular details are flexible, we hypothesized that S. mutilans similarly utilizes sex-specific Dsx isoforms. The results supported this hypothesis, leading us to designate the male-biased isoforms as SmDsxM and the female-biased Dsx5 as SmDsxF.
In D. magna, Dsx1 is essential for the development of male-specific traits [68]. However, Dsx expression varies significantly among species. In Cherax quadricarinatus [69] and S. paramamosain [70], Dsx expression is significantly higher in the ovary than in the testis. Conversely, in Penaeus monodon, Dsx expression is significantly lower in the ovary [13]. Our results revealed that Dsx expression levels were generally higher in males than in females in S. mutilans, except for Dsx5 (Figure 5). These results suggest that Dsx may be functionally divergent, with some isoforms promoting male-specific traits and others regulating female-specific development.
In S. mutilans, we observed a sexually dimorphic expression pattern of Sxl and Fem-1C, with significantly higher expression levels in females than in males. Correlation analysis revealed significant associations between Sxl and Dsx, Fem-1C and Dsx, and Sxl and Fem-1C (Figure 6), suggesting a potential functional relationship among these sex determination genes. Notably, the expression pattern of Fem-1C in S. mutilans resembles that reported in Dendroctonus armandi [71]. The molecular mechanism of sex determination involving these genes appears to follow a conserved regulatory cascade, where the spliced form of the Fem protein regulates the alternative splicing of dsx transcripts, while the un-spliced Fem remains non-functional and fails to process Dsx [72]. This splicing cascade is crucial for proper sexual differentiation in honey bees [73]. Based on the expression profiling, we classified Dsx2, Dsx3, and Dsx4 as male-biased Dsx isoforms (renamed as SmDsxM), while Dsx5 was identified as a female-biased isoform (renamed as SmDsxF). The results suggest that Fem-1C may play a dual regulatory role in S. mutilans, potentially up-regulating SmDsxF expression while down-regulating SmDsxM, similar to the regulatory mechanisms observed in other arthropods.
5. Conclusions
In this study, we investigated the Dmrt transcription factors in S. mutilans and examined their potential roles in sexual determination and differentiation. We identified eight Dmrt genes (Dsx1, Dsx2 (with five different alternative splices), Dsx3, Dsx4, Dsx5, Dmrt11E, Dmrt99B, and Dmrt93B). Based on their sexually dimorphic expression patterns, we proposed a sex-specific regulatory model. Our results reveal that terminal Dsx signal itself can be complex, the results supporting a model of sexual differentiation driven by the balance between SmDsxM and SmDsxF. Dsx5 (renamed SmDsxF) was significantly upregulated in females. In contrast, Dsx2, Dsx3, and Dsx4 (collectively renamed as SmDsxM) exhibited male-biased expression. These findings provide a genomic framework for future functional analyses aimed at understanding Dmrt-mediated sexual development in S. mutilans, and highlight both conserved and divergent mechanisms within arthropods.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bachtrog D. Mank J.E. Peichel C.L. Kirkpatrick M. Otto S.P. Ashman T.-L. Hahn M.W. Kitano J. Mayrose I. Ming R. Sex determination: Why so many ways of doing it?P Lo S Biol.201412 e 100189910.1371/journal.pbio.100189924983465 PMC 4077654 · doi ↗ · pubmed ↗
- 2Liu T. Li R. Liu L. Wu S. Zhang L. Li Y. Wei H. Shu Y. Yang Y. Wang S. The effect of temperature on gonadal sex differentiation of yesso scallop Patinopecten yessoensis Front. Cell. Dev. Biol.2022980304610.3389/fcell.2021.80304635174162 PMC 8841428 · doi ↗ · pubmed ↗
- 3Carter A.W. Paizt R.T. Kohno S. Sex determination and differentiation in reptiles Hormones and Reproduction of Vertebrates 2nd ed. Norris D.O. Lopez K.H. Academic Press Cambridge, MA, USA 2024133
- 4Zhang Q. Huang J. Fu Y. Chen J. Wang W. Genome-wide identification and expression profiles of sex-related gene families in the Pacific abalone Haliotis discus hannai Comp. Biochem. Physiol. Part D Genom. Proteom.20245010120510.1016/j.cbd.2024.10120538364653 · doi ↗ · pubmed ↗
- 5Nagahama Y. Chakraborty T. Paul-Prasanth B. Ohta K. Nakamura M. Sex determination, gonadal sex differentiation, and plasticity in vertebrate species Physiol. Rev.20211011237130810.1152/physrev.00044.201933180655 · doi ↗ · pubmed ↗
- 6Peng W. Zheng W. Handler A.M. Zhang H. The role of the transformer gene in sex determination and reproduction in the tephritid fruit fly, Bactrocera dorsalis (Hendel)Genetica 201514371772710.1007/s 10709-015-9869-726481008 · doi ↗ · pubmed ↗
- 7Bell L.R. Maine E.M. Schedl P. Cline T.W. Sex-lethal, a Drosophila sex determination switch gene, exhibits sex-specific RNA splicing and sequence similarity to RNA binding proteins Cell 1988551037104610.1016/0092-8674(88)90248-63144435 · doi ↗ · pubmed ↗
- 8Mac Dougall C. Harbison D. Bownes M. The developmental consequences of alternative splicing in sex determination and differentiation in Drosophila Dev. Biol.199517235337610.1006/dbio.1995.80478612957 · doi ↗ · pubmed ↗