ICECleSHZ29: Novel Integrative and Conjugative Element (ICE)-Carrying Tigecycline Resistance Gene tet(X6) in Chryseobacterium lecithinasegens
Xi Chen, Yifei Zhang, Chunling Jiang, Yafang Lin, Xiaohui Yao, Wansen Nie, Lin Li, Jianchao Wei, Donghua Shao, Ke Liu, Zongjie Li, Yafeng Qiu, Zhiyong Ma, Beibei Li, Lining Xia

TL;DR
A new ICE carrying the tigecycline resistance gene tet(X6) was discovered in a multidrug-resistant Chryseobacterium lecithinasegens strain from China.
Contribution
This is the first report of tet(X6) in Chryseobacterium lecithinasegens and the characterization of its novel ICE carrier, ICECleSHZ29.
Findings
ICECleSHZ29 is a 74,906 bp ICE carrying tet(X6) and multiple resistance genes.
ICECleSHZ29-like ICEs were classified into two types based on insertion sites.
Conjugation experiments failed to transfer ICECleSHZ29 to Escherichia coli.
Abstract
Background/Objectives: The global dissemination of tet(X) variants critically threatens tigecycline efficacy as a last-resort antibiotic. The aim of this study was to characterize a tet(X6)-carrying integrative and conjugative element (ICE) in a multidrug-resistant Chryseobacterium lecithinasegens strain, SHZ29, isolated from Shanghai, China. Methods: Minimum inhibitory concentrations (MICs) were determined by broth microdilution for SHZ29. Whole genomic sequencing and bioinformatic analysis were performed to depict the structure of the novel tet(X6)-carrying ICE. Inverse PCR and conjugation experiments were conducted to investigate the transfer ability of the ICE. Results: We depicted a novel tet(X6)-carrying ICE, named ICECleSHZ29, which is 74,906 bp in size and inserted into the 3′ end of tRNA-Met-CAT gene of the C. lecithinasegens strain SHZ29, with 17 bp direct repeats…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
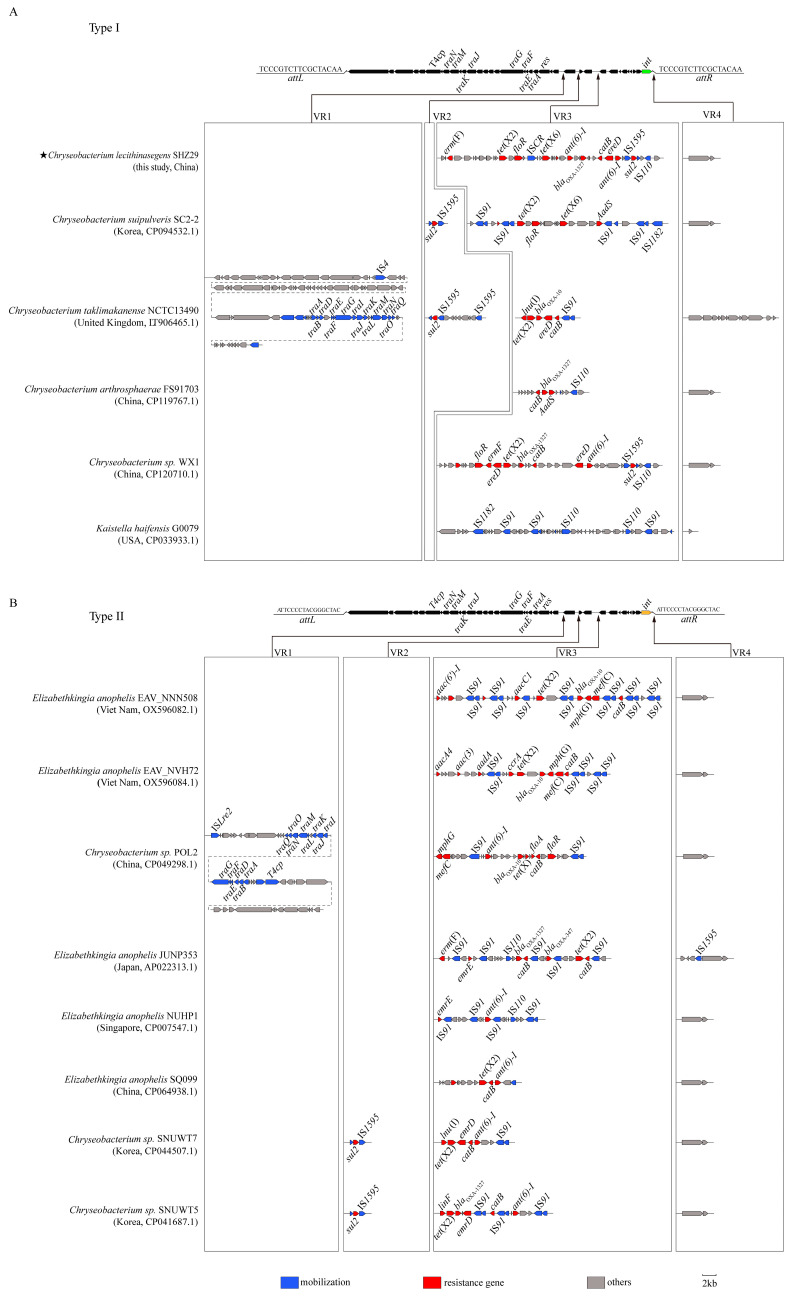 Figure 1
Figure 1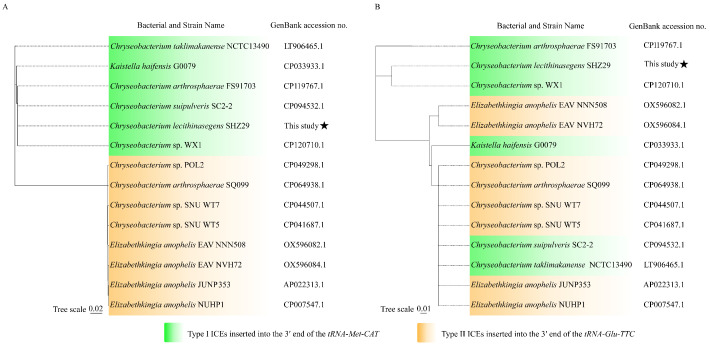 Figure 2
Figure 2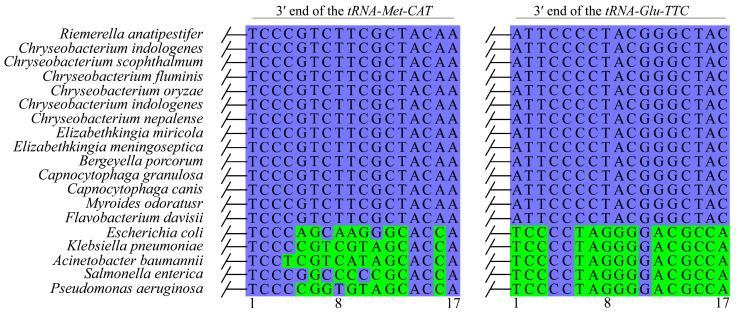 Figure 3
Figure 3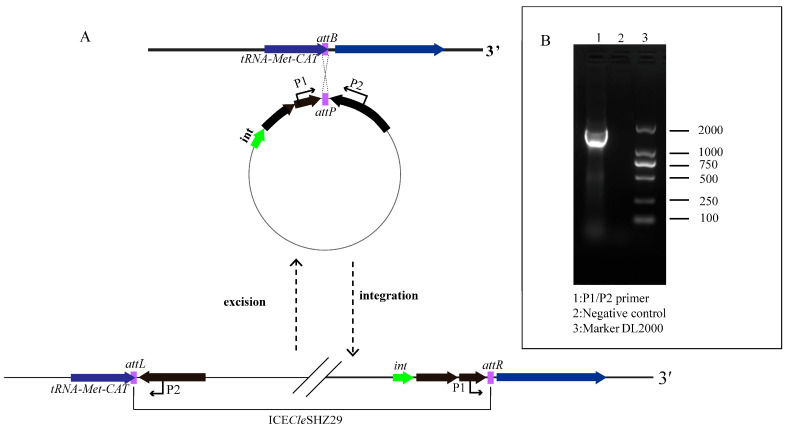 Figure 4
Figure 4- —Shanghai Agriculture Applied Technology Development Program, China
- —National Natural Science Foundation of China
- —Xinjiang Uygur Autonomous Region “Tianshan Talents” Cultivation Program—“Three Rural” Key Talent Development Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInfections and bacterial resistance · Antibiotic Resistance in Bacteria · Cancer Research and Treatments
1. Introduction
The rapid dissemination of mobile antibiotic resistance genes represents a critical challenge to global public health [1]. Integrative and conjugative elements (ICEs), as a class of mobile genetic elements, play an important role in the horizontal transfer of antibiotic resistance genes [2]. They can autonomously excise from the host chromosome, transfer via conjugation in an intermediate circular form, and site-specifically insert into and replicate within a new host chromosome [3,4]. A number of clinically significant resistance genes have been found to be harbored and disseminated by ICEs, which poses serious challenges to bacterial infection treatment. For instance, the oxazolidinone resistance gene optrA has been documented to be harbored by the novel ICE ICESsu988S in Streptococcus suis, which also co-harbors the macrolide resistance gene erm(T) and enables horizontal transfer of multidrug-resistance among isolates [5]. The colistin resistance gene mcr-1 was identified on the ICE ICEAsp1 in a swine-origin Actinobacillus species in an Actinobacillus species of swine origin. This ICE contains a multidrug resistance region, and multiple ISApl1 elements mediate the formation of composite transposons to promote antibiotic resistance dissemination [6]. The carbapenem resistance gene blaNDM-1 has been identified on two novel SXT/R391 ICEs, namely ICEPmiChnXH1653 and ICEPvuChnBC22 in Proteus mirabilis and P. vulgaris, respectively. ICEPmiChnXH1653 in P. mirabilis harbors two tandem copies of blaNDM-1 along with 21 other antibiotic resistance genes. ICEPvuChnBC22 in P. vulgaris carries blaNDM-1 along with 20 other resistance genes, including cfr and aac(6′)-Ib-cr. These data indicate that ICEs serve as a key platform for the horizontal transfer of not only the blaNDM-1 gene, but also other multidrug-resistance genes [7,8].
Tigecycline is regarded as a “last-resort” antibiotic for treating multidrug-resistant Gram-negative bacterial infections [9]. However, the emergence and dissemination of plasmid-mediated mobile tigecycline resistance gene tet(X) variants severely compromises the clinical efficacy of tigecycline and poses a significant threat to global public health [10]. To date, at least 22 tet(X) variants have been reported [11]. Of them, the tet(X6) gene was initially identified on an SXT/R391 ICE, ICEPgs6Chn1, in Proteus species [12]. Since then, it has been detected across a wide range of bacteria such as Proteus, Acinetobacter, Escherichia, and Sphingobacterium [13]. This gene has been documented to be present on a diverse range of ICEs. For instance, tet(X6) was identified on an SXT/R391-family ICE within the chromosome of P. cibarius isolate, where it co-occurs with a novel efflux pump gene cluster, tnfxB3–tmexCD3–toprJ1b, another tigecycline resistance determinant [14]. A circularizable ICE, designated ICEEmeChn3, that harbors the tet(X6) gene has been identified in Elizabethkingia meningoseptica. This element facilitates mobilization and recombination through ISwz1-mediated rearrangements, promoting the dissemination of high-level tigecycline resistance from aquaculture settings into natural ecosystems [15]. Beyond its association with ICEs, the tet(X6) gene has also been found to be harbored by various types of plasmids [16]. For example, the tet(X6) gene was identified on broad-host-range IncA/C_2_-type plasmids in animal-derived Proteus spp. [17]. In an A. baumannii isolate of chicken origin, a resistant Pseudomonas aeruginosa strain from poultry, the tet(X6) gene was located on an Inc_pRBL16_ plasmid, co-harboring blaIMP-45 and tmexCD3–toprJ3, which together conferred resistance to last-resort agents including tigecycline and carbapenems [18].
The genus Chryseobacterium are non-fermentative, oxidase-positive Gram-negative bacilli that are widely distributed in natural environments, particularly in water and soil [19]. Although Chryseobacterium species are generally considered low-virulence opportunistic pathogens, some members of them can cause severe clinical infections in immunocompromised patients, including respiratory tract infections, bloodstream infections, meningitis, and urinary tract infections. For instance, C. gleum has been reported to cause respiratory infections in neonates [19], while C. indologenes has been associated with urinary tract infections and other diseases [20]. Bacteria of this genus has been found to harbor various resistance genes, including several novel ones, thereby suggesting its potential role as a significant reservoir of antibiotic resistance [21]. For instance, the ICE ICECiPOL15, identified in C. indoltheticum, carries the β-lactam resistance gene bla_AQU_ and has been experimentally demonstrated to transfer via conjugation [22]. ICECspPOL2, identified in C. sp. POL2, harbors multiple antibiotic resistance genes, including a functional carbapenemase gene [23].
In the present study, we reported, for the first time, the identification of the tet(X6) gene in C. lecithinasegens. Moreover, we characterized the genetic structure and mobile ability of the tet(X6) carrier, a novel ICE, namely ICECleSHZ29.
2. Results and Discussion
2.1. C. lecithinasegens SHZ29 Is a Multidrug-Resistant Strain
C. lecithinasegens SHZ29 was isolated from a fecal sample obtained from a pig farm in Shanghai, China. MIC determination revealed that C. lecithinasegens SHZ29 exhibited resistance to tigecycline (MIC, 16 μg/mL), tetracycline (MIC, 256 μg/mL), ceftriaxone (MIC, 64 μg/mL), meropenem (MIC, 16 μg/mL), amikacin (MIC, 256 μg/mL), ciprofloxacin (MIC, 256 μg/mL), florfenicol (MIC, 128 μg/mL), and colistin (MIC, 256 μg/mL). To investigate the molecular mechanism of tigecycline resistance in C. lecithinasegens SHZ29, whole-genome sequencing was performed and analysis of the resistance gene profile showed that C. lecithinasegens SHZ29 harbored tet(X2) and tet(X6) genes. Both tet(X2) and tet(X6) have been demonstrated to confer resistance to tigecycline [12,13,24,25]. However, a study has shown that tet(X2) was not transcribed and did not contribute to tigecycline resistance in C. indologenes [26]. Comparative analysis revealed that the tet(X2) gene in C. lecithinasegens SHZ29 shared an identical ribosome-binding site (RBS) upstream with that of the aforementioned C. indologenes strain, suggesting that the tet(X2) gene in C. lecithinasegens SHZ29 is also non-functional [26]. The tet(X6) gene, originally identified on ICEPgs6Chn1 in Proteus spp., confers high-level tigecycline resistance across diverse bacterial species [11]. To date, several naturally occurring variants of tet(X6) have been reported. In C. lecithinasegens SHZ29, the tet(X6) gene exhibits four amino acid substitutions compared to the tet(X6) prototype [13]. Functional studies have confirmed that this variant retains the ability to confer tigecycline resistance [13]. It is therefore likely that the tet(X6) variant is the primary determinant of tigecycline resistance in C. lecithinasegens SHZ29. Besides tet(X2) and tet(X6), C. lecithinasegens SHZ29 harbored various other resistance genes, including macrolide resistance genes erm(F) and ere(D), the β-lactam resistance gene blaOXA-1327, phenicol resistance genes floR and catB, the sulfonamide resistance gene sul2, the carbapenem resistance gene blaIND-14, and the aminoglycoside resistance gene ant(6)-I.
2.2. Structure of ICECleSHZ29
To investigate the genetic environment of tet(X6) in C. lecithinasegens SHZ29, the contig harboring tet(X6) and its flanking contigs were assembled via PCR gap filling. A novel integrative and conjugative element (ICE), designated ICECleSHZ29, was identified. ICECleSHZ29 is 74,906 bp in size with a G/C content of 38%. The element is flanked by a 17 bp perfect direct repeat (5′-tcccgtcttcgctacaa-3′) at both ends and is inserted into the 3′ end of the tRNA-Met-CAT gene. RAST annotation identified 90 open reading frames (ORFs) within ICECleSHZ29. Among these, some ORFs were predicted to encode core functional components of ICE, including those involved in site-specific recombination and integration (integrase and resolvase) and conjugative transfer (TraAEGJKMN and T4CP). Furthermore, ICECleSHZ29 harbored a gene cluster consisting of nine antibiotic resistance genes, including tet(X2), tet(X6), erm(F), ere(D), floR, catB, sul2, ant(6)-I (two copies), and blaOXA-1327. Within this cluster, three intact insertion sequences (IS110, IS1595, and ISCR), one truncated IS1595 element, and several other ORFs encoding hypothetical proteins are interspersed among the aforementioned resistance genes (Figure 1A).
2.3. Phylogentic Analysis of ICECleSHZ29
BLAST analysis of the ICECleSHZ29 revealed high homology with ICEs from multiple genera within the Weeksellaceae family, including Elizabethkingia, Chryseobacterium, and Kaistella. This family of ICEs shares a conserved backbone of about 38 kb and contains four variable regions (VR1-VR4) (Figure 1). In VR1, The ICEs from C. taklimakanense NCTC13490 (GenBank accession no: LT906465) and C. sp. POL2 (GenBank accession no: CP049298) contained several conjugative transfer-related proteins. For VR2, the ICEs from C. suipulveris SC2-2 (GenBank accession no: CP094532.1), C. taklimakanense NCTC13490, C. sp. SNU WT5 (GenBank accession no: CP041687.1), and C. sp. SNU WT7 (GenBank accession no: CP064938.1) carried the sulfonamide resistance gene sul2, flanked by IS1595. The VR3 region serves as an aggregation hub for antibiotic resistance genes (Figure 1). For instance, the ICE derived from C. suipulveris SC2-2 harbored the tet(X6) gene, while ICEs from C. taklimakanense NCTC13490, E. anophelis EAV_NNN508 (GenBank accession no: OX596082.1), E. anophelis EAV_NVH72 (GenBank accession no: OX596084.1), C. sp. POL2, and C. sp. WX1 (GenBank accession no: CP120710.1) encoded carbapenem resistance genes, such as blaOXA-10 or blaOXA-347. Moreover, similar to ICECleSHZ29, the VR3 region of these ICEs harbored diverse IS elements and intact or truncated transposase genes interspersed among these antibiotic resistance genes. The abundance of transfer-associated elements may facilitate the accumulation of antibiotic resistance genes, thereby promoting the formation of resistance gene clusters within the VR3 regions of these ICEs [27,28]. The contents of VR4 exhibit a high degree of diversity and primarily encode hypothetical proteins of unknown function.
Comparative analysis of these ICECleSHZ29-like ICEs revealed that they shared a conserved structural backbone. However, further bioinformatic analysis demonstrated that these ICEs could be classified into two distinct types based on their distinct insertion sites, designated as Type I and Type II (Figure 1). Type I ICEs insert at the 3′ end of the tRNA-Met-CAT gene, forming a perfect 17 bp direct repeat (5′-tcccgtcttcgctacaa-3′) at both insertion junctions (Figure 1A). By contrast, Type II ICEs insert at the 3′ end of the tRNA-Glu-TTC gene, generating a perfect 17 bp direct repeat (5′-attcccctacgggctac-3′) at both insertion junctions (Figure 1B).
Integrases and relaxases play critical roles in the excision and integration processes during ICE transfer [29]. Integrases, as core components of the ICE recombination module, mediate ICE integration into host chromosomes and, upon activation, cooperate with recombination directionality factors (RDFs) for ICE excision. Relaxases, central to the conjugation module, recognize the origin of transfer (oriT), cleave ICE DNA to generate single-stranded intermediates, and further facilitate their horizontal transfer via the type IV secretion system (T4SS) [30]. To investigate the insertion site divergence within ICECleSHZ29-like ICEs, we extracted the amino acid sequences of integrase and relaxase from the 14 ICEs and performed multiple sequence alignment and phylogenetic analysis. Sequence comparison revealed 4–5 amino acid differences among Type I integrases, whereas Type II integrases showed 100% amino acid identity. Substantial divergence (>100 amino acid differences) was found between Type I and Type II integrases, resulting in their clustering within distinct phylogenetic clades (Figure 2A and Supplementary Figure S1). Unlike the integrases, the relaxases of Type I and Type II ICEs did not exhibit distinct phylogenetic clustering by type. Instead, they were distributed across three divergent clades, displaying an interspersed distribution pattern (Figure 2B and Supplementary Figure S2). Previous studies have demonstrated that integrases of different types, as well as variants of different subtypes within the same family, can lead to variations in the insertion sites of ICE [31,32,33]. Taken together, the distinct insertion sites of Type I and Type II ICEs may be attributed to differences in integrases, despite sharing a conserved backbone.
To investigate the potential bacterial host range of ICECleSHZ29-like ICEs, we first performed BLAST analysis of the 3′ ends of tRNA-Met-CAT and tRNA-Glu-TTC genes, which are the attachment sites (attB sites) of this ICE family. These two attB sequences perfectly matched the 3′ ends of tRNA-Met-CAT and tRNA-Glu-TTC genes in bacterial species from the families Flavobacteriaceae and Weeksellaceae, but exhibited certain discrepancies with the 3′ ends of these two tRNA genes in other bacterial species. We then conducted a sequence comparison of the attB sites across diverse bacterial species (Figure 3). Representative species of the families of Flavobacteriaceae and Weeksellaceae, as well as several clinically significant Gram-negative pathogenic bacteria, including E. coli, Klebsiella pneumonia, A. baumannii, Salmonella enterica, and P. aeruginosa, were included in the analysis. Comparative analysis revealed fully conserved 3′-end sequences in tRNA-Met-CAT and tRNA-Glu-TTC genes across representative species of the Flavobacteriaceae and Weeksellaceae families (Figure 3). In contrast, the 3′ ends of the two tRNA genes of the five pathogenic bacteria exhibited significant differences compared with those in the families of Flavobacteriaceae and Weeksellaceae. This observation indicated that the ICECleSHZ29-like ICEs only possess the capacity for transfer exclusively among bacterial species within the families Flavobacteriaceae and Weeksellaceae, which may also explain why this ICE family has thus far been exclusively identified in Weeksellaceae in NCBI databases.
2.4. Detection of Circular Intermediate Form
Inverse PCR was conducted using the primer set P1/P2, with the genomic DNA of C. lecithinasegens SHZ29 serving as the template. The amplification yielded a positive result, indicating that ICECleSHZ29 is capable of excising itself from the chromosome and forming an intermediate circular structure (Figure 4). However, the conjugation experiment using E. coli EC600 as the recipient strain failed. As mentioned above, E. coli lacks the attB site of ICECleSHZ29, which may account for the failure of conjugative transfer. ICECleSHZ29 was originally isolated from C. lecithinasegens, a member of the Weeksellaceae family, and related ICEs often show host specificity for taxa closely related to their native hosts. To evaluate the transfer capacity of ICECleSHZ29, a conjugation experiment should be performed using bacterial species within the families Flavobacteriaceae and Weeksellaceae. Unfortunately, these strains were not available to us. Interestingly, a previous study demonstrated that ICECspPOL2, a type II ICECleSHZ29-like ICE, fails to transfer to E. coli but successfully conjugates into specific Elizabethkingia species [23]. These findings collectively substantiate that ICECleSHZ29-like ICEs exhibit transfer competence exclusively in bacterial hosts within the Flavobacteriaceae and Weeksellaceae families.
3. Materials and Methods
3.1. Bacteria Strain and Antimicrobial Susceptibility Testing
During routine surveillance of tigecycline-resistant bacteria in 2024, a C. lecithinasegens strain designated SHZ29 was isolated from a fecal sample collected from a swine farm in Shanghai, China. Minimum inhibitory concentrations (MICs) were determined by broth microdilution according to the CLSI document M100-S30 [34,35,36]. Briefly, cation-adjusted Mueller–Hinton broth containing serially diluted test antibiotics was inoculated with the bacterial suspension at 10^5^ CFU mL^−1^. Each plate included positive controls (bacteria only) and negative controls (broth only). After incubation at 35 ± 2 °C for 16–20 h, MICs were defined as the lowest concentration of antibiotics that inhibited visible bacterial growth. To ensure the reliability of the results, we performed the MIC test with three replicates. The following antibiotics were tested: tigecycline, tetracycline, ceftriaxone, meropenem, amikacin, ciprofloxacin, florfenicol, and colistin. Given the absence of species-specific interpretive criteria for Chryseobacterium spp., resistance breakpoints for tetracycline, amikacin, ciprofloxacin, ceftriaxone, and meropenem were applied according to CLSI guidelines for “other non-Enterobacteriaceae” as described previously [26,37]. For tigecycline, breakpoints defined by the European Committee on Antimicrobial Susceptibility Testing (EUCAST) were utilized. Florfenicol resistance was interpreted using veterinary standards (CLSI VET08) [38]. Quality control strains E. coli ATCC 25,922 and P. aeruginosa ATCC 27,853 were included in all assays.
3.2. Whole Genome Sequencing and Bioinformatic Analyses
Genomic DNA of C. lecithinasegens SHZ29 was extracted using the TIANamp Bacterial Genomic DNA Kit (Tiangen Biotech Co., Ltd., Beijing, China). Whole genome sequencing was performed using the Illumina Hiseq 2000 platform (Majorbio, Shanghai, China), and the obtained raw data were assembled using SOAPdenovo2 [39]. Resistance genes and ICEs were identified using ResFinder [40] and ICEfinder [41], respectively. To obtain the complete sequence of the tet(X6)-carrying ICE, a gap-filling strategy was employed. Briefly, the contig containing tet(X6) was identified and used as a query in BLASTn against the NCBI nucleotide database to identify structurally related ICEs as references. All contigs from the WGS assembly were then mapped to these reference ICE sequences to identify potentially tet(X6)-associated contigs. Specific primer pairs were designed to span the gaps between these contigs, followed by PCR amplification (primer set P14/P16, Supplementary Table S1) and Sanger sequencing of the amplicon. The resulting amplicon sequence was then assembled with the existing tet(X6)-containing contigs to produce the closed, full-length ICE sequence. The ICE annotation was performed with the RAST server (https://rast.nmpdr.org/), followed by manual verification against the NCBI database (https://www.ncbi.nlm.nih.gov/). Sequence comparison was performed using BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 24 December 2024).
3.3. Phylogenetic Analysis and Sequence Comparison
Phylogenetic analysis was performed for the integrases and relaxases of the ICECleSHZ29 and another 13 relevant ICEs with similar backbones. A phylogenetic tree was constructed by the neighbor-joining method using MEGA11 [42]. Bootstrap values were calculated with 1000 replications. In addition, the 3′ end nucleotide sequences of the tRNA-Met-CAT and tRNA-Glu-TTC of different bacteria species were collected and compared to explore the transfer potential of the ICECleSHZ29-like ICEs.
3.4. Detection of Circular Intermediates of ICECleSHZ29 and Conjugation Experiments
Circular intermediates of ICECleSHZ29 were detected by inverse PCR using primer set P1 (5′-GTGGGAACAGAAAGCGAA-3′)/P2 (5′-CTACAAACCGGAAGAAGTCG-3′). The amplicon sequence was obtained by Sanger sequencing and subjected to alignment with the attL and attR of ICECleSHZ29. To evaluate the mobility of ICECleSHZ29, conjugation experiments were conducted using rifampicin-resistant E. coli EC600 as the recipient strain. Conjugation experiments were performed as previously described with some modifications [43]. Briefly, donor and recipient strains with overnight growth were combined at a 1:1 ratio. Following centrifugation at 4000× g for 10 min, the cell pellet was resuspended in 100 μL of LB broth and spotted onto non-selective Tryptic Soy Agar (TSA) plates for mating. After incubation at 37 °C for 6 h, transconjugants were isolated on LB agar plates containing rifampicin (100 mg/L) and tigecycline (2 μg/mL).
3.5. Nucleotide Sequence Accession Number
The 74906 bp nucleotide sequence of ICECleSHZ29 has been deposited in the GenBank database and assigned the accession No. PRJNA1264973.
4. Conclusions
In conclusion, we identified a novel tet(X6)-carrying ICE, ICECleSHZ29, in a tigecycline-resistant C. lecithinasegens isolate of swine origin. Further studies are warranted to investigate the presence and prevalence of tet(X6) and ICECleSHZ29-like elements in Chryseobacterium species, as well as more broadly in the families Flavobacteriaceae and Weeksellaceae.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1He L.Y. Ying G.G. Liu Y.S. Su H.C. Chen J. Liu S.S. Zhao J.L. Discharge of swine wastes risks water quality and food safety: Antibiotics and antibiotic resistance genes from swine sources to the receiving environments Environ. Int.201692–9321021910.1016/j.envint.2016.03.02327107226 · doi ↗ · pubmed ↗
- 2Burrus V. Waldor M.K. Shaping bacterial genomes with integrative and conjugative elements Res. Microbiol.200415537638610.1016/j.resmic.2004.01.01215207870 · doi ↗ · pubmed ↗
- 3Burrus V. Marrero J. Waldor M.K. The current ICE age: Biology and evolution of SXT-related integrating conjugative elements Plasmid 20065517318310.1016/j.plasmid.2006.01.00116530834 · doi ↗ · pubmed ↗
- 4Wozniak R.A. Waldor M.K. Integrative and conjugative elements: Mosaic mobile genetic elements enabling dynamic lateral gene flow Nat. Rev. Microbiol.2010855256310.1038/nrmicro 238220601965 · doi ↗ · pubmed ↗
- 5Zhang J. Yang Y. Sun H. Luo X. Cui X. Miao Q. He D. Zhao J. Yan F. Pan Y. Prevalence of the optr A gene among Streptococcus suis isolates from diseased pigs and identification of a novel integrative conjugative element ICE Ssu 988S Res. Microbiol.202317410407810.1016/j.resmic.2023.10407837149078 · doi ↗ · pubmed ↗
- 6Gao Y. Xia L. Pan R. Xuan H. Guo H. Song Q. Wei J. Shao D. Liu K. Li Z. Identification of mcr-1 and a novel chloramphenicol resistance gene cat T on an integrative and conjugative element in an Actinobacillus strain of swine origin Vet. Microbiol.202125410898310.1016/j.vetmic.2021.10898333486327 · doi ↗ · pubmed ↗
- 7Kong L.H. Xiang R. Wang Y.L. Wu S.K. Lei C.W. Kang Z.Z. Chen Y.P. Ye X.L. Lai Y. Wang H.N. Integration of the bla NDM-1 carbapenemase gene into a novel SXT/R 391 integrative and conjugative element in Proteus vulgaris J. Antimicrob. Chemother.2020751439144210.1093/jac/dkaa 06832155266 · doi ↗ · pubmed ↗
- 8He J. Sun L. Zhang L. Leptihn S. Yu Y. Hua X. A Novel SXT/R 391 Integrative and Conjugative Element Carries Two Copies of the bla NDM-1 Gene in Proteus mirabilism Sphere 20216 e 005882110.1128/m Sphere.00588-2134378988 PMC 8386438 · doi ↗ · pubmed ↗