The Protective Effect of Phenolic Acids on Liver Disease: A Review of Possible Mechanisms
Xinyi Ma, Suhui Xiong, Feng Xiang, Yamei Li, Yan Lin, Yuexin Liu, Limei Lin, Jingchen Xie

TL;DR
This review explores how phenolic acids may protect the liver from various diseases through mechanisms like reducing inflammation and regulating metabolism.
Contribution
The paper systematically reviews the mechanisms by which phenolic acids protect against multiple liver diseases.
Findings
Phenolic acids show protective effects against drug-induced liver injury, alcoholic liver disease, and liver fibrosis.
Their benefits include anti-inflammatory, antioxidant, and lipid-regulating properties.
Phenolic acids also modulate gut microbiota and inhibit cell apoptosis in liver diseases.
Abstract
(1) Background: This article reviews the biological characteristics of phenolic acid compounds, focusing on their mechanisms of action in various liver diseases. (2) Methods: The review adheres to the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines. We utilized PubMed and Web of Science databases to search for relevant studies on the use of phenolic acids in liver diseases from 2015 to 2025. (3) Results: Phenolic acids can improve different types of liver diseases, including drug-induced liver injury (DILI), alcoholic liver disease (ALD), metabolic dysfunction-associated steatotic liver disease, liver fibrosis, and liver cancer. Their beneficial effects are attributed to mechanisms such as anti-inflammatory properties, antioxidant activity, regulation of lipid metabolism, inhibition of cell apoptosis, and modulation of gut microbiota. (4)…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
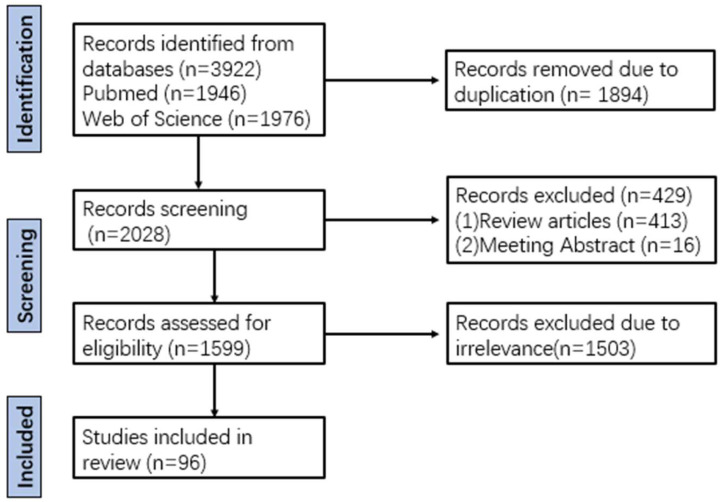 Figure 1
Figure 1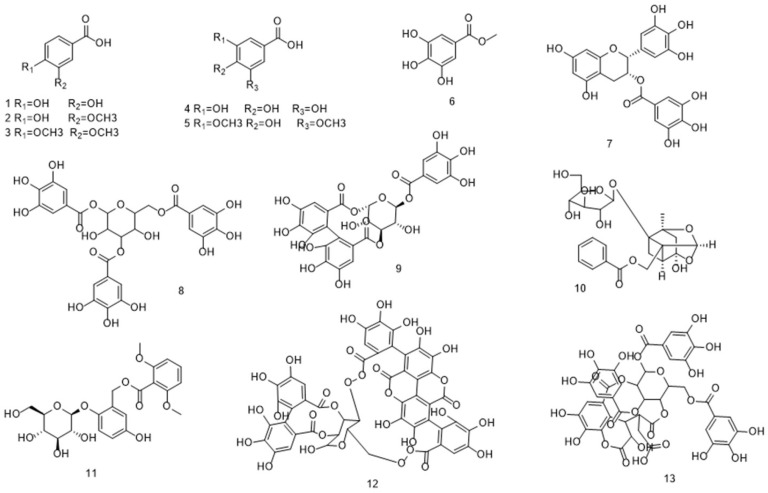 Figure 2
Figure 2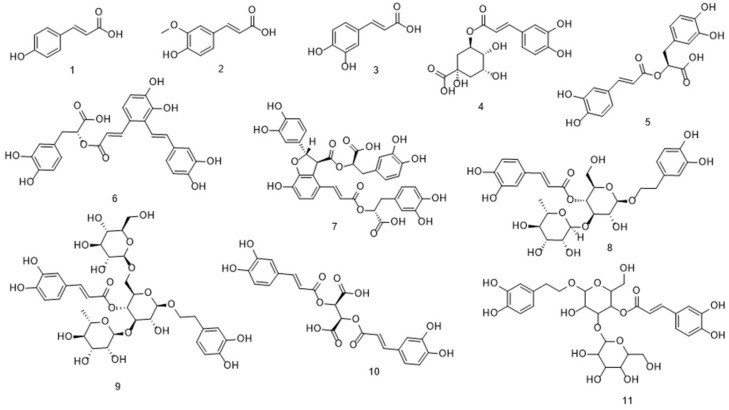 Figure 3
Figure 3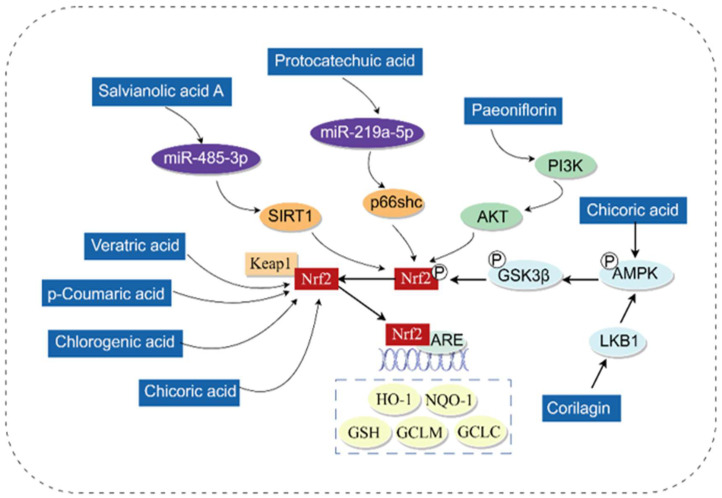 Figure 4
Figure 4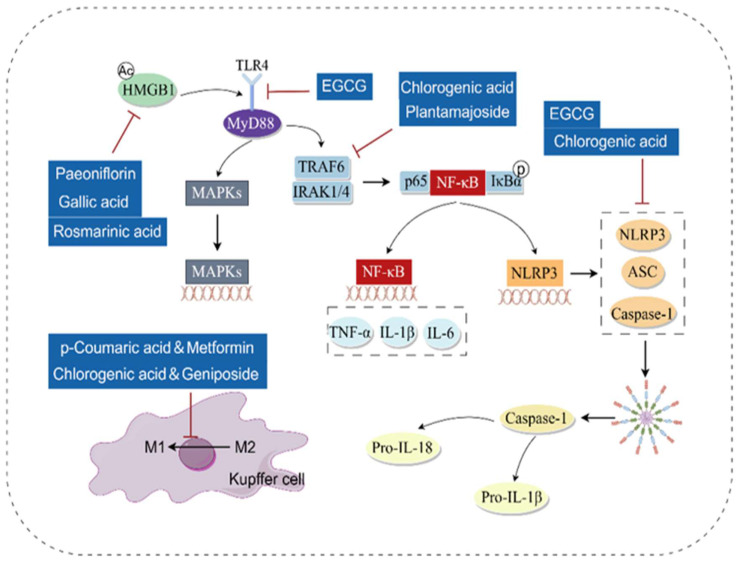 Figure 5
Figure 5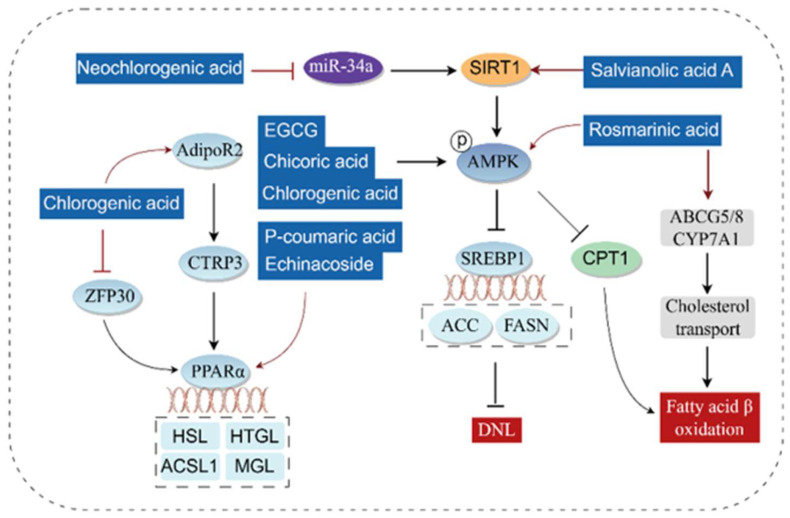 Figure 6
Figure 6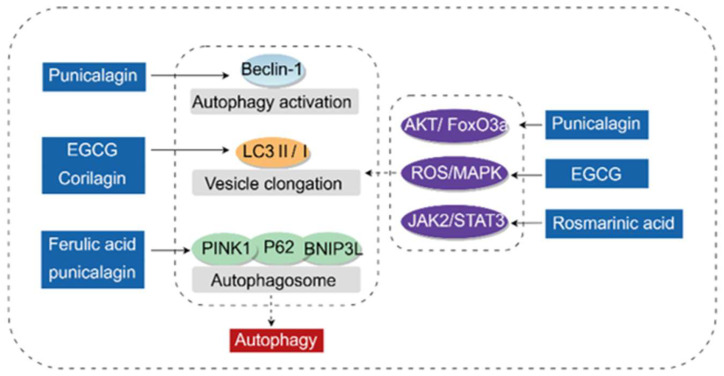 Figure 7
Figure 7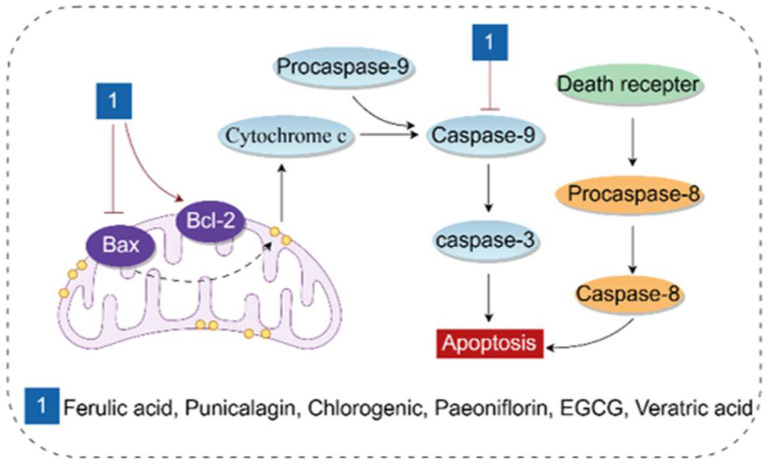 Figure 8
Figure 8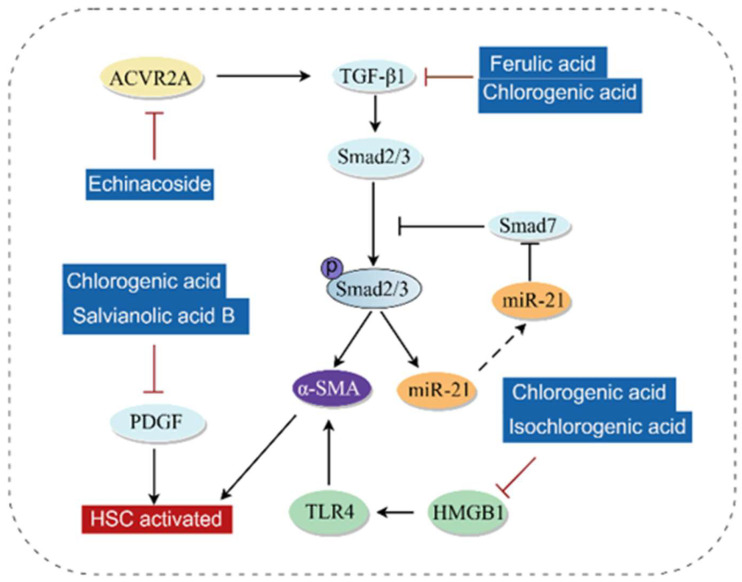 Figure 9
Figure 9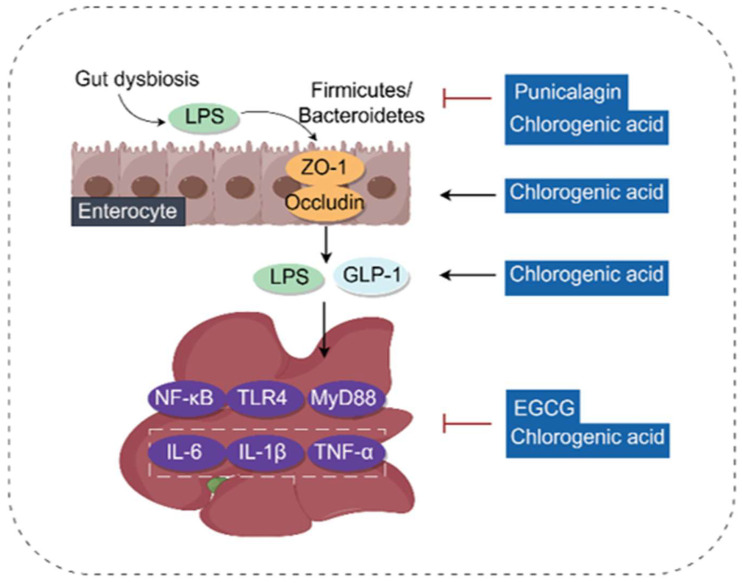 Figure 10
Figure 10- —National Natural Science Foundation of China
- —Project of Hunan University of Chinese Medicine
- —Key Research Foundation of Education Bureau of Hunan Province
- —Key Discipline Project on Chinese Pharmacology of Hunan University of Chinese Medicine
- —Hunan Provincial Natural Science Foundation
- —Scientific Research Fund of Hunan Provincial Education Department
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLiver Disease Diagnosis and Treatment · Biochemical Acid Research Studies · Drug-Induced Hepatotoxicity and Protection
1. Introduction
The liver is the largest and most important organ in the human body, with basic functions such as metabolism, secretion, and detoxification. Liver diseases cause more than 2 million deaths annually, accounting for 4% [1] of all deaths worldwide and have become a major cause of death. With the improvement in living standards, the prevalence of metabolic liver diseases, including nonalcoholic fatty liver disease (NAFLD) and ALD, is constantly rising, ultimately leading to an increase in cases of end-stage liver disease (liver fibrosis and liver cancer) [2]. Currently, no study has specifically analyzed the real incidence of DILI worldwide, but in follow-up studies, the proportion of DILI patients caused by herbal medicines and dietary supplements has been increasing [3].
Common hepatoprotective drugs in clinical practice include silibinin, ursodeoxycholic acid (UDCA), glycyrrhetinic acid preparations, bicyclol, and others. Silymarin is a substance extracted from Silybum marianum and is a widely used hepatoprotective drug in clinical settings. It exerts a good protective effect on various diseases such as NAFLD, ALD, and hepatitis [4]. However, caution is still required not only for the use of silymarin but also for other hepatoprotective drugs like glycyrrhizic acid preparations and bicyclol. This is because there are still very few clinical studies on their appropriate dosages and actual efficacy in different types of diseases. UDCA is the only drug approved by the U.S. Food and Drug Administration (FDA) for the treatment of primary biliary cholangitis. It was originally identified as a naturally active product from the bile of polar bears, and its current acquisition mainly relies on chemical or biosynthetic approaches for preparation. Nevertheless, the preparation process of UDCA still poses a major challenge in terms of cost and environmental protection [5]. Despite significant progress made in the field of drug research and development, there is no fully effective drug that can enhance liver function, provide whole-organ protection, or promote hepatocyte regeneration. Therefore, research on safer, more effective, and more economical hepatoprotective drugs is of crucial importance.
Phenolic acids are a class of bioactive substances widely present in nature, belonging to aromatic secondary metabolites. According to the differences in carbon skeleton and the position and quantity of hydroxyl groups on the aromatic ring, they can be classified into hydroxybenzoic acids (protocatechuic acid, vanillic acid, gallic acid, 4-hydroxybenzoic acid) and hydroxycinnamic acids (chlorogenic acid, ferulic acid, caffeic acid, rosmarinic acid) [6]. And a large number of studies have reported the hepatoprotective effects of phenolic acids [7]. Compared with existing clinical hepatoprotective drugs, phenolic acids, as naturally derived compounds, have potential advantages such as easy accessibility, low cost, and multiple pharmacological activities. However, they also have current limitations including low bioavailability and a lack of clinical trials.
However, most current studies focus on reports of individual phenolic acid monomers in single diseases, and there is a lack of systematic sorting and summary of the mechanisms underlying the hepatoprotective effects of phenolic acids, which limits their application. Therefore, this review followed the PRISMA guidelines. Several literature databases were searched, including PubMed and Web of Science, for papers published from 2015 to 2025. Keywords included “drug-induced liver injury”, “alcoholic liver disease”, “metabolic (dysfunction) associated fatty liver disease”, “liver fibrosis”, “liver cancer”, “phenolic acid” and “polyphenols”. Duplicate articles were excluded, and review articles and conference abstracts were excluded, along with unrelated studies, resulting in a final count of 96 eligible papers (Figure 1). Their pharmacological activities and related mechanisms were sorted out, aiming to provide a valuable theoretical basis for the research and development of phenolic acids in liver diseases and reduce the socioeconomic burden of liver diseases.
2. Biological Characteristics of Phenolic Acids
2.1. Chemical Structure of Phenolic Acids
Through literature review, it was found that there are 24 types of phenolic acid components with relevant hepatoprotective effects, mainly consisting of hydroxybenzoic acids and hydroxycinnamic acids. Among them, there are 13 types of hydroxybenzoic acids and 11 types of hydroxycinnamic acids.
Phenolic acids with a hydroxybenzoic acid skeleton are collectively referred to as hydroxybenzoic acids, which can be classified into simple hydroxybenzoic acids, condensed hydroxybenzoic acids, hydroxybenzoic acid esters, and hydroxybenzoic acid glycosides based on their structures. It has been reported that among these phenolic acids with hepatoprotective effects, simple hydroxybenzoic acids are the main type, including protocatechuic acid, vanillic acid, gallic acid, and veratric acid. Meanwhile, hydroxybenzoic acid esters, such as methyl gallate and epigallocatechin-3-gallate (EGCG), have been the subject of most studies. Their structures are shown in Figure 2.
Hydroxycinnamic acids can be classified by structure into simple hydroxycinnamic acids, hydrogenated hydroxycinnamic acids, condensed hydroxycinnamic acids, hydroxycinnamic acid esters, hydroxycinnamic acid glycosides and hydroxycinnamate salts, among which simple hydroxycinnamic acids and condensed hydroxycinnamic acids are the most common types. It has been reported that among these phenolic acids with hepatoprotective effects, the most common simple hydroxycinnamic acids are p-coumaric acid, ferulic acid, and caffeic acid. Next are condensed hydroxycinnamic acids condensed with caffeic acid as the parent nucleus, including chlorogenic acid, rosmarinic acid, and chicoric acid; in addition, there are hydroxycinnamic acid esters and hydroxycinnamic acid glycosides such as salvianic acid A and salvianic acid B. Their structures are shown in Figure 3.
2.2. The Relationship Between Chemical Structure and Biological Activity
Studies on the relationship between compound structure and biological activity are of great significance for improving the targeting and pharmacokinetic properties of drugs, as well as for the research and development of new drugs. Phenolic acids are a class of aromatic compounds containing both phenolic hydroxyl groups and carboxylic acid groups. The number and position of hydroxyl groups are closely related to the activity of phenolic acids. A structure-activity relationship (SAR) study on phenolic acid compounds using the crocin bleaching assay revealed that the position of the carboxyl group relative to the hydroxyl substituent is crucial. Among hydroxybenzoic acids, gallic acid exhibits the strongest activity, which is 1.6 times and 3.4 times that of protocatechuic acid and syringic acid, respectively [8]. The SAR value of gallic acid, which has an additional hydroxyl group at the ortho position of the phenolic hydroxyl group, is more than twice that of protocatechuic acid (3.40 vs. 1.50). This indicates that the pyrogallol moiety in the molecular structure of phenolic compounds has better antioxidant activity than the catechol moiety [9,10]. This is because the ortho hydroxyl groups (electron donors) on the aromatic ring reduce the dissociation enthalpy of hydroxyl groups, thereby enhancing free radical scavenging activity [11].
In addition, special phenolic hydroxyl structures (such as ortho-dihydroxyl groups) and unsaturated bonds in side chains can also affect the activity of phenolic acids. The presence of free phenolic hydroxyl groups in caffeic acid reduces the dissociation enthalpy of hydroxyl groups, increases the hydrogen atom transfer rate of peroxyl radicals and the number of peroxyl radicals on the benzene ring, thereby enhancing the activity of phenolic acids. Furthermore, the presence of double bonds in the carbon chain increases the stability of phenolic hydroxyl radicals. These chemical factors related to caffeic acid are closely associated with its ability to scavenge free radicals and inhibit the production of reactive oxygen species (ROS) [12,13,14]. Meanwhile, some substituents can also enhance the activity of phenolic acids. Rosmarinic acid is an ester of caffeic acid and 3,4-dihydroxyphenyllactic acid. Previous studies have shown that rosmarinic acid has relatively weak antibacterial activity. However, among a series of successfully synthesized rosmarinic acid derivatives, RA-N8 (6-trifluoromethoxy-benzothiazol-2-yl) is the most effective antibacterial agent (against Staphylococcus aureus and Escherichia coli) [15]. In addition, rosmarinic acid can also form derivatives by combining with metal ions (sodium, silver), amine or imidazole, thereby exerting an anti-glioblastoma effect [16].
2.3. Pharmacokinetic Properties
Although phenolic acids are widely present in plant-based foods, their bioavailability is generally low. After ingestion, caffeic acid undergoes esterification; the esterified part of caffeic acid in the colon is cleaved by microbial esterases, and approximately 95% of it is then absorbed by the intestinal mucosa in free form. After absorption, enzymatic reactions (methylation, sulfation, and glucuronidation) occur to increase hydrophilicity, reduce toxic effects, and promote elimination [17]. Rosmarinic acid is easily absorbed from the gastrointestinal tract and through the skin and is distributed in various tissues. However, a small portion of rosmarinic acid is degraded into various components, such as glucuronic acid or sulfated conjugates of caffeic acid, ferulic acid, and trace amounts of coumaric acid, which are then rapidly excreted in urine [18]. The gastrointestinal tract has difficulty absorbing chlorogenic acid because neither the intestinal mucosa nor the intestinal flora possesses esterases that can cleave chlorogenic acid, due to the presence of the quinic acid moiety in the chlorogenic acid structure [19]. EGCG also has poor bioavailability. It is metabolized by the intestinal microbiota before absorption: it can be hydrolyzed into epigallocatechin (EGC) and gallic acid by rat intestinal bacteria and bacterial strains, followed by a series of transformations and degradation processes before being excreted from the body [20]. The main site of ferulic acid absorption in the human body is the colon; its absorption rate is fast, but the extent is very low. Intestinal microbial esterases hydrolyze esterified ferulic acid in food matrices to produce free ferulic acid, which is mainly excreted as feruloyl glucuronide [21]. In summary, the absorption and metabolism of phenolic acids are highly dependent on their structural characteristics, as well as the activity of intestinal esterases and intestinal flora. Caffeic acid and rosmarinic acid are well absorbed in the intestine. In contrast, chlorogenic acid, EGCG, and ferulic acid have low bioavailability. The lack of intestinal esterases and rapid pre-absorption metabolism by the intestinal flora reduces their absorption.
3. Molecular Mechanisms of Phenolic Acids in Modulating Liver Diseases
3.1. Antioxidant
Oxidative stress is caused by an imbalance between the production of ROS and antioxidant defense [22]. A substantial body of evidence suggests that oxidative stress plays a critical role in the pathophysiology of various liver diseases [23]. When exposed to stressors, the body’s antioxidant defense system is activated, in which nuclear factor erythroid 2-related factor 2 (Nrf2) acts as the primary regulator [24,25]. The dissociation of Kelch-like ECH-associated protein 1 (Keap1) from Nrf2 enables Nrf2 to translocate into the nucleus, where it binds to small Musculoaponeurotic fibrosarcoma oncogene homolog (MAF) proteins that associate with promoters containing Antioxidant response elements (ARE) [26,27]. This binding regulates the expression of a series of antioxidant factors, such as Heme oxygenase 1 (HO-1), NAD(P)H: Quinone oxidoreductase 1 (NQO1), Glutamate cysteine ligase catalytic (GCLC), and Glutamate cysteine ligase modifier (GCLM). Phenolic acids exert their antioxidant effects mainly by activating the Nrf2 pathway.
Research has shown that various phenolic acids can alleviate liver damage by activating Keap1/Nrf2 to exert antioxidant effects. For example, p-Coumaric acid, Caffeic acid, Rosmarinic acid, Veratric acid, and Chlorogenic acid can activate Nrf2 and upregulate the expression of downstream antioxidant proteins (HO-1, NQO1, GCLC, and GCLM), thereby playing a hepatoprotective role [28,29,30,31]. In the exploration of the antioxidant mechanism of phenolic acids, it has been found that phenolic acids can also regulate Nrf2 by acting on upstream pathways, including the AMP-activated protein kinase (AMPK)/Glycogen synthase kinase 3β (GSK3β) and Phosphoinositide 3-kinase (PI3K)/Protein kinase B (AKT) signaling pathways. Both chicoric acid and corilagin can increase the levels of Phospho-AMP-activated protein kinase α (P-AMPKα) and Phospho-Glycogen synthase kinase 3β (P-GSK3β), thereby activating Nrf2 and alleviating Lipopolysaccharides (LPS)/D-Galactosamine (GalN)-induced Acute liver failure (ALF) and Acetaminophen(APAP)-induced hepatotoxicity [32,33]. Paeoniflorin activates Nrf2 through the PI3K/AKT-dependent pathway, improving Alpha-Naphthylisothiocyanate (ANIT)-induced cholestatic liver injury in rats [34]. In addition, phenolic acids can exert antioxidant effects by targeting MicroRNA (miRNA). For instance, protocatechuic acid can reduce the formation of ROS through the MicroRNA-219a-5p (miR-219a-5p)/p66shc signaling pathway, thereby alleviating ALD [35]. Salvianolic acid A regulates the MicroRNA-485-3p (miR-485-3p)/Sirtuin 1 (SIRT1) pathway to mitigate APAP-induced oxidative stress [36].
Phenolic acids exhibit protective effects against liver injury induced by various drugs, particularly DILI caused by drugs such as APAP, Isoniazid, Rifampicin, Tetrachloromethane (CCl_4_), and Methotrexate (MTX). Impaired liver metabolic capacity leads to sustained oxidative stress in the liver; caffeic acid, gallic acid, and punicalagin can achieve hepatoprotective effects by exerting antioxidant activity [37,38,39,40]. DILI includes acute and chronic injury. For anti-tuberculosis drugs like isoniazid and rifampicin—which have a marked tendency to induce liver injury and require long-term administration—hepatoprotective drugs are generally co-administered in clinical practice to prevent liver injury. Currently, most animal experiments focus on preventive administration, so the therapeutic effect on unknown acute liver injury remains unclear. Drugs such as CCl_4_ and MTX are frequently used in animal experiments but lack sufficient clinical relevance.
The molecular mechanism by which phenolic acids exert antioxidant activity through the Nrf2 pathway is shown in Figure 4.
3.2. Anti-Inflammatory
When the liver is damaged by various etiologies (such as alcohol, hepatitis virus, lipid accumulation, cholestasis, etc.), the inflammatory response is a crucial link in the liver’s initiation of self-repair mechanisms. Nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) is a key transcriptional regulator of the inflammatory response. Pathogen-associated molecular patterns (PAMPs) such as LPS, as well as Damage-associated molecular patterns (DAMPs) including High mobility group box 1 (HMGB1), can all activate NF-κB by acting on Toll-like receptors (TLRs), especially Toll-like receptor 4 (TLR4), leading to liver inflammation [41]. Studies have found that phenolic acids alleviate liver inflammation and mitigate liver diseases by inhibiting the NF-κB pathway.
(1) A variety of phenolic acids can reduce liver inflammation by inhibiting the TLR4/NF-κB pathway. For example, Chlorogenic acid and EGCG alleviate liver inflammation by directly inhibiting TLR4, thereby slowing the progression of nonalcoholic steatohepatitis (NASH) and liver injury caused by Pseudomonas aeruginosa infection [42,43]. Plantamajoside and chlorogenic acid inhibit the expression of Interleukin-1 Receptor-Associated Kinase 1 (IRAK1) and TNF Receptor-Associated Factor 6 (TRAF6), preventing NF-κB from translocating into the nucleus. This reduces the mRNA transcription of inflammatory factors and alleviates liver inflammation [44,45]. Protocatechuic acid and punicalagin can decrease the levels of pro-inflammatory cytokines such as Interleukin-6 (IL-6) and Tumor necrosis factor-α (TNF-α), thereby improving liver injury [40,46].
(2) In addition, as a DAMP, HMGB1 can be passively released from necrotic cells or actively secreted from immunocompetent cells during inflammation [47]. It interacts with TLR4 to further activate NF-κB, exacerbating the inflammatory response. Gallic acid, rosmarinic acid, paeoniflorin, and chlorogenic acid all exert anti-inflammatory effects by inhibiting the HMGB1/NF-κB/TLR4 pathway [38,48,49,50].
(3) On the other hand, the activation of NF-κB also induces the transcription of NOD-, LRR- and pyrin domain-containing protein 3 (NLRP3). This enables the NLRP3 inflammasome to recruit the adaptor protein Apoptosis-associated Speck-like protein containing a CARD (ASC) and Caspase 1, inducing the assembly of the NLRP3 inflammasome. Consequently, this activates Caspase 1 through autocatalysis and promotes the maturation and secretion of pro-inflammatory cytokines [51]. Both EGCG and chlorogenic acid can inhibit the activation of the NLRP3 inflammasome by downregulating the expression of NLRP3, ASC, and Interleukin-1β (IL-1β), thereby exerting hepatoprotective effects [31,52].
Meanwhile, phenolic acids can also inhibit Macrophage type 2 (M2) macrophage polarization, reduce the inflammatory infiltration of hepatic macrophages, and alleviate the inflammatory response. Combined treatment with metformin and p-coumaric acid, as well as combined administration of geniposide and chlorogenic acid, can improve liver inflammation by promoting the transition of macrophages from the Macrophage type 1 (M1) phenotype to the M2 phenotype, thereby inhibiting the inflammatory infiltration of hepatic macrophages [53,54]. Notably, opposite results have been observed in liver fibrosis models. Corilagin mediates the repolarization of M2 macrophage to M1 macrophage by inhibiting the expression of Indoleamine 2,3-Dioxygenase 1 (IDO1), thereby improving CCl_4_-induced liver fibrosis [55]. This is because chronic diseases such as chronic viral hepatitis, ALD, and NASH often lead to an imbalance between M1 and M2 responses. Activated M2 macrophages promote the activation of hepatic stellate cells (HSCs), resulting in liver fibrosis. This reflects the complexity of the inflammatory microenvironment during liver injury and repair: in the early stage of injury, the focus may be on inhibiting the pro-inflammatory M1 type, while in the repair stage, promoting the anti-inflammatory M2 type is more critical. Most existing studies fail to clearly define the temporal and spatial context of their interventions. Future research should not be limited to achieving phenotypic changes but must deeply reveal the specific immune microenvironment and molecules that drive such changes.
The molecular mechanism by which phenolic acids exert anti-inflammatory effects through the NF-κB pathway is shown in Figure 5.
3.3. Regulating Lipid Metabolism
An increase in fatty acids (FAs) delivered by adipose tissue, accompanied by an imbalance between lipid degradation and De novo lipid (DNL) synthesis, leads to excessive lipid accumulation in the liver and triggers the development of various liver diseases [56,57]. Phenolic acids can promote FAs β-oxidation and inhibit DNL by regulating pathways such as the (AMP)-activated protein kinase (AMPK)/Carnitine Palmitoyltransferase 1 (CPT1) pathway, AMPK/Sterol Regulatory Element-Binding Protein 1 (SREBP-1) pathway, and Peroxisome Proliferator-Activated Receptors (PPARs) pathway.
After FAs in the liver bind non-covalently to Fatty Acid Binding Protein-1 (FABP-1), CPT1 converts them into acylcarnitines, which are then shuttled into mitochondria. There, these acylcarnitines undergo β-oxidation to produce energy [58]. As a cellular “energy sensor”, the activation of AMPK phosphorylates and inhibits the activity of Acetyl CoA Carboxylase (ACC), leading to a decrease in malonyl-CoA levels. This, in turn, relieves the inhibition of CPT1 activity and promotes FAs entry into mitochondria for β-oxidation [59]. Combined treatment with metformin and p-coumaric acid, as well as rosmarinic acid, can promote FAs β-oxidation by activating AMPK-mediated CPT1 [53,60]. At the cellular level, studies have also shown that arctigenin chlorogenic acid from burdock roots enhances FAs β-oxidation through the AMPK/ACC/CPT1 pathway, thereby alleviating oleic acid-induced steatosis in Human Hepatoma G2 (HepG2) cells [61]. SIRT1 is a class III Nicotinamide Adenine Dinucleotide (NAD+)-dependent histone/protein deacetylase. A decrease in its activity may reduce the deacetylation of LKB1 and inhibit this kinase [62], which in turn regulates the activation of AMPK [63] and ultimately affects FAs metabolism. Salvianolic acid A upregulates the protein levels of phosphorylated AMPK and SIRT1 in a dose-dependent manner, improving hepatic lipotoxicity in mice with fatty liver [64]. Neochlorogenic acid alleviates lipid accumulation by downregulating MicroRNA-34a (miR-34a), thereby activating the SIRT1/AMPK pathway to improve oleic acid-induced intracellular lipid accumulation [65].
Phenolic acids can act on the AMPK/SREBP-1 pathway to inhibit lipogenesis and lipid accumulation. SREBP-1 is a key transcription factor for lipogenic genes [66]. AMPK phosphorylates SREBP-1 and inhibits its transcriptional activity, reducing the protein expression of ACC and Fatty Acid Synthase (FASN) [67,68], thereby suppressing DNL. Combined use of chicoric acid and fish oil can reduce palmitic acid-induced cellular lipid accumulation by regulating the AMPK-mediated SREBP-1/FASN pathway [69]. EGCG, chicoric acid, chlorogenic acid and combined use with metformin can all activate the p-AMPK/AMPK, downregulate the expression of Sterol Regulatory Element-Binding Protein 1c (SREBP1C) and its downstream lipogenesis-related targets (ACC and FASN), thereby alleviating NAFLD [70,71,72].
In addition, phenolic acids can act on the PPARs pathway to promote FAs β-oxidation. PPARs are transcription factors of the nuclear receptor family that regulate the gene transcription of key enzymes involved in FAs β-oxidation [73]. Among them, PPARα is mainly expressed in the liver. Studies have shown that chlorogenic acid can upregulate the hepatic expression of Adiponectin Receptor 2 (AdipoR2), increase the level of C1q/TNF-Related Protein 3 (CTRP3), and upregulate hepatic PPARα expression, thereby exerting hypoglycemic and lipid-regulating effects [74]. p-Coumaric acid activates PPARα to upregulate Hormone-Sensitive Lipase (HSL), Hepatic Triglyceride Lipase (HTGL), Monoacylglycerol Lipase (MGL), and Acyl-CoA Synthase Long-Chain 1 (ACSL1), thereby reducing serum and hepatic lipids and inhibiting the fusion and growth of lipid droplets [75]. Echinacoside can alleviate hepatic steatosis by upregulating PPARα [76]. Methyl ferulate can also promote the expression of PPARα and CPT1 by upregulating SIRT1, significantly inhibiting ethanol-induced hepatic steatosis [77]. ZFP30 is a member of the Krüppel-associated box zinc finger protein (KRAB-ZFP) family; recent studies have shown that chlorogenic acid inhibits the upregulation of ZFP30 in NAFLD and promotes the expression of PPARα, thereby enhancing FAs β-oxidation and alleviating hepatic steatosis [78]. Peroxisome Proliferator-Activated Receptor γ (PPARγ) is mainly expressed in adipose tissue. Peroxisome Proliferator-Activated Receptor Gamma Coactivator-1α (PGC-1α) is a transcriptional coactivator of PPARγ and is involved in the transcriptional regulation of PPARγ. Rosmarinic acid significantly alleviates NAFLD by repairing mitochondrial damage and regulating the PPARγ/PGC-1α signaling pathway [79].
Beyond the aforementioned common mechanisms, phenolic acids can also regulate FAs β-metabolism through pathways such as the Epidermal Growth Factor Receptor (EGFR)-AKT-mechanistic Target of Rapamycin (mTOR) pathway and Forkhead Box O1 (FoxO1) pathway. Salvianolic acid A inhibits the nuclear translocation of Carbohydrate Response Element-Binding Protein (ChREBP) by suppressing Thioredoxin-Interacting Protein (TXNIP), thereby improving high-fat diet (HFD)-induced hepatic steatosis [80]. Tannic acid and vitamin E-loaded-poly D, L-lactide-co-glycolic acid (PLGA) nanoparticles prevent hepatic steatosis and improve liver injury in a chronic alcoholic liver injury model by inhibiting the EGFR-AKT-mTOR pathway [81]. FoxO1 is a member of the FoxO family and plays an important role in hepatic steatosis [77]. Methyl ferulate downregulates the phosphorylated expression of p38 Mitogen-Activated Protein Kinase (p38 MAPK), c-Jun N-Terminal Kinase (JNK), Extracellular Signal-Regulated Kinase (ERK), and FoxO1 in L-02 cells, inhibiting ethanol-induced hepatic steatosis [77]. Rosmarinic acid exerts lipid-lowering effects by increasing the expression of ATP Binding Cassette G5 (ABCG5), ATP Binding Cassette G8 (ABCG8), and Cholesterol 7α-Hydroxylase A1 (CYP7A1) [60].
The molecular mechanism by which phenolic acids regulate lipid metabolism is shown in Figure 6.
3.4. Regulating Autophagy
Autophagy is an evolutionarily conserved cellular degradation process [82] which plays a key role in maintaining cellular homeostasis by degrading protein aggregates, pathogens, lipids, and aged/damaged subcellular organelles (e.g., damaged mitochondria) [83]. Dysfunction of hepatic autophagy may lead to various liver diseases, including NAFLD, DILI, cholestasis, and hepatocellular carcinoma (HCC). Autophagy can be classified into microautophagy, macroautophagy, and chaperone-mediated autophagy. Phenolic acids primarily exert hepatoprotective effects by regulating the macroautophagy pathway. This process mainly involves the following key steps: (a) formation or nucleation of phagophores; (b) conjugation of Autophagy-related gene 5—Autophagy-related gene 12 (Atg5-Atg12), interaction with Autophagy-related gene 16-like (Atg16L), and polymerization of phagophores; (c) processing of Microtubule-associated protein 1A/1B-light chain 3 (LC3) and its insertion into the membrane of elongating phagophores; (d) fusion of autophagosomes with lysosomes, followed by proteolytic degradation of engulfed molecules via lysosomal proteases.
The hepatoprotective effect of phenolic acids via autophagy is mainly achieved by promoting LC3 processing and regulating autophagy-related proteins such as Beclin1, PTEN-induced kinase 1 (PINK1), and Parkin.
(1) Beclin1 is a key protein involved in autophagy. It participates in the vesicle nucleation process by forming the Beclin1-Vps34 complex with Vacuolar protein sorting 34 (Vps34), Vacuolar protein sorting 15 (Vps15), and Autophagy-related gene 14-like (ATG14L) [84]. Punicalagin increases hepatic Beclin1 expression and the number of hepatic autophagosomes, thereby inhibiting insulin resistance in HFD-fed mice [85].
(2) After autophagy induction, LC3 is hydrolyzed to generate LC3I. Subsequently, LC3I is activated by Autophagy-related gene 7 (Atg7) and transferred to Autophagy-related gene 3 (Atg3). Finally, LC3I conjugates with Phosphatidylethanolamine (PE) via carboxyglycine to form processed LC3II [86]. Punicalagin, EGCG, and corilagin promote the processing of LC3II, upregulate autophagy, and alleviate diabetic liver injury, as well as liver injury mediated by APAP and HFD [85,87,88].
(3) PINK1 can translocate to the outer mitochondrial membrane to recruit Parkin [89], and is then ubiquitinated by Parkin. This ubiquitinated complex is subsequently recognized by Sequestosome 1 (p62/sequestosome 1) [90,91], which bridges mitochondria targeted for autophagy to LC3 on the autophagosome surface, thereby driving the elongation of phagophore membranes. In addition to PINK1-mediated autophagy, BCL2/adenovirus E1B interacting protein 3-like (BNIP3L) can also mediate autophagosome formation. Punicalagin increases the protein expression of hepatic P62 and BNIP3, enhances the number of hepatic autophagosomes, and inhibits insulin resistance and diabetic liver injury [92]. Ferulic acid activates the autophagic pathway by increasing the levels of PINK1, Parkin, and P62 [93].
(4) Beyond directly regulating autophagy-related proteins, in-depth studies have shown that certain phenolic acid components can also regulate autophagy by acting on signaling pathways such as AKT/Forkhead box O3 (FoxO3a), ROS/MAPK, and Janus kinase 2 (JAK2)/Signal transducer and activator of transcription 3 (STAT3). Punicalagin inhibits the phosphorylation of AKT/FoxO3a, upregulates the expression of the autophagy-related protein LC3B, and downregulates p62 expression, thereby protecting against liver injury induced by type 2 diabetes [94] and CCl_4_ [39]. EGCG promotes autophagy via the ROS/MAPK pathway [95]. Rosmarinic acid enhances autophagy by acting on the JAK2/STAT3 pathway [96].
In summary, most existing studies determine the role of phenolic acids in regulating autophagy by detecting autophagy-related proteins and generally lack comprehensive monitoring of the autophagic process. Moreover, excessive autophagy can lead to cellular self-digestion, making it difficult to determine whether the observed autophagy activation exerts a protective effect. Additionally, vanillic acid alleviates liver fibrosis by inhibiting autophagy in HSCs via the Macrophage Migration Inhibitory Factor (MIF)/Cluster of differentiation 74 (CD74) signaling pathway [97]. This study provides a crucial reverse perspective: it relieves fibrosis by inhibiting autophagy in HSCs, which contrasts sharply with the pro-autophagic effect of most phenolic acids in hepatocytes. Future research should further explore how phenolic acids coordinate the autophagic status of different cell types to achieve overall therapeutic effects in complex systems with multiple liver cell types coexisting, and clarify the precise connection between their upstream signals (e.g., Akt/FoxO3a, ROS/MAPK) and core autophagic mechanisms.
The molecular mechanism by which phenolic acids regulate autophagy is shown in Figure 7.
3.5. Regulating Apoptosis
Apoptosis is the primary mode of cell death in various liver diseases, characterized by an increased protein degradation rate [98,99] and elevated caspase activity [100]. Caspase activation can be triggered by two well-characterized apoptotic pathways: the mitochondria-mediated (intrinsic) pathway and the cell surface death receptor (extrinsic) pathway [101]. Phenolic acids mainly exert anti-apoptotic effects by activating the intrinsic pathway.
The B-cell lymphoma 2 (BCL-2) family consists of proteins with pro-apoptotic and anti-apoptotic properties, and plays a crucial role in the mitochondria-mediated intrinsic pathway [102]. The pro-apoptotic protein BAX induces mitochondrial outer membrane permeabilization, leading to the release of cytochrome c from the mitochondrial intermembrane space into the cytosol [103]. Cytochrome c then binds to Apoptotic protease-activating factor 1 (Apaf-1) to form a multimeric Apaf-1/cytochrome c complex. This complex recruits Procaspase-9, resulting in the formation of the apoptosome [104]. Subsequently, Procaspase-9 is activated via proteolysis [103], leading to its dissociation from the complex, and then activates Caspases 3, 6, and 7 [104]. The extrinsic apoptotic pathway relies on the activation of cell surface death receptors, which induce receptor trimerization and the recruitment of specific intracellular receptor-associated proteins such as Procaspase-8. Procaspase-8 is immediately cleaved into Caspase-8, which activates downstream effector caspases, ultimately leading to cell apoptosis [105].
Studies have shown that phenolic acids can exert hepatoprotective effects in liver injury models induced by drugs, toxic substances, etc., by regulating apoptosis-related proteins. Particularly in DILI, for example, APAP-induced DILI is commonly used to study hepatocellular necrosis. APAP is mainly metabolized into glucuronide and sulfate conjugates; a small portion is converted by the CYP enzyme family into the toxic metabolite N-acetyl-p-benzoquinone imine (NAPQI). NAPQI causes glutathione (GSH) depletion and ROS release, ultimately leading to cell death. (1) Ferulic acid, paeoniflorin, and EGCG can all improve DILI by regulating the expression of apoptosis-related proteins, including BAX, BCL-2, Caspase 3, and Caspase 9 [106,107,108]. Punicalagin can alleviate liver injury caused by acrylamide (ACR), arsenic trioxide (ATO), and methotrexate (MTX) by inhibiting apoptosis [40,109,110]. The combined use of chlorogenic acid, rutin, and quercetin can also reduce triptolide (TP)-induced liver injury by inhibiting apoptosis [111]. (2) In addition to apoptosis induced by chemical substances, veratric acid, paeoniflorin, and chlorogenic acid can also inhibit liver ischemia/reperfusion (I/R) injury, ANIT-induced cholestasis, and liver fibrosis by downregulating BAX, upregulating BCL-2, and reducing the expression of cleaved Caspase-3 and cleaved caspase-9 [50,112,113].
In summary, the mechanisms by which phenolic acids regulate apoptosis all boil down to the modulation of “BAX/BCL-2” and “Caspase-3/9”. Although this demonstrates the hepatoprotective effects of phenolic acids, no studies have clarified how different phenolic acids specifically interfere with the initial upstream signals of this pathway. Secondly, the vast majority of evidence from existing studies comes from DILI models (e.g., APAP-induced), which are characterized by acute cellular necrosis and apoptosis. However, in chronic liver diseases, cell death is a continuous process, and there is currently a lack of robust evidence to confirm whether phenolic acids can still exert protective effects through a single anti-apoptotic mechanism in these more complex pathological environments that are closer to clinical reality. Additionally, there is a dynamic transition between apoptosis and necrosis. In models such as APAP-induced hepatotoxicity, inhibiting the apoptotic pathway may also shift the mode of cell death to more inflammatory necrosis, inadvertently exacerbating liver injury. Existing studies generally fail to simultaneously monitor multiple modes of cell death, which makes the evaluation of the overall biological consequences of the anti-apoptotic effects of phenolic acids incomplete.
The molecular mechanism of phenolic acids inhibiting apoptosis is shown in Figure 8.
3.6. Anti-Fibrosis
Liver fibrosis is caused by excessive production of the extracellular matrix (ECM) and abnormal deposition of fibrous connective tissue in the liver. The massive production and activation of myofibroblasts (MFBs)—which secrete components including collagen, α-smooth muscle actin (α-SMA), and tissue inhibitor of metalloproteinases 1 (TIMP1) [114,115]-are critical to the progression of liver fibrosis, and MFBs mainly originate from the activation of HSCs. Studies have shown that phenolic acids can exert anti-fibrotic effects by regulating the transforming growth factor β (TGF-β)/Smad pathway, HMGB1/TLR4/NF-κB pathway, and targeting the platelet-derived growth factor receptor (PDGFR).
Numerous studies have reported that phenolic acids inhibit liver fibrosis by regulating the TGF-β/Smad pathway. TGF-β is a key cytokine driving liver fibrosis [116]. When injury occurs, the secretion of TGF-β increases in the fibrotic microenvironment and promotes Smad2/3 activation through TGF-β receptor 1/2 (TGFBR1/2) [117]. Subsequently, phosphorylated Smad2/3 forms an oligomeric complex with Smad4, translocates into the nucleus, and regulates the transcription of target genes, thereby mediating fibrosis [118]. Smad7 is a negative feedback regulator of TGF-β signaling [119], and increasing Smad7 expression can delay liver fibrosis [120,121,122]. Ferulic acid can attenuate TGF-β1-induced HSCs activation by inhibiting collagen I, Smad2/3 phosphorylation, and Smad4 signal transduction [123]. Chlorogenic acid inhibits the expression of miR-21, α-SMA, and TGF-β, while increasing the protein expression of Smad7 and matrix metalloproteinase-9 (MMP-9), thereby alleviating liver fibrosis [124]. Echinacoside exerts anti-fibrotic effects by inhibiting the TGF-β1/Smad signaling axis mediated by activin A receptor, type II A (ACVR2A) [125]. In vitro experiments have also confirmed that protocatechuic acid significantly reduces the cell viability of HSCs and downregulates the protein expression of TGF-β and phosphorylated Smad2 [126].
In recent years, PDGFR has become a promising target for anti-fibrotic drugs [127,128]. Activation of the platelet-derived growth factor-BB homodimer (PDGF-BB)/PDGFRβ pathway exhibits a significant pro-fibrotic response [129]. Studies have shown that salvianolic acid B may inhibit HSCs activation by targeting PDGFRβ and significantly reduce the levels of α-SMA and collagen I [130]. Chlorogenic acid prevents liver fibrosis by inhibiting the PDGF-induced pro-fibrotic effect via the NAD(P)H oxidase (NOX)/ROS/MAPK pathway [50].
Phenolic acids can also inhibit the development of liver fibrosis by acting on the HMGB1/TLR4/NF-κB pathway. In addition to excessive ECM accumulation, persistent chronic inflammation also drives fibrosis [131,132]. Existing evidence indicates that inhibiting HMGB1 significantly reduces the inflammatory response and liver fibrosis [133]. Therefore, targeting the signaling pathway between HMGB1 and its receptors may provide a new approach to alleviate liver fibrosis. Isochlorogenic acid and chlorogenic acid can inhibit the protein expression of HMGB1 and TLR4, suppress the overexpression of α-SMA in HSCs, and prevent HSCs activation, thereby alleviating liver fibrosis [134,135]. However, these studies still lack mechanistic depth and cannot confirm that phenolic acids alleviate liver fibrosis by inhibiting HMGB1. In the author’s view, phenolic acids only inhibit the expression of fibrosis-related proteins while reducing inflammation. Since inflammation is one of the drivers of fibrosis, targeting key targets such as TGF-β is obviously a more promising therapeutic strategy. The molecular mechanism of phenolic acid compounds against liver fibrosis is shown in Figure 9.
3.7. Regulating Gut Microbiota and Gut–Liver Axis
The intestinal barrier protects the body from potentially toxic metabolites, bacteria, and their antigens. Therefore, the intestinal barrier and the immune control of the symbiotic microbiota are of crucial importance [136]. Based on the gut–liver axis, the intestinal microbiota regulates pro-inflammatory changes in the liver and intestines, thereby influencing the development of hepatitis, liver fibrosis, cirrhosis, and hepatocellular carcinogenesis [137].
Many phenolic acids contribute to this multifaceted and dynamic host microbe symbiosis. EGCG [138], punicalagin [92], Chicoric Acid [29], TLP-H [139] (gallic acid, corilagin and chebulagic acid) and Chlorogenic [140] can reduce the ratio of Firmicutes/Bacteroidetes and decrease the abundance of co-bacteria and anaerobic bacteria. Caffeic acid also helps balance intestinal microbiota dysbiosis associated with LPS [141]. TLP-H, EGCG, and Chlorogenic acid can also significantly alleviate intestinal dysbiosis and liver inflammation by inhibiting the activation of the TLR4/MyD88/NF-κB signaling pathway, thereby alleviating liver injury [139,140,142]. Chlorogenic acid can increase the expression of occludin and zonula occludens-1 (ZO-1) in intestinal tissues to regulate the intestinal microbiota; in addition, it can reduce the level of LPS in the portal vein and increase the level of glucagon-like peptide-1 (GLP-1) to exert anti-inflammatory effects [30].
In summary, we observe that phenolic acid treatment is accompanied by improved microbiota composition, intestinal barrier repair, and alleviated liver injury. However, very few studies have directly confirmed that specific microbiota changes are a necessary cause for the hepatoprotective effects of phenolic acids through experiments such as fecal microbiota transplantation or gene-knockout animal models. Furthermore, an unavoidable translational medicine challenge lies in the contradiction between the bioavailability of phenolic acids and their local intestinal concentration. Many phenolic acids are poorly absorbed in the upper intestine, which, conversely, allows them to reach the large intestine at higher concentrations to interact with the microbiota. Future research must clarify whether their hepatoprotective effects rely on systemic pharmacological effects after absorption, or on influencing the liver through signal transmission (e.g., via GLP-1) or metabolite absorption after exerting local effects in the intestine. This is crucial for deciding whether to develop derivatives that enhance systemic exposure or to design formulations targeting colonic delivery. The molecular mechanism of phenolic acids regulating gut microbiota is shown in Figure 10.
3.8. Other Mechanisms
In addition to the mechanisms mentioned above, phenolic acids can also exert hepatoprotective effects through biological processes such as epigenetic regulation, endoplasmic reticulum (ER) stress modulation, ferroptosis inhibition, suppression of HCC cell proliferation, promotion of liver repair and regeneration, and pyroptosis regulation.
4. Clinical Translation Prospects of Phenolic Acids
Phenolic acids are abundant in plant-derived foods and exhibit high safety. For example, chlorogenic acid is one of the most accessible phenolic acid compounds; it exists naturally and in high concentrations in raw coffee extracts and tea [143], and possesses pharmacological activities such as antibacterial, antiviral, and antioxidant effects. Grains like rice, wheat, and oats are the main sources of ferulic acid, which is also present in various fruits and vegetables. In Japan, ferulic acid is used as a food additive, antioxidant, and food preservative, with a long history of safe consumption [144]. Various berries, such as blueberries, are rich in gallic acid, caffeic acid, protocatechuic acid, and more [145]. However, this does not mean that phenolic acids are absolutely safe when used as therapeutic drugs. Studies have shown that rosmarinic acid is non-toxic to humans, even when patients take a dose of 500 mg/day [146]; yet, the safety of its long-term application remains unknown.
Low oral bioavailability is a common challenge faced by most phenolic acids. Due to their large molecular weight and high polarity, some phenolic acids are difficult to pass through the intestinal mucosa via passive diffusion, resulting in poor intestinal absorption. Those phenolic acids that are absorbed undergo Phase II conjugation metabolism in the intestines and liver, converting into metabolites that are excreted from the body. This leads to extremely low concentrations of the parent drug entering the systemic circulation. To address this bottleneck, numerous studies have shown that advanced dosage forms such as nanotechnology (e.g., liposomes, polymeric nanoparticles), phospholipid complexes, and emulsion-based delivery systems can be used to improve the bioavailability of phenolic acids. Chitosan is a naturally occurring biopolymer; the binding of natural bioactive molecules (e.g., polyphenols) to chitosan can enhance bioactivity and water solubility [147]. Research has demonstrated that the caffeic acid-chitosan conjugate exerts a significant hepatoprotective effect against ethanol-induced hepatotoxicity in mice [148]. Additionally, with the development of nanotechnology, the pharmaceutical and biotechnology industries have gradually explored new directions to substantially improve the efficacy of antioxidants. Gold nanoparticles (AuNPs) have attracted great interest due to their biocompatibility, colloidal stability, water solubility, and catalytic activity. SAuPTB (AuNP-tannic acid hybrid nanozymes) exhibit strong antioxidant activity [149].
Significant pharmacokinetic differences in phenolic acids exist between humans and animals, this means that dose conversion between animals and humans cannot simply rely on conversion formulas. Studies have shown that the maximum therapeutic dose of chlorogenic acid in a rat model of liver fibrosis is 60 mg/kg, which converts to an approximate adult equivalent dose of 580 mg/day—far higher than the daily dietary intake of humans. In a randomized, double-blind clinical trial (n = 101), researchers administered chlorogenic acid (200 mg/day) to patients with non-alcoholic fatty liver disease (NAFLD) for 6 months. They found that compared with the placebo group, there was no significant reduction in liver biochemical indicators; however, this dose was completely safe without any side effects [150]. Therefore, one of the top priorities of future clinical research is to determine the true pharmacokinetic characteristics of phenolic acids in the human body. It may also be necessary to improve their bioavailability through novel delivery systems, thereby making the effective doses based on animal models truly clinically relevant.
Although numerous animal and cell experiments have confirmed the hepatoprotective effects of phenolic acids, it is undeniable that design flaws in preclinical studies exist. Widely used cell lines (e.g., HepG2, HEK293) undergo genetic and phenotypic drift during long-term passage, and their biological characteristics differ significantly from primary human cells. Animal models typically “induce” diseases in a short period through gene knockout or strong chemical/physical stimulation. For instance, CCl_4_ injection or bile duct ligation can induce liver fibrosis within a few weeks. This differs from the multifactorial, long-term, and chronic disease progression process in humans and cannot fundamentally replicate the actual disease development. In animal studies, drugs are usually administered either simultaneously with model induction (preventive use) or immediately after model establishment (therapeutic use). In contrast, clinical patients typically begin treatment only after the disease has been established, or even after fibrosis or carcinogenesis has occurred. Organoid [151] and Patient-Derived Tumor Xenograft (PDX) [152] models can enhance the clinical relevance of research.
5. Discussion
Liver diseases pose a significant challenge to global public health, characterized by complex pathogenic mechanisms and limited therapeutic options. In recent years, phenolic acid compounds have emerged as a research hotspot in liver disease studies due to their diverse pharmacological activities. In clinical settings, there have been epidemiological studies on the association between phenolic acid intake and metabolic diseases such as diabetes [153], hypertension [154], and metabolic syndrome [155]. There are few clinical studies on phenolic acids for liver diseases beyond their use as dietary supplements. In a cross-sectional study, the correlation between phenolic acid intake and the risk of NAFLD was analyzed among 9894 enrolled adults. The results showed that participants with higher dietary phenolic acid intake had a lower risk of developing NAFLD [156]. Some Chinese patent medicines rich in salvianolic acids, such as Danshen Tablets and Salvia Miltiorrhiza Ligustrazine Injection, have been clinically used for liver diseases including hepatitis and fatty liver. Although clinical applications of individual phenolic acid monomers are limited, these findings highlight the potential importance of phenolic acid-rich dietary patterns as a component of strategies to reduce liver disease risk.
This article reviews the pharmacological reports on the hepatoprotective effects of phenolic acids in vivo and in vitro. Among them, hydroxycinnamic acids have been the most extensively studied, including caffeic acid, ferulic acid, rosmarinic acid, and chlorogenic acid. For hydroxybenzoic acids, EGCG has garnered the widest research attention. These compounds are not only effective in various liver injury models but, more importantly, can simultaneously regulate multiple core nodes of disease networks, making them recommended as priority candidates for in-depth development as potential drugs.
Prioritize the Development of the Most Promising Lead Compounds. Our analysis reveals that chlorogenic acid and EGCG have the most extensive research and relatively abundant evidence in the field of hepatoprotection. Existing literature has comprehensively summarized their dietary sources, bioavailability, pharmacological effects, and potential targets. However, clinical trial data on these compounds remain scarce. In a randomized controlled trial, NAFLD patients were administered Altilix^®^ (Bionap, Belpasso, Catania, Italy; a formulation containing chlorogenic acid and its derivatives, as well as luteolin and its derivatives) for 6 months. Compared with the placebo group, significant reductions in liver function indicators were observed [157]. Meanwhile, active exploration of the synergistic therapeutic potential of standardized extracts rich in multiple phenolic acids (e.g., Altilix^®^) is warranted, as such multi-component formulations may offer advantages over single-component agents.
Focus on the Most Core and Validated Mechanisms. At present, the primary target cells and specific key molecular targets of phenolic acids for hepatoprotection remain unclear. Taking chlorogenic acid, the most extensively studied phenolic acid, as an example, it exhibits various pharmacological effects such as anti-inflammation, antioxidant activity, regulation of lipid metabolism, and anti-liver fibrosis. However, compared with other phenolic acids that also possess these pharmacological effects, this does not reflect the specificity of the regulatory targets of phenolic acids, nor how different phenolic acids precisely and preferentially regulate specific links among these processes. In recent years, research on the gut-liver axis has advanced significantly. Phenolic acids can exert broad-spectrum therapeutic effects by improving intestinal microbiota dysbiosis, enhancing intestinal barrier function, and regulating bile acid metabolism. Furthermore, based on individual differences in intestinal microbiota, the development of combined phenolic acid-prebiotic [158] therapies may be feasible in the future. These findings suggest that attention should be paid to crosstalk between multiple organs. Integrating multi-omics technologies (e.g., proteomics and transcriptomics) with clinical cases will help identify new key targets, facilitate in-depth mechanistic studies, and achieve precise regulation.
Overcoming Core Bottlenecks in Clinical Translation. Optimize delivery systems to address bioavailability challenges. Priority must be given to developing liver-targeted delivery systems based on nanotechnology (e.g., liposomes, polymeric nanoparticles) to increase their accumulation concentration in the liver. Conduct standardized preclinical and clinical studies. Before initiating human trials, systematic toxicological and pharmacokinetic evaluations complying with Good Laboratory Practice (GLP) standards must be completed. Subsequently, rigorous Phase I and Phase II clinical trials should be designed to first verify the safety and preliminary efficacy of phenolic acids in high-risk populations with NAFLD or DILI. Advance precision medicine and rational combination therapy. Meanwhile, explore combination therapy strategies of phenolic acids with existing standard drugs (e.g., metformin, obeticholic acid) to exert synergistic effects, reduce the dosage of each drug, and minimize toxic side effects. The possible mechanisms by which phenolic acids modulate liver diseases are shown in Table 1.
Therefore, we believe that future translational research and clinical trials can prioritize focusing on widely studied phenolic acids such as chlorogenic acid and EGCG, taking them as “lead compounds” for in-depth development and application exploration. At the same time, on the basis of promoting research on phenolic acid combination therapy and the application of dietary supplements, attention should also be paid to obtaining nutrients, antioxidants, bioactive compounds, or phytochemicals from a balanced diet, so as to achieve optimal nutrition, health, and well-being.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Devarbhavi H. Asrani S.K. Arab J.P. Nartey Y.A. Pose E. Kamath P.S. Global burden of liver disease: 2023 update J. Hepatol.20237951653710.1016/j.jhep.2023.03.01736990226 · doi ↗ · pubmed ↗
- 2Xiao J. Wang F. Wong N.K. He J. Zhang R. Sun R. Xu Y. Liu Y. Li W. Koike K. Global liver disease burdens and research trends: Analysis from a Chinese perspective J. Hepatol.20197121222110.1016/j.jhep.2019.03.00430871980 · doi ↗ · pubmed ↗
- 3Andrade R.J. Chalasani N. Björnsson E.S. Suzuki A. Kullak-Ublick G.A. Watkins P.B. Devarbhavi H. Merz M. Lucena M.I. Kaplowitz N. Drug-induced liver injury Nat. Rev. Dis. Primers 201955810.1038/s 41572-019-0105-031439850 · doi ↗ · pubmed ↗
- 4Camini F.C. Costa D.C. Silymarin: Not just another antioxidant J. Basic Clin. Physiol. Pharmacol.2020312019020610.1515/jbcpp-2019-020632134732 · doi ↗ · pubmed ↗
- 5Song P. Zhang X. Feng W. Xu W. Wu C. Xie S. Yu S. Fu R. Biological synthesis of ursodeoxycholic acid Front. Microbiol.202314114066210.3389/fmicb.2023.114066236910199 PMC 9998936 · doi ↗ · pubmed ↗
- 6Sun Y. Ji X. Cui J. Mi Y. Zhang J. Guo Z. Synthesis, Characterization, and the Antioxidant Activity of Phenolic Acid Chitooligosaccharide Derivatives Mar. Drugs 20222048910.3390/md 2008048936005492 PMC 9410539 · doi ↗ · pubmed ↗
- 7Simón J. Casado-Andrés M. Goikoetxea-Usandizaga N. Serrano-MaciáM. Martínez-Chantar M.L. Nutraceutical Properties of Polyphenols against Liver Diseases Nutrients 202012351710.3390/nu 1211351733203174 PMC 7697723 · doi ↗ · pubmed ↗
- 8Ordoudi S.A. Tsimidou M.Z. Crocin bleaching assay (CBA) in structure-radical scavenging activity studies of selected phenolic compounds J. Agric. Food Chem.2006549347935610.1021/jf 062115 d 17147417 · doi ↗ · pubmed ↗