Schizochytrium Supplementation in Compound Feed: Effects on Growth, Metamorphosis, Intermediate Metabolism, and Intestinal Health of Bullfrogs (Lithobates catesbeianus)
Hao Ding, Yinglin He, Yujian Song, Jingjing Liang, Woxing Li, Chao Xu, Huirong Yang

TL;DR
Adding 15 g/kg of Schizochytrium to bullfrog feed improves growth, metamorphosis, digestion, and intestinal health by enhancing key biological processes.
Contribution
Identifies optimal Schizochytrium meal levels for bullfrog growth and intestinal health through comprehensive metabolic and physiological analysis.
Findings
A 15 g/kg Schizochytrium meal increased weight gain by 23.78% and improved metamorphosis rates.
Intestinal enzyme activity and microbial diversity were highest at 15 g/kg Schizochytrium.
Key genes related to growth and intestinal function were upregulated with 15 g/kg Schizochytrium.
Abstract
Schizochytrium is often added to feed to enhance the growth and health of farmed animals, yet research on its effects on amphibians remains relatively scarce. Here, this study investigated the effects of dietary Schizochytrium meal on growth, metamorphosis, intermediate metabolism, and intestinal health of bullfrogs. Six compound feeds (S0–S5) containing different gradients of Schizochytrium meal (0.00, 2.00, 5.00, 10.00, 15.00, and 20.00 g/kg diets) were formulated. After 90 days, the S4 group (15.00 g/kg) exhibited significantly superior growth performance, with the weight gain rate (WGR) increasing by up to 23.78% compared to the control (S0). Metamorphosis rate (MR) peaked at 23.33% in the S4 group. The enzyme activities of digestion (amylase (AMS), lipase (LPS), protease), brush border membrane (Na+, K+-ATPase, alkaline phosphatase (AKP), γ-glutamyl transferase (γ-GT), creatine…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1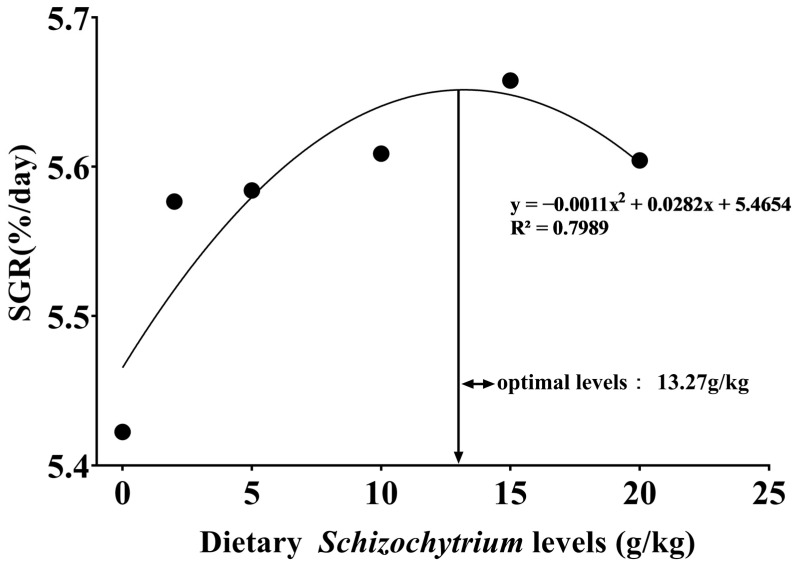 Figure 2
Figure 2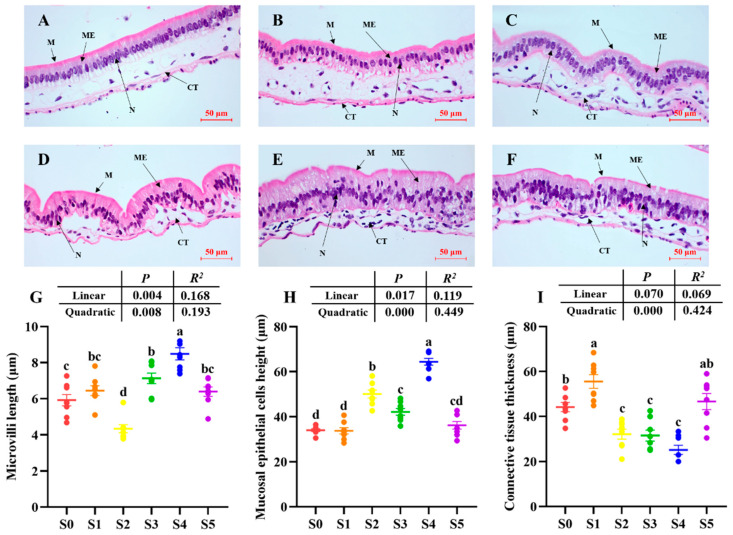 Figure 3
Figure 3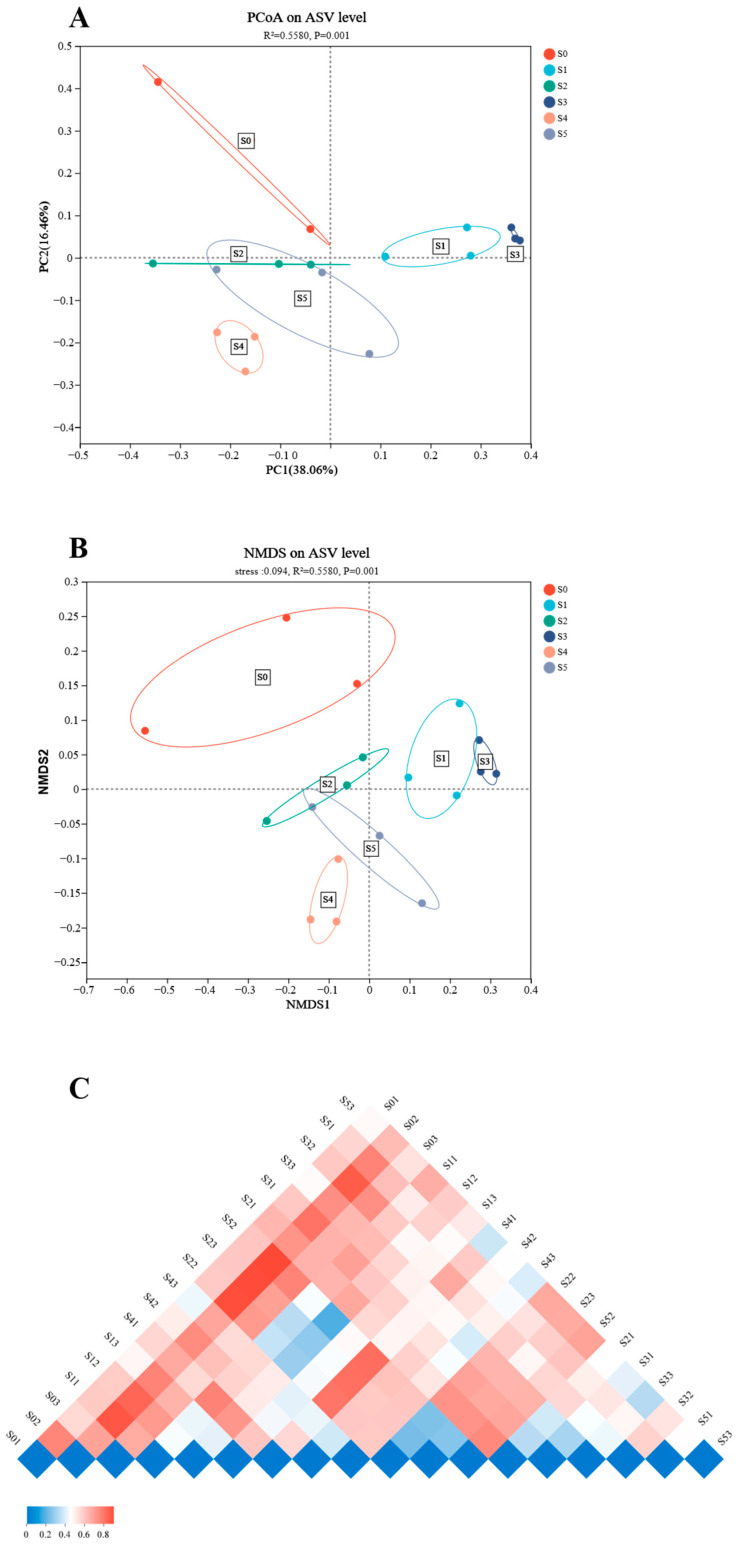 Figure 4
Figure 4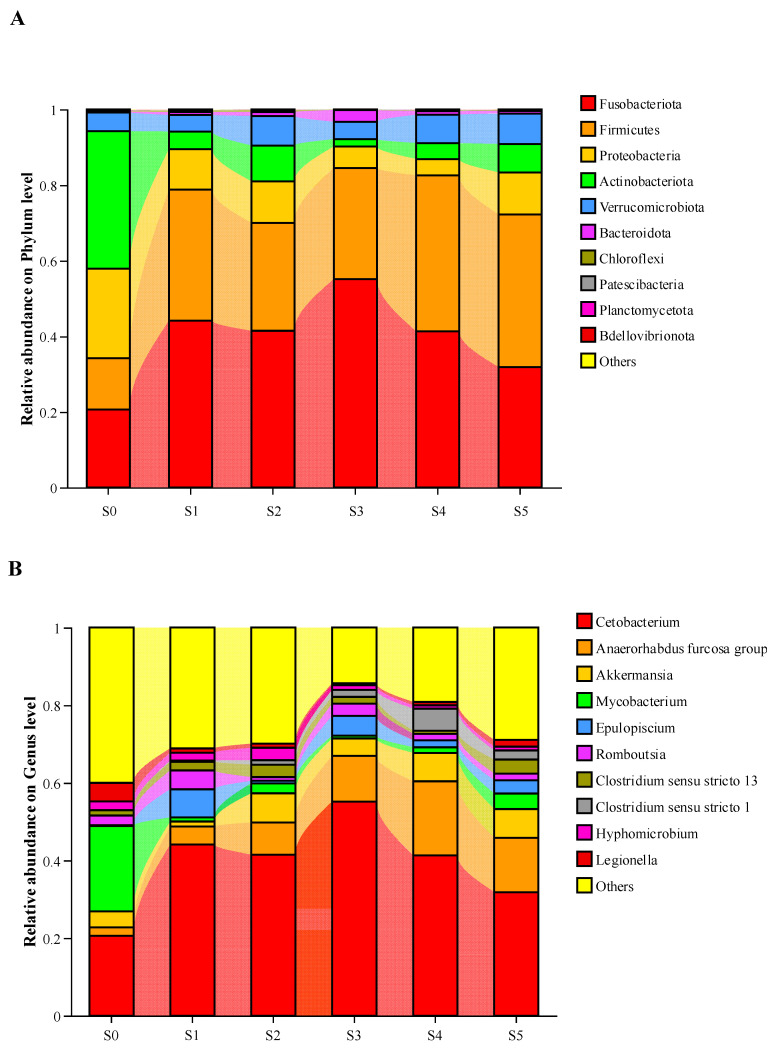 Figure 5
Figure 5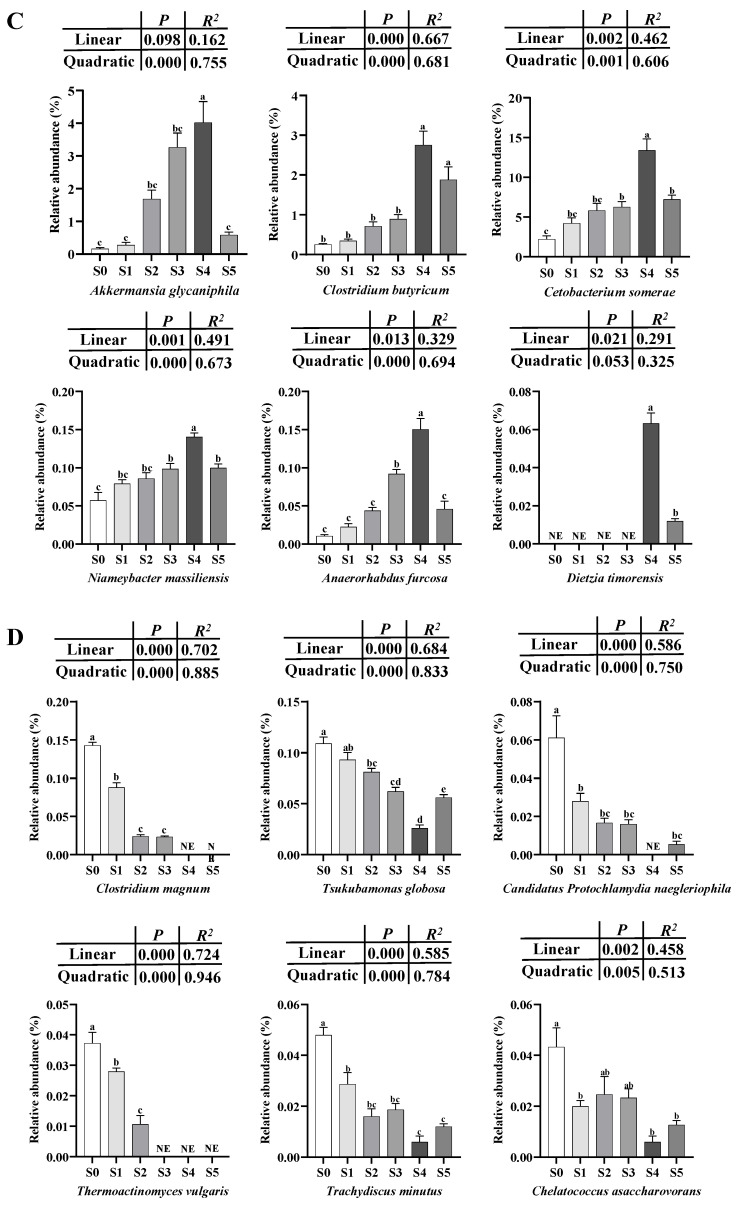 Figure 6
Figure 6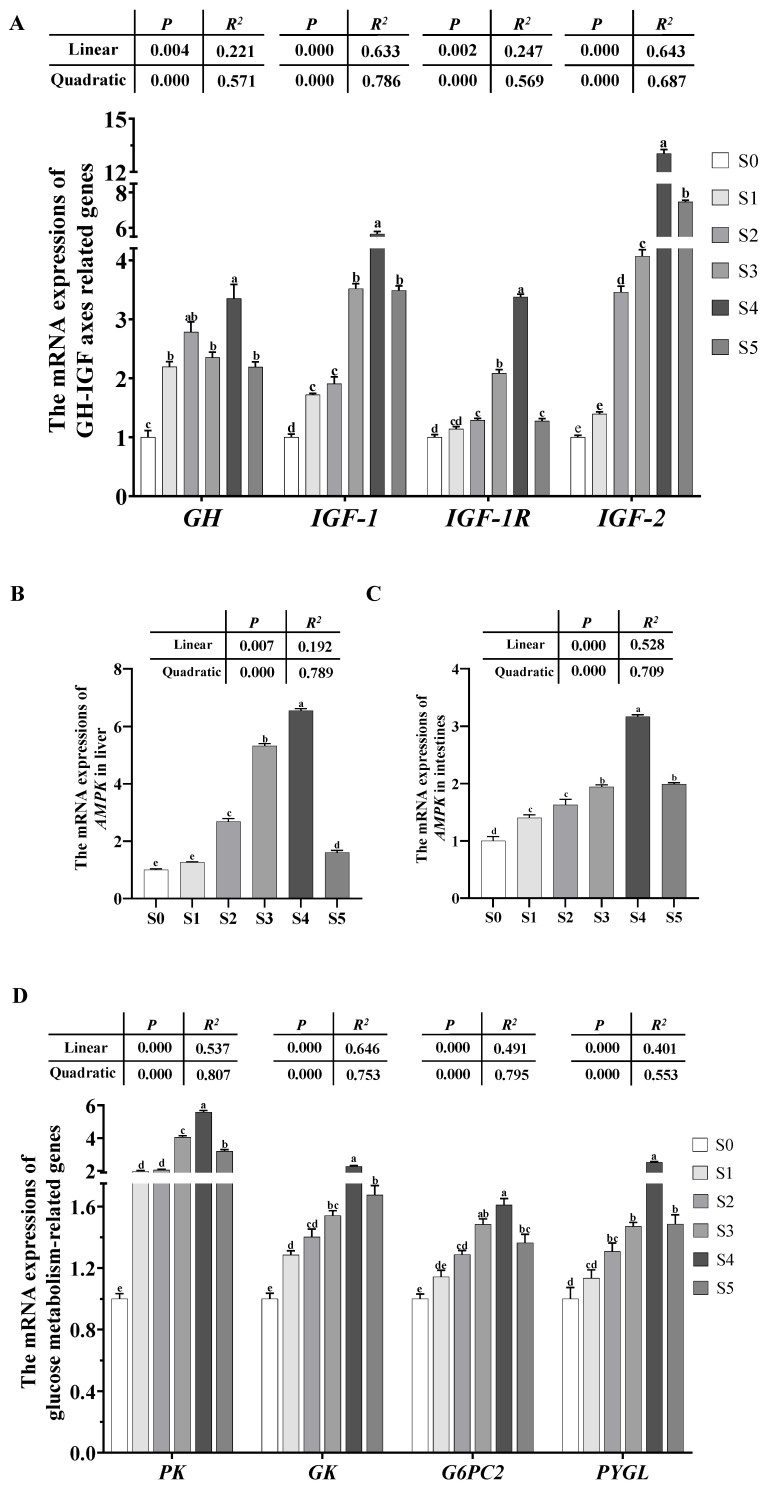 Figure 7
Figure 7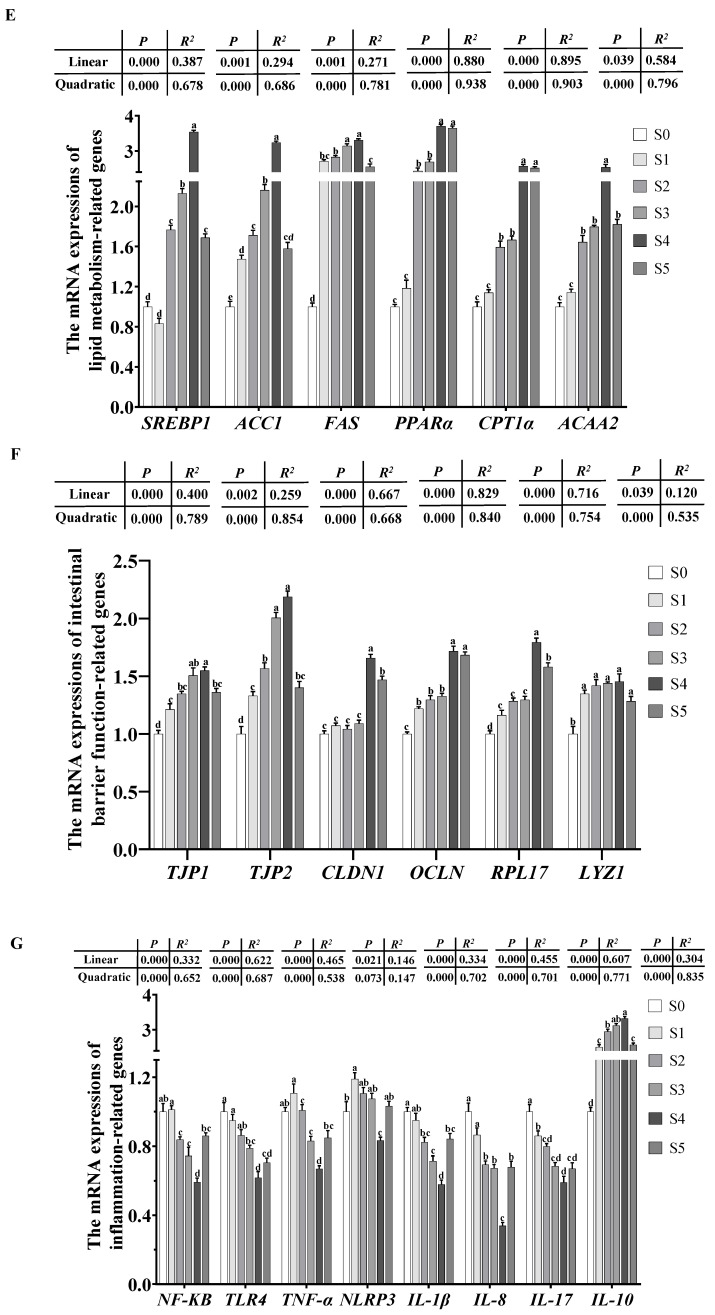 Figure 8
Figure 8- —Guangdong Forestry Science and Technology Innovation Project
- —Provincial Science and Technology Special Fund Project for Zhongshan City (Major Special Project + Task List Management Mode)
- —Agricultural Product Quality and Safety Risk Assessment Project Ministry of Agriculture and Rural Affairs of the People’s Republic of China
- —The second round of rural science and technology special correspondent project of the Hundred-Thousand-Ten-Thousand Project
- —Natural Science Foundation of Guangdong Province
- —Special Fund Project for Marine Economic Development (Six Marine Industries) of Guangdong Province
- —Key-Area Research and Development Program of Guangdong Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGut microbiota and health · Pharmaceutical and Antibiotic Environmental Impacts
1. Introduction
With the increasing demand of consumers for healthy white meat products, bullfrog (Lithobates catesbeianus) farming has great market potential. A critical stage in bullfrog production is tadpole rearing, as it directly influences the health and growth performance of subsequent juvenile frogs [1]. This phase is characterized by metamorphosis, a complex developmental process involving the transformation of aquatic tadpoles into amphibious frogs, marked by fundamental remodeling of tissues and organs such as the development of limbs and lungs [2,3,4]. Therefore, the efficiency of this metamorphic process is a key determinant of overall breeding success, highlighting the need for nutritional strategies to support tadpole development.
Food nutrition is a key factor to guarantee high breeding efficiency of farmed animals. The marine microalgae Schizochytrium has emerged as a promising aquafeed ingredient, renowned for its high content of polyunsaturated fatty acids (PUFAs), particularly docosahexaenoic acid (DHA) [5]. This microalga is commonly used as a live food source for the larval and juvenile stages of aquatic animals due to its high nutritional value and suitable cell size [6]. Its nutritional value has been well-documented in various fish species, where dietary supplementation has been shown to improve growth performance, feed utilization, stress tolerance, and disease resistance [7,8]. Specifically, dietary supplementation with Schizochytrium significantly reduced the feed conversion ratio to 1.20% in golden pompano (Trachinotus ovatus) and zebrafish (Danio rerio), while also improving non-specific immunity and increasing the abundance of beneficial intestinal bacteria [9,10]. In Nile tilapia (Oreochromis niloticus), Schizochytrium completely replaced fish oil in feed, enhancing weight gain, feed efficiency, protein efficiency, and optimizing the DHA:EPA ratio in filets, demonstrating that Schizochytrium modulates growth performance and fatty acid metabolism in aquatic species [11]. Furthermore, Schizochytrium upregulates the expression of antioxidant and immune-related genes in rainbow trout (Oncorhynchus mykiss) [12]. However, the existing body of evidence is predominantly derived from fish, and direct evidence for its application in amphibians is currently lacking.
The role of DHA in amphibian development, particularly during the critical window of metamorphosis, warrants greater attention. As in other vertebrates, DHA is recognized for its crucial role in neural development and visual function in amphibians [13,14,15]. Adequate lipid nutrition, including a balanced provision of essential fatty acids, is vital to support the extensive tissue remodeling and high energy demands characteristic of metamorphosis [16], but the specific requirements and optimal levels of fatty acids for amphibians are not well-defined. Most existing inferences are drawn from fish or general vertebrate models [17].
Beyond systemic growth, intestinal health is paramount for nutrient acquisition and overall development. A well-functioning intestine, supported by a balanced interaction between the gut microbiota, epithelial barrier, and immune system, is essential for efficient digestion and absorption [18,19,20,21]. In addition, the enzyme activity in the intestine is the reliable indicator for evaluating intestinal absorption and digestion capacity, due to the fact that these enzymes affect the ability of animals to obtain and utilize nutrients in food [22,23]. While the positive impacts of Schizochytrium on intestinal health have been observed in fish, its influence on the gut function of amphibian tadpoles, particularly during the metabolically demanding period of metamorphosis, remains uninvestigated.
To address these research gaps, the present study was designed to systematically evaluate the effects of dietary Schizochytrium meal supplementation on growth, metamorphosis, intermediary metabolism, and intestinal health in bullfrog tadpoles over a 90-day feeding trial. We hypothesized that dietary Schizochytrium meals would promote tadpole growth and metamorphosis by enhancing digestive function, modulating key metabolic pathways, and improving intestinal health. The findings are expected to provide novel insights into the application of Schizochytrium in amphibian aquaculture and contribute to developing effective nutritional strategies for improving bullfrog breeding efficiency.
2. Materials and Methods
2.1. Ethics Statement
The Animal Care and Use Committee in South China Agricultural University have approved the present study (Permit No.: 2017D006).
2.2. Diets, Bullfrog, and Sampling
Schizochytrium meal was obtained from Guanxing Agricultural Technology Co., Ltd. (Guangzhou, China). Graded doses of Schizochytrium meal (0, 2.00, 5.00, 10.00, 15.00, and 20.00 g/kg) were added to the basal diet (Table 1). Schizochytrium meal doses were selected according to the research on Ictalurus punctatus and Lateolabrax maculatus [24,25]. The diets were made by a granulation machine (Beijing Modern Yanggong Machinery Sand Development Co., Ltd., Beijing, China).
Bullfrog tadpoles were obtained from Guanxing Agricultural Technology Co., Ltd. (Guangzhou, China). A total of 720 tadpoles (0.04 ± 0.00 g) were dispersed into 18 indoor tanks (40 L) with equal individuals (40/tank). Temperature, dissolved oxygen, pH, and ammonia–nitrogen were monitored daily and maintained within optimal ranges for bullfrog tadpole farming [26]. Six compound feeds (S0–S5) were fed to tadpoles four times a day (08:30, 12:00, 17:30, and 22:00). After 90 days, total number, weight, and body length of bullfrogs in each tank were recorded for the analysis of weight gain rate, specific growth rate, and condition factor. Number of tadpoles with hind limb buds was recorded for the analysis of post-premetamorphosis rate (PPR) and metamorphosis rate (MMR). A total of 10 tadpoles from each tank were sampled for the analysis of proximate composition. Four tadpoles per tank were anesthetized via MS-222 at 100 mg/L, and brain, liver, and intestine samples were obtained for PCR and biochemical analysis. In addition, four tadpoles from each tank were anesthetized to collect intestine samples for the analysis of intestinal microbiota and hematoxylin–eosin (HE) staining.
2.3. Analytic Procedures
2.3.1. Analysis of Proximate Composition and Intestinal Histomorphometry
The proximate composition of compound feeds and tadpoles were determined by AOAC protocols [15]. Intestinal HE staining was carried out via the methods of Xu et al. [16]. In short, middle intestine samples were sectioned using a slicing machine (RD-2230, Shenyang Roundfin Technology, Co., Ltd., Shenyang, China) and were then stained by hematoxylin and eosin (HE) staining kit (NO. S20202-2×100, Shanghai Shangbao Biotechnology Co., Ltd., Shanghai, China). The images were obtained via a light-polarizing microscope (BX53P, Olympus, Tokyo, Japan).
2.3.2. Analysis of Intestinal Biochemical Indicators and Microbiota
Intestinal amylase (AMS, C016-1-1), lipase (LPS, A054-2-1), protease (A080-1-1), Na^+^, K^+^-ATPase (A070-2), alkaline phosphatase (AKP, A059-2), γ-glutamyl transferase (γ-GT, C017-2-1), creatine kinase (CK, A032-1-1), superoxide dismutase (SOD, A001-3), catalase (CAT, A007-1-1), total antioxidant capacity (T-AOC, A015-3-1), and malondialdehyde (MDA, A003-1) were detected using commercial kits (Jiancheng Biotech, Co., Ltd., Nanjing, China).
Bacterial DNA was extracted from intestine via the QiAamp DNA stool Mini Kit (Qiagen, Dusseldorf, NRW, Germany). DNA quality and concentration were assessed with a NanoDrop2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). The V3-V4 regions of the bacterial 16S rRNA gene were amplified using primers 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′). PCR was performed with denaturation at 94 °C for 2 min, followed by 30 cycles (98 °C for 10 s, 55 °C for 30 s, 68 °C for 30 s) and a final extension at 68 °C for 5 min. Purified amplicons were sequenced using a 2 × 300 bp paired-end format on the Illumina MiSeq platform (Illumina, San Diego, CA, USA). The sequence data have been deposited at NCBI GenBank database under accession number PRJNA1335207.
The qualified raw sequence data was split based on barcodes and the primer and adapter sequences were removed using the cutadapt plugin in QIIME2 (version 2024). Paired-end reads were merged based on their overlap relationship to obtain optimized data after quality control. Chimera sequences were removed and clustered into amplicon sequence variants (ASVs) using DADA2 (version 2024). A total of 1,286,881 raw sequences were obtained from bacterial samples, and subsequent processing of high-quality reads identified 2491 bacterial ASVs. Species annotation was performed using the classifier-sklearn plugin with the Silva-138 reference database (https://www.arb-silva.de/, accessed on 15 August 2025). Community structure at both the phylum and genus levels was analyzed using Python software (version 2.7). Alpha diversity indices were calculated using QIIME2. Beta diversity was analyzed with principal coordinate analysis (PCoA) and non-metric multidimensional scaling (NMDS) based on Bray–Curtis dissimilarity using Vegan package (version 2.5-3), with significance tested through permutation multivariate analysis of variance (Adonis).
2.3.3. Analysis of Polymerase Chain Reaction (PCR)
PCR analysis was also performed following the methods previously described by Xu et al. [16]. Briefly, total RNA was extracted from brain, liver, and intestine samples by Total RNA Isolation Kit (Vazyme Biotech Co., Ltd., Nanjing, China). The cDNA was synthesized using the HiScript IV 1st Strand cDNA Synthesis Kit (Vazyme, Nanjing, China) and T100 thermal cycler (Bio-rad, Hercules, CA, USA). RT-qPCR assays were carried out on a CFX Duet RT-PCR System (Bio-rad, Hercules, CA, USA) using Taq Pro Universal SYBR qPCR Master Mix (Vazyme, Nanjing, China). The primers of the target genes are presented in Table 2. β-actin was used as the internal reference gene, and the relative expression levels of the target gene were determined using the 2^−ΔΔCT^ method [27].
2.4. Statistical Analysis
All statistical analyses were performed through SPSS program (version 22.0). The normality of the data was confirmed using the Shapiro–Wilk test, and the homogeneity of variances was confirmed using Levene’s test. One-way ANOVA (SPSS program version 22.0) followed by Tukey’s multiple comparison test was used to identify differences among all experimental groups. Data are presented as mean ± standard error (mean ± S.E.M) of the mean, and p < 0.05 is significantly different.
3. Results
3.1. Growth Performance, Feed Utilization, and Metamorphosis Rate
The condition factor, whole-body moisture, and ash contents showed no significant differences (p > 0.05) among all groups (Table 3). Final weight, weight gain rate (WGR) (Figure 1), specific growth rate (SGR), feed intake (FI), PPR, and MR increased significantly (p < 0.05) as the Schizochytrium levels increased to 15.00 g/kg diet but decreased with further increasing Schizochytrium levels. Feed conversion ratio (FCR) declined obviously (p < 0.05) as Schizochytrium levels increased to 15.00 g/kg diet but increased with further increasing Schizochytrium levels. The highest contents of whole-body crude protein and lipids were observed in diets S5 and S3, respectively. Based on the quadratic regression analysis of SGR against Schizochytrium levels, optimal Schizochytrium levels for bullfrog tadpoles were estimated to be 13.27 g/kg diet (Figure 2).
3.2. Intestinal Biochemical Indicators and Histomorphometry
Total antioxidant capacity showed no significant differences (p > 0.05) among all groups (Table 4). The activities of AMS, LPS, PES, Na^+^, K^+^-ATPase, AKP, γ-GT, CK, SOD, and CAT, as well as microvilli length and mucosal epithelial cell height in the intestine increased obviously (p < 0.05) as the Schizochytrium levels increased to 15.00 g/kg diet but decreased with further increasing Schizochytrium levels. Malondialdehyde contents and connective tissue thickness in the intestine declined obviously (p < 0.05) as Schizochytrium levels increased to 15.00 g/kg diet but increased with further increasing Schizochytrium levels (Figure 3).
3.3. Analyses of Intestinal Microbiota
Diversity analysis indicated that both α-diversity and β-diversity were higher in the S4 group. The values of Shannon, Simpson, Shannoneven, Simpsoneven, and phylogenetic diversity showed no significant differences (p > 0.05) among all groups (Table 5 and Figure 4). The values of ASV, Ace, and Chao1 increased obviously (p < 0.05) as the Schizochytrium levels raised to 15.00 g/kg diet but decreased with further increasing Schizochytrium levels.
Intestinal bacterial composition at the phylum, genus, and species levels is presented in Figure 5. On the phyla level (Figure 5A), Fusobacteriota, Firmicutes, Verrucomicrobiota, and Bacteroidota in tadpoles fed diets with Schizochytrium were all more abundant than those of the S0 group, but the opposite trend was found in Proteobacteria, Actinobacteriota, Chloroflexi, Patescibacteria, Planctomycetota, and Bdellovibrionota. The abundances of Firmicutes increased as the Schizochytrium levels increased to 15.00 g/kg diet but decreased with further increasing Schizochytrium levels. Meanwhile, fish fed the S4 diet have the highest values of Verrucomicrobiota.
On the genus level (Figure 5B), the abundances of Cetobacterium, Anaerorhabdus furcosa, Epulopiscium, and Clostridium sensu stricto 1 in tadpoles fed diets with Schizochytrium were higher than those of the S0 group, but the opposite trend was found in Legionella. As for the supplementation of Schizochytrium, the abundances of Anaerorhabdus furcosa and Clostridium sensu stricto 1 increased with the increasing levels up to 15.00 g/kg. The lowest values of Hyphomicrobium were seen in the S4 group.
On the species level (Figure 5C,D), the abundances of Clostridium magunm, Tsukubamonas globosa, Candidatus Protochlamydia naegleriophila, Thermooactinomyces vulgaris, Trachydiscus minutus, and Chelatococcus asaccharovorans of the S0 group were relatively higher than those of Schizochytrium treatments, but the opposite trend was found in Akkermansia glycanispila, Clostridium butyricum, Cetobacterium somerae, Niameybacter massiliensis, Anaerorhabdus furcosa, and Dietzia timorensis. As for the supplementation of Schizochytrium, the abundances of Clostridium magunm and Thermooactinomyces vulgaris declined obviously (p < 0.05) with the levels rising up to 20.00 g/kg. The abundances of Akkermansia glycanispila, Clostridium butyricum, Cetobacterium somerae, Niameybacter massiliensis, Anaerorhabdus furcosa, and Dietzia timorensis increased obviously (p < 0.05) as the Schizochytrium levels raised to 15.00 g/kg diet but decreased with further increasing Schizochytrium levels.
3.4. The Expression of Genes Related to Growth Performance, Energy Metabolism, and Immune Response
3.4.1. Expression of the Genes Involved in GH-IGF-1 Axes
The expression of GH, IGF-1, IGF-1R, and IGF-2 increased obviously (p < 0.05) as Schizochytrium levels raised to 15.00 g/kg diet but decreased with further increasing Schizochytrium levels (Figure 6A).
3.4.2. Expression of the AMPK Gene
The expression of AMPK in the liver and intestine increased obviously (p < 0.05) as the Schizochytrium levels increased to 15.00 g/kg diet but decreased with further increasing Schizochytrium levels (Figure 6B,C).
3.4.3. Expression of the Genes Involved in Glycolipid Metabolism
The expression of PK, GK, G6PC2, PYGL, SREBP1, ACC1, FAS, PPARα, CPT1α, and ACAA2 in the liver increased obviously (p < 0.05) as the Schizochytrium levels increased to 15.00 g/kg diet but decreased with further increasing Schizochytrium levels (Figure 6D,E).
3.4.4. Expression of the Genes Involved in Intestinal Barrier Function
The expression of TJP1, TJP2, CLDN1, OCLN, RPL17, and LYZ1 in the intestine increased obviously (p < 0.05) as Schizochytrium levels raised to 15.00 g/kg diet but decreased with further increasing Schizochytrium levels (Figure 6F).
3.4.5. Expression of the Genes Involved in Inflammation
The expression of NF-KB, TLR4, TNF-α, NLRP3, IL-1β, IL-8, and IL-17 in the intestine declined obviously (p < 0.05) as Schizochytrium levels increased to 15.00 g/kg diet but increased with further increasing Schizochytrium levels. The expression of IL-10 in intestine rose obviously (p < 0.05) as the Schizochytrium levels increased to 15.00 g/kg diet but decreased with further increasing Schizochytrium levels (Figure 6G).
4. Discussion
In the present study, the highest values of final weight, WGR, SGR, FI, PPR, and MR were observed in the S4 group. Additionally, the higher contents of whole-body crude protein and lipids were also observed in S4 group. The results indicated that 15.00 g/kg Schizochytrium supplementation could effectively improve the growth performance, metamorphosis, feed utilization, and protein and lipid contents of tadpoles. A key contributing factor appears to be the significantly increased enzyme activities of digestion (e.g., AMS, LPS, PES) and brush border membrane (e.g., Na^+^, K^+^-ATPase, AKP, γ-GT, and CK) in the intestines of the S4 group. These enzymes are crucial for the digestion and absorption of carbohydrates, lipids, and proteins [28,29,30,31].The increased enzyme activities could improve the digestive and absorptive capacity of the intestine, thereby benefitting growth and nutrition utilization and subsequent metamorphosis of tadpoles [32,33]. Moreover, the improvement of intestinal morphology by Schizochytrium supplementation should not be neglected. Previous studies displayed that the increase in intestinal microvilli length and mucosal epithelial cell height can improve the digestive and absorptive capacity of the intestine, thereby directly affecting nutrient utilization [22,34,35].
Additionally, intestinal microbial diversity and abundance of bullfrog tadpoles were also significantly modulated by Schizochytrium levels. The supplementation of Schizochytrium exhibited higher values of ASV, Ace, and Chao1 compared with the S0 group, and the highest levels were observed in the S4 group. The findings indicated that 15.00 g/kg Schizochytrium supplementation could increase the intestinal microbial diversity of bullfrogs. Previous studies have shown that the increased microbial diversity is beneficial to the enhancement of immune function in the host [36,37,38]. These findings were further supported by the results of intestinal microbial abundance, where the supplementation of Schizochytrium increased the levels of the phyla Fusobacteriota, Firmicutes, Verrucomicrobiotam and Bacteroidota, and the S4 diet-fed tadpoles had the highest abundances of Firmicutes. A previous study has confirmed that Fusobacteriota can improve the integrity of the intestinal barrier, thus enhancing the host’s immune function [39]. Firmicutes and Bacteroidota are both involved in converting complex carbohydrates into beneficial metabolites such as short chain fatty acids (SCFAs), thereby providing energy for host resistance to pathogens [40]. Verrucomicrobiota members can produce antibiotic like substances, which are beneficial for inhibiting the growth and colonization of pathogenic bacteria in the host’s intestine [41]. Meanwhile, Firmicutes was also regarded as a sensitive indicator in the change in host weight, since the increased Firmicutes abundance is often found in hosts who have gained weight [42,43,44]. This result corresponds to the fact that S4-fed bullfrog tadpoles have relatively high growth performance. In addition, the results on the genus and species levels also showed that supplementation of Schizochytrium could increase the abundance of beneficial microbiota, such as Akkermansia glycanispila, Clostridium butyricum, and Cetobacterium somerae. Accumulating evidence suggested that all of them can improve intestinal barrier function by degrading mucin to produce SCFAs, thus leading to the enhancement of the host’s immune function [45,46,47,48].
The GH-IGF-1 axis plays an important role in the regulation of growth and feed utilization of aquatic animals. In this study, the highest expression of GH, IGF-1, IGF-1R, and IGF-2 were observed in the S4 group, which is in line with the results of growth performance. This may be attributed to the fact that DHA-abundant Schizochytrium can stimulate the generation of GH, thus leading to an increase in GH expression [49]. Then, the increase in GH induces the release of IGF-1 [50], thereby improving the growth performance of tadpoles.
At the metabolic level, the expression of genes related to energy metabolism (AMPK, PK, GK, G6PC2, PYGL, SREBP1, ACC1, FAS, PPARα, CPT1α, and ACAA2) in the liver upregulated obviously as the Schizochytrium levels increased to 15.00 g/kg diet. The results indicated that the S4 diets significantly enhance glycolysis, gluconeogenesis, glycogenolysis, lipogenesis, and lipolysis of tadpoles. According to a previous study, AMP-activated protein kinase (AMPK) is a highly conserved sensor of cellular energy status that can regulate the energy metabolism of organisms [51], and DHA-rich food can activate the AMPK [52]. The activation of AMPK can participate in a series of physiological consequences concerning glycolipid metabolism, such as (1) the enhancement of glycolysis and glycogenolysis by upregulating GK and PYGL activities, respectively [53,54]; and (2) the increase in lipolysis by activating PPARα and PPARα-downstream target genes (CPT1α and ACAA2) [55]. In addition, DHA has been demonstrated to upregulate expression of lipogenesis-related genes, thereby resulting in an increase in lipid contents [56]. For gluconeogenesis, this may be attributed to the enhanced glycolysis that could result in an increase in pyruvate content, which in turn stimulates gluconeogenesis characterized by the upregulation of G6PC2 expression [57].
Intestinal health partly depends on intestinal barrier function which is associated with the activities of tight junction proteins, such as claudins and occlusions [58]. In this study, the mRNA levels of AMPK and tight junction proteins (e.g., TJP1, TJP2, CLDN1, OCLN, RPL17, and LYZ1) in the intestine increased significantly as the Schizochytrium levels increased to 15.00 g/kg diet, indicating that the S4 diet significantly enhanced the intestinal barrier function of tadpoles. The results may be due to the fact that the activation of AMPK could promote tight junction-related protein synthesis, thereby leading to the enhancement of intestinal barrier function characterized by the upregulation of tight junction proteins expression [59,60]. According to previous research, the enhanced intestinal barrier function can enhance the immunity of aquatic animals [61]. Consistent with an enhanced barrier and AMPK’s role in modulating redox and inflammatory states, the S4 group exhibited significantly elevated activities of antioxidant enzymes (SOD, CAT) and a marked down-regulation of pro-inflammatory genes (NF-KB, TLR4, TNF-α, NLRP3, IL-1β, IL-8, IL-17). AMPK can activate the Nrf2-mediated antioxidant pathway and inhibit NF-kB signaling by deacetylating the p65 subunit [62,63,64], thereby collectively reducing intestinal inflammation and oxidative stress.
5. Conclusions
In conclusion, dietary supplementation of Schizochytrium significantly enhanced bullfrog tadpole growth and metamorphosis by simultaneously improving digestive function, intestinal barrier integrity, and intermediate metabolism, while attenuating intestinal inflammation. Optimal Schizochytrium levels for bullfrog tadpoles were estimated to be 13.27 g/kg diet. This study provided a practical nutritional strategy for sustainable bullfrog aquaculture. A limitation of this work was that the precise causal relationships between the reshaped gut microbiota and the observed metabolic benefits warrant further investigation. Future studies should focus on validating these findings under commercial farming conditions and elucidating the underlying molecular mechanisms in greater detail.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mansano C.F.M. Stéfani M.V.D. Pereira M.M. Macente B.I. Non-linear growth models for bullfrog tadpoles Cienc. Agrotecnol.20123645446210.1590/S 1413-70542012000400010 · doi ↗
- 2Scott D.E. Casey E.D. Donovan M.F. Lynch T.K. Amphibian lipid levels at metamorphosis correlate to post-metamorphic terrestrial survival Oecologia 200715352153210.1007/s 00442-007-0755-617530291 · doi ↗ · pubmed ↗
- 3Stamper C. Downie J. Stevens D. Monaghan P. The effects of perceived predation risk on pre-and post-metamorphic phenotypes in the common frog J. Zool.200927720521310.1111/j.1469-7998.2008.00526.x · doi ↗
- 4Alvarez R. Real M. Significance of initial weight of post-metamorphosis froglets for growth and fattening of Rana perezi Seoane, 1885, raised in captivity Aquaculture 200625542943510.1016/j.aquaculture.2005.12.026 · doi ↗
- 5Cheng R.B. Ma R.J. Li K. Rong H. Lin X.Z. Wang Z.K. Yang S.J. Ma Y. Agrobacterium tumefaciens mediated transformation of marine microalgae Schizochytrium Microbiol. Res.201216717918610.1016/j.micres.2011.05.00321641193 · doi ↗ · pubmed ↗
- 6Shields R. Bell G. Evaluation of a DHA-rich microalga (Schizochytruim sp.) as a diet enrichment for larval Atlantic halibut (Hippoglossus hippoglossus L.)Spec. Publ. Eur. Aquac. Soc.199524188191
- 7Wang Y. Li M. Filer K. Xue Y. Ai Q. Mai K. Replacement of fish oil with a DHA-rich Schizochytrium meal on growth performance, activities of digestive enzyme and fatty acid profile of Pacific white shrimp (Litopenaeus vannamei) larvae Aquacult. Nutr.2017231113112010.1111/anu.12479 · doi ↗
- 8Xie S.W. Wei D. Tan B.P. Liu Y.J. Tian L.X. Niu J. Schizochytrium limacinum supplementation in a low fish-meal diet improved immune response and intestinal health of juvenile Penaeus monodon Front. Physiol.20201161310.3389/fphys.2020.0061332714197 PMC 7344155 · doi ↗ · pubmed ↗