Hippo Signaling Dysregulation in Breast Cancer: Subtype-Independent Gene and miRNA Signatures
Katarzyna Król-Jatręga, Elżbieta Mitka-Krysiak, Kacper Boroń, Nikola Zmarzły, Piotr Ossowski, Aleksandra Plata-Babula, Paweł Ordon, Wojciech Kulej, Tomasz Sirek, Julia Gajdeczka, Yuriy Prudnikov, Krzysztof Bereza, Olga Nowotny-Czupryna, Dariusz Boroń, Beniamin Oskar Grabarek

TL;DR
This study identifies genes and miRNAs linked to Hippo signaling that are consistently dysregulated across all breast cancer subtypes, suggesting potential universal biomarkers and therapeutic targets.
Contribution
The study reveals subtype-independent Hippo-related gene and miRNA signatures that are consistently dysregulated in all breast cancer subtypes.
Findings
STK4, RASSF6, and FGF1 are consistently downregulated across all breast cancer subtypes.
BIRC5 and SERPINE1 are overexpressed, indicating activation of downstream oncogenic effectors.
miR-522-3p, miR-199b-5p, and miR-30a-3p are potential regulators of Hippo-related genes.
Abstract
Background/Objectives: Breast cancer represents a diverse group of malignancies and continues to rank among the leading causes of cancer-related deaths in women. Altered Hippo pathway signaling has been increasingly recognized as a contributor to tumor growth, therapeutic resistance, and metastatic spread. This study aimed to identify miRNAs targeting Hippo pathway-related genes that are consistently dysregulated across all five breast cancer subtypes. Methods: The study cohort included patients representing five breast cancer subtypes: 130 luminal A, 96 HER2-positive luminal B, 100 HER2-negative luminal B, 36 non-luminal HER2-positive, and 43 triple-negative breast cancer (TNBC). Tumor samples were collected during surgery, along with adjacent healthy tissue that served as controls. Expression of Hippo-related genes was analyzed using mRNA microarrays and validated with reverse…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
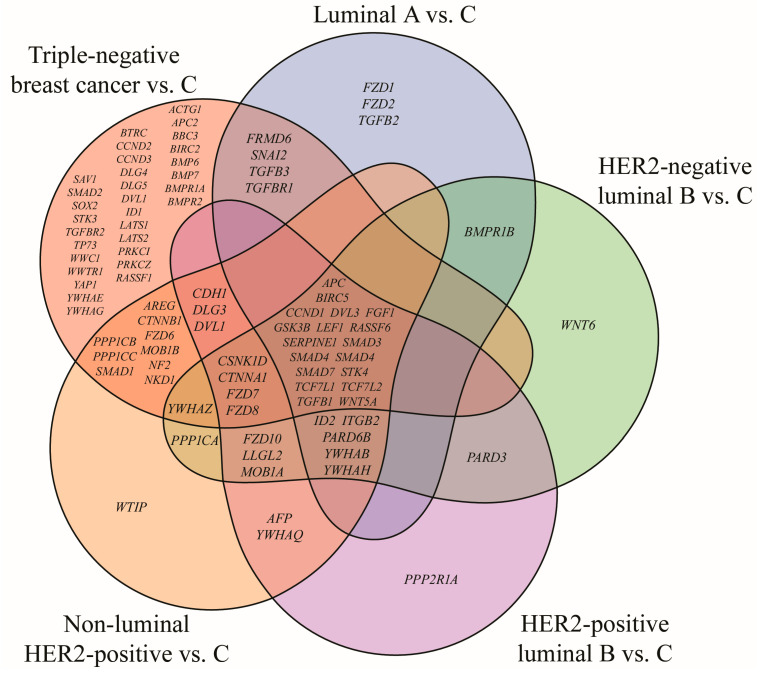 Figure 1
Figure 1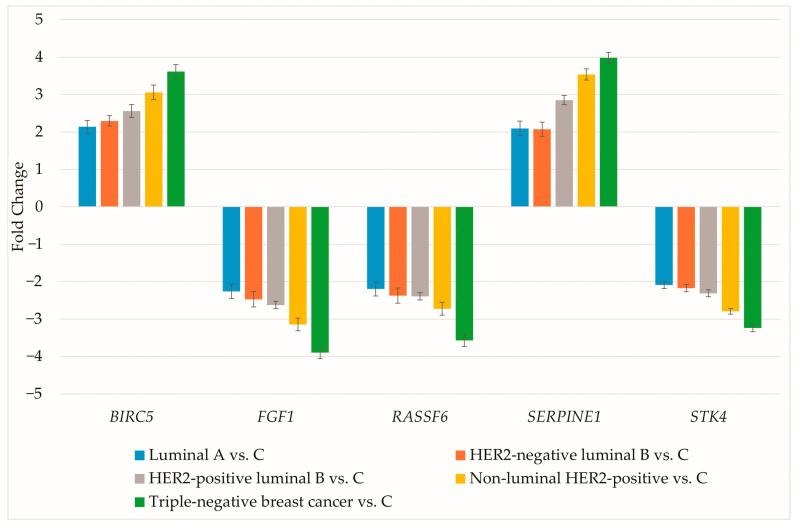 Figure 2
Figure 2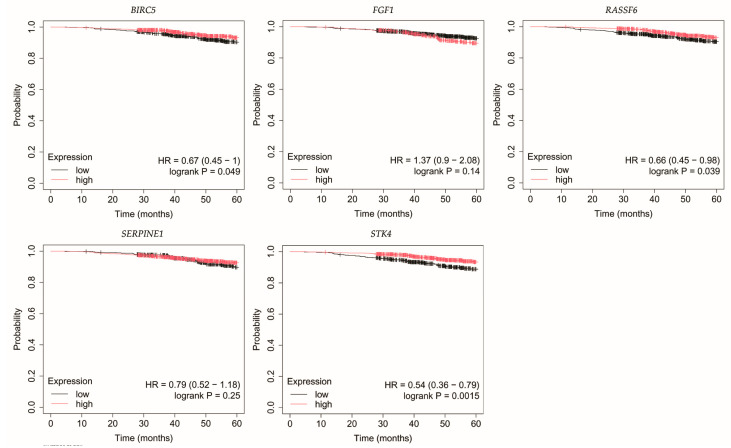 Figure 3
Figure 3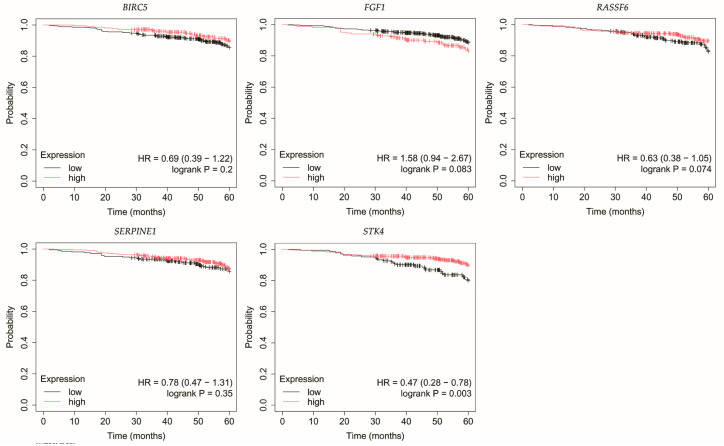 Figure 4
Figure 4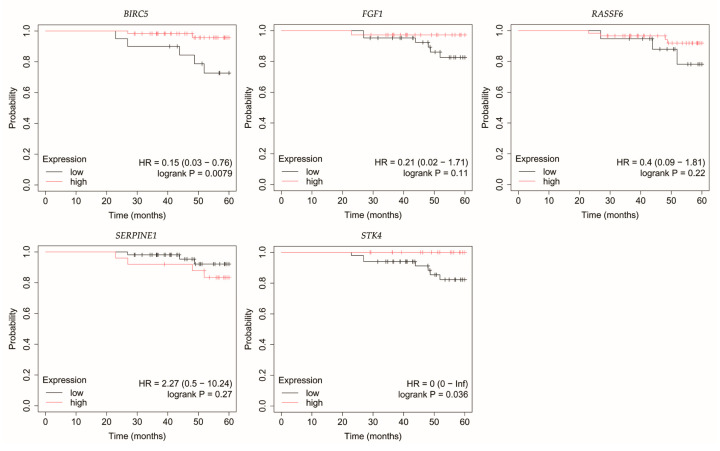 Figure 5
Figure 5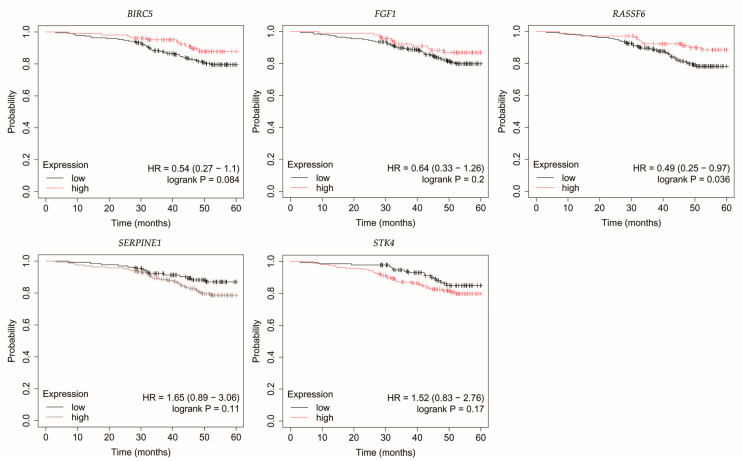 Figure 6
Figure 6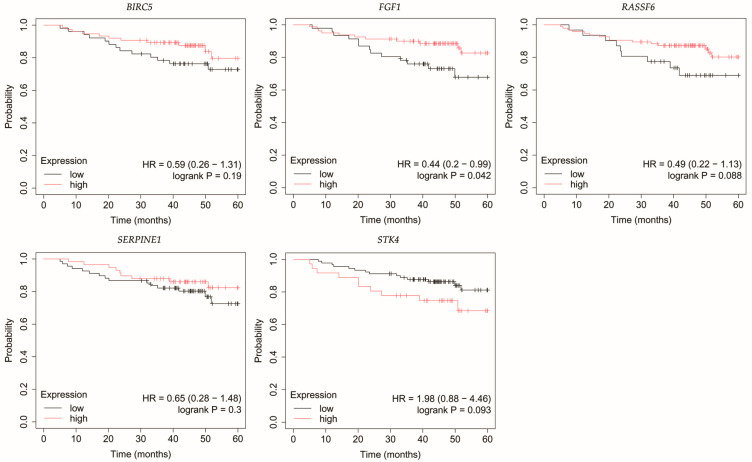 Figure 7
Figure 7Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHippo pathway signaling and YAP/TAZ · Wnt/β-catenin signaling in development and cancer
1. Introduction
Breast cancer continues to be the most common malignancy diagnosed in women and the foremost cause of cancer mortality worldwide [1]. It significantly impacts patients beyond survival by causing long-term morbidity, reducing quality of life through physical, psychological, and financial burdens and imposing substantial economic costs on healthcare systems and societies [2]. Breast cancer represents a biologically diverse disease composed of different molecular subtypes: luminal A, luminal B HER2-positive, luminal B HER2-negative, non-luminal HER2-positive, and triple-negative breast cancer (TNBC). Each subtype is defined by specific transcriptomic features, clinical behavior, and therapeutic approaches [3].
The Hippo signaling pathway is a fundamental regulator of tissue homeostasis, controlling cell proliferation, differentiation, and apoptosis [4]. Its core components include the serine/threonine-protein kinases 4/3 (STK4/3) and the adaptor Salvador (SAV1), which activate the large tumor suppressor kinases 1/2 (LATS1/2) through phosphorylation. In cooperation with MOB kinase activators 1A and 1B (MOB1A/B), activated LATS1/2 phosphorylates the transcriptional co-activators Yes-associated protein (YAP) and transcriptional coactivator with PDZ-binding motif (TAZ), leading to their cytoplasmic retention and degradation [5]. When Hippo signaling is functional, nuclear translocation of YAP/TAZ and activation of growth-promoting genes is suppressed [6]. In contrast, Hippo inactivation enables nuclear accumulation of YAP and TAZ, where they interact with transcriptional enhanced associate domain (TEAD) transcription factors to activate gene programs promoting cell proliferation, resistance to apoptosis, migration, and epithelial–mesenchymal transition (EMT) [7,8].
Altered Hippo signaling has therefore emerged as a hallmark of oncogenesis, contributing not only to tumor initiation but also to therapy resistance and metastatic progression [9]. The pathway is not limited to its canonical kinase cascade but also integrates multiple upstream signals, including extracellular matrix stiffness, G protein-coupled receptor signaling, and growth factor pathways [10,11]. Through this integration, Hippo interacts with several major oncogenic pathways, such as Wnt/β-catenin, TGF-β, PI3K/AKT, and MAPK [12,13,14,15]. This positioning highlights Hippo as both a driver of tumor cell-intrinsic behavior and a mediator of tumor-microenvironment interactions.
MicroRNAs (miRNAs) provide an additional layer of regulation by modulating gene expression at the post-transcriptional level. Dysregulated miRNA activity has been linked to breast cancer development, prognosis, and treatment resistance [16]. Yet, their contribution to Hippo signaling across breast cancer subtypes is not fully understood.
This study aimed to identify miRNAs that target Hippo pathway-related genes consistently dysregulated across all five breast cancer subtypes.
2. Materials and Methods
2.1. Patients
A total of 405 patients were included in the cohort. Obtained samples were classified as luminal A (130 cases), HER2-negative luminal B (100 cases), HER2-positive luminal B (96 cases), non-luminal HER2-positive (36 cases), and triple-negative breast cancer (TNBC; 43 cases). Control samples from adjacent non-cancerous tissue were also obtained intraoperatively. Every patient was staged as T1N0M0. Across all subtypes, the majority of patients were older than 50 years, with an average BMI above 30 (Table 1) [17].
The study protocol complied with the ethical guidelines of the Declaration of Helsinki and was approved by the Bioethical Committee of the Regional Medical Chamber in Krakow (81/KBL/OIL/2023; 10 March 2023). All participants provided written informed consent.
2.2. Isolation of Total RNA
Isolation of total RNA was performed using TRIzol Reagent (Invitrogen Life Technologies, Carlsbad, CA, USA; cat. no. 15596026). Subsequent purification was carried out with the RNeasy Mini Kit (QIAGEN, Hilden, Germany; cat. no. 74104), followed by DNase I treatment (Fermentas International Inc., Burlington, ON, Canada; cat. no. 18047019). RNA integrity and yield were verified by 1% agarose gel electrophoresis and spectrophotometry.
2.3. mRNA Profiling by Microarrays
HG-U133A 2.0 microarrays (Affymetrix, Santa Clara, CA, USA) in combination with the GeneChip™ 3′ IVT PLUS kit (Thermo Fisher Scientific, Waltham, MA, USA; cat. no. 902416) were employed for transcriptome profiling. Genes linked to the Hippo signaling pathway were retrieved from the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway database (hsa04390), yielding 157 genes mapped to 411 mRNA probes on the platform.
2.4. Gene Expression Analysis by Reverse Transcription Quantitative Polymerase Chain Reaction (RT-qPCR)
Validation of microarray data was carried out by RT-qPCR employing the SensiFast SYBR No-ROX One-Step Kit (Bioline, London, UK). Five candidate genes (BIRC5, FGF1, RASSF6, SERPINE1, STK4; Table 2) were analyzed. Relative expression was determined using the 2^−ΔΔCt^ method with β-actin (ACTB) as the reference gene.
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
Protein expression was assessed via ELISA with MyBioSource (San Diego, CA, USA) kits targeting BIRC5 (cat. no. MBS2887028), FGF1 (cat. no. MBS167170), RASSF6 (cat. no. MBS7612748), SERPINE1 (cat. no. MBS763621), STK4 (cat. no. MBS167492).
2.6. miRNA Profiling and Target Prediction
miRNA Microarray 2.0, in combination with the FlashTag Biotin HSR RNA Labeling Kit and the Hybridization Wash and Stain Kit (Affymetrix, Santa Clara, CA, USA), was utilized to detect miRNAs differentiating breast cancer from control tissue. The miRDB database (http://mirdb.org; accessed on 2 August 2025) was employed to predict regulatory links between miRNAs and the selected genes (BIRC5, FGF1, RASSF6, SERPINE1, STK4), considering only targets with a prediction score ≥ 80 [18].
2.7. Statistical Analysis
Transcriptome Analysis Console (Thermo Fisher Scientific, Waltham, MA, USA) was used for microarray data processing. One-way ANOVA and Tukey’s post hoc test were applied with thresholds set at p < 0.05 and FC > 2 or <−2. Gene lists for each breast cancer subtype and their intersections were generated using Python (ver. 3.13.7). Venn diagram was created in R (ver. 4.4.0; package Venn ver. 1.12). Final figures were assembled in Adobe Photoshop. For RT-qPCR and ELISA data, Statistica 13.3 (StatSoft, Krakow, Poland) was employed. Data distribution was verified with the Shapiro-Wilk test. The Kruskal-Wallis test and Dunn’s post hoc test were then carried out.
Overall survival (OS) across breast cancer subtypes was assessed using the Kaplan-Meier plotter (http://kmplot.com/, accessed on 28 June 2025), applying a 60-month follow-up cut-off [19,20].
Sample size estimation with G*Power 3.1.9.718 (f = 0.25, α = 0.05, power = 0.95) yielded n = 324, while the study’s 405 participants provided a post hoc power of 0.98 [21].
3. Results
3.1. mRNA Microarray-Based Gene Expression Profiling
Among the 418 mRNAs associated with the Hippo signaling pathway, 145 were found to be significantly dysregulated in breast cancer compared with control tissue. Subsequent Tukey’s post hoc testing demonstrated subtype-specific expression changes, with 45 transcripts altered in luminal A, 52 in HER2-negative luminal B, 58 in HER2-positive luminal B, 70 in non-luminal HER2-positive, and 117 in TNBC. The overlapping and unique genes among the subtypes are depicted in the Venn diagram (Figure 1).
The analysis identified subtype-specific genes, with TNBC displaying the largest number. In addition, 18 genes were common to all breast cancer cases irrespective of subtype. 13 of these have been reported in our previous studies, while the current study concentrated on five novel genes: BIRC5, FGF1, RASSF6, SERPINE1 and STK4 (Table 3).
Expression profiling indicated upregulation of BIRC5 and SERPINE1 independent of breast cancer subtype. In contrast, FGF1, RASSF6 and STK4 exhibited significant downregulation in this cohort.
3.2. RT-qPCR and ELISA Analysis of BIRC5, FGF1, RASSF6, SERPINE1 and STK4 Expression
The expression of genes differentiating breast cancer irrespective of subtype was evaluated using RT-qPCR. Results are presented as mean ± standard deviation (Figure 2).
RT-qPCR data aligned with the microarray findings. Protein concentrations for the selected genes were then determined (Table 4).
The results obtained from protein quantification were consistent with those observed for mRNA expression.
3.3. Prediction of miRNA Targets
The subsequent step involved evaluating whether BIRC5, FGF1, RASSF6, SERPINE1 and STK4 could serve as targets for miRNAs differentiating breast cancer from the control (Table 5).
BIRC5, FGF1 and RASSF6 are unlikely to be targets of miRNAs significantly altered across five breast cancer subtypes in this study. SERPINE1 overexpression may be associated with low levels of miR-199b-5p and miR-30a-3p. Moreover, decreased expression of STK4 may be the result of high miR-522-3p activity.
3.4. Overall Survival Outcomes Across Breast Cancer Subtypes
The outcomes of the overall survival (OS) analysis are presented in Figure 3, Figure 4, Figure 5, Figure 6 and Figure 7.
In luminal A breast cancer, reduced expression of RASSF6 and STK4 correlated with poorer OS (Figure 3). Similarly, for BIRC5, however, it should be noted that p was 0.049.
Loss of STK4 expression in HER2-negative luminal B tumors correlated with reduced OS (Figure 4).
In HER2-positive luminal B breast cancer, reduced STK4 expression was correlated with decreased OS (Figure 5).
In non-luminal HER2-positive breast cancer, reduced RASSF6 expression was linked with poorer OS (Figure 6).
Analysis of TNBC cases indicated that patients with low FGF1 expression exhibited reduced OS (Figure 7).
4. Discussion
Through an integrated evaluation of five molecular subtypes, we identified a core set of Hippo-related genes that are consistently altered across breast cancers. We observed upregulation of BIRC5, and SERPINE1, along with FGF1, RASSF6 and STK4 downregulation, which was further confirmed at the protein level. Furthermore, we identified miRNAs potentially targeting SERPINE1 and STK4.
STK4, also known as MST1, is a pivotal upstream kinase of the Hippo pathway responsible for phosphorylating and activating LATS1/2, thereby restricting YAP/TAZ nuclear translocation and oncogenic activity [22]. Its loss has been linked to tumor initiation and progression, suggesting a role as an early destabilizer of Hippo signaling [23]. Lin et al. confirmed the suppressive role of STK4 in breast cancer, emphasizing that its low levels were associated with reduced patient survival [24] and proposed its utility as a prognostic and predictive biomarker, as well as a therapeutic target [25]. Similarly, Jun et al. demonstrated that STK4 overexpression suppressed proliferation and migration of breast cancer cells, whereas the application of an STK4 inhibitor reversed these effects and inhibited the Hippo pathway [26]. Li et al. demonstrated that STK4 can undergo deacetylation, which leads to its degradation in lysosomes during autophagy. This resulted in decreased STK4 expression and promotion of breast cancer cell growth [27]. Our results align with these findings, as we observed consistent STK4 downregulation across all breast cancer subtypes. Bioinformatic predictions suggest that this effect may, at least in part, be mediated by miR-522-3p. Interestingly, this miRNA has been reported to exert both oncogenic and tumor-suppressive roles depending on the cancer context [28,29,30,31,32]. In breast cancer, bioinformatic analysis by Tan et al. revealed that miR-522 participates in the regulation of proliferation, migration, and EMT [33]. Dong et al. demonstrated that targeting p63 protein by this miRNA in breast epithelial cells resulted in the acquisition of mesenchymal morphology, which accelerated cell migration. Reversing this process may therefore constitute a potential anti-metastatic strategy [34]. In addition, Wang et al. showed that miR-522 can target BRCA1 in TNBC, where its overexpression predicted reduced overall patient survival and a higher incidence of lymph node metastasis [35]. Collectively, we have identified STK4 as a potential target of miR-522-3p regardless of breast cancer subtype. Importantly, low STK4 expression was linked to poorer overall survival in luminal A and luminal B subtypes, reinforcing its value as a prognostic biomarker and potential therapeutic target.
RASSF6 is recognized as a tumor suppressor that is downregulated in various cancers [36,37,38], including breast cancer [39]. Mechanistically, it interacts with MDM2, promoting its degradation and thereby stabilizing p53 [40]. Remarkably, even in the absence of p53, RASSF6 retains the ability to induce apoptosis and cell cycle arrest through the pRb-E2F1 axis [41]. Moreover, RASSF6 directly activates the Hippo pathway by enhancing the phosphorylation of STK4/3 and LATS1/2 kinases [39], thereby reinforcing its role as a critical upstream activator of Hippo tumor-suppressive signaling. In our study, RASSF6 was consistently downregulated across all breast cancer subtypes, indicating a subtype-independent mechanism for Hippo pathway impairment. Unlike STK4, this effect was not attributable to any of the miRNAs identified in our analysis. Clinically, low RASSF6 expression correlated with poorer overall survival in patients with luminal A and non-luminal HER2-positive cancers, underscoring its prognostic significance and potential utility as a biomarker of Hippo pathway integrity.
FGF1 can influence the Hippo pathway through interactions with receptor tyrosine kinases, notably FGFR1, which subsequently activate downstream MAPK and PI3K signaling cascades [42,43,44]. This signaling cross-talk represents a complex regulatory network in which FGF1 binding induces downstream changes that impact cell proliferation, survival, and differentiation, which are also modulated by the Hippo pathway [42,45]. The FGF1/FGFR axis has been linked to breast cell proliferation and migration, suggesting its potential as a therapeutic target [46]. Szymczak et al. confirmed the involvement of FGF1 in drug resistance in the MCF-7 breast cancer line [47]. Castillo-Castrejon et al. demonstrated that FGF1 can activate ER independently of estrogen in obese women, promoting breast cancer progression [48]. In our study, FGF1 expression was decreased across all subtypes, with low levels correlating with worse overall survival in TNBC. Notably, no miRNAs meeting our selection criteria were predicted to target FGF1, indicating that alternative regulatory mechanisms may be responsible. Overall, FGF1 downregulation may disrupt upstream extracellular signaling that modulates Hippo activity, further contributing to pathway suppression.
BIRC5, also known as survivin, belongs to the family of antiapoptotic proteins whose expression can be induced by YAP/TAZ, resulting in inhibition of apoptosis [49]. It promotes tumorigenesis through two principal mechanisms: preventing apoptosis via caspase inhibition, which allows cancer cells to survive, and facilitating mitotic progression, thereby supporting cancer cell survival and proliferation [50]. Overexpression of BIRC5 is frequently observed in various cancers and is linked to tumor initiation and progression [51], making it a valuable diagnostic and prognostic biomarker as well as a potential therapeutic target [52]. In breast cancer, high BIRC5 expression has been reported particularly in basal-like and luminal B subtypes, correlating with poorer patient survival independent of estrogen receptor status or lymph node involvement [53]. Targeting BIRC5 in combination with standard therapies has been suggested by Mehraj et al. as a strategy to improve treatment efficacy [54]. Hamilton et al. emphasized higher risk of breast cancer tumors with BIRC5 overexpression in Black and younger women [55]. Al-Yahya et al. demonstrated that post-transcriptional regulation by RNA-binding proteins, such as tristetraprolin, can reduce BIRC5 levels, inhibiting proliferation and enhancing patient outcomes [56]. Consistent with previous findings, our analysis revealed BIRC5 overexpression across all breast cancer subtypes, reflecting a functional consequence of Hippo pathway inactivation that promotes tumor cell survival. Importantly, BIRC5 expression in our cohort was not significantly influenced by miRNAs, suggesting that transcriptional activation (likely via YAP/TAZ–TEAD complexes) is the predominant mechanism. This consistent upregulation positions BIRC5 as a central Hippo effector and a promising therapeutic target, especially in contexts of apoptosis-resistant tumors.
SERPINE1, also known as PAI-1, is a serine protease inhibitor involved in extracellular matrix remodeling, angiogenesis, and metastatic progression [57]. It is a recognized transcriptional target of YAP/TAZ [58] and is frequently upregulated across multiple cancer types, where its overexpression correlates with tumor promotion and poor patient prognosis [57,59,60,61]. In breast cancer specifically, elevated SERPINE1 levels have been associated with worse overall and relapse-free survival [62]. Zhang et al. demonstrated a link between high SERPINE1 levels and paclitaxel resistance in TNBC [63], whereas Su et al. reported similar observation for obesity-associated tumor radioresistance [64]. In our study, SERPINE1 was consistently overexpressed across all breast cancer subtypes, underscoring its role as both a Hippo pathway effector and a contributor to tumor progression. Notably, increased SERPINE1 expression correlated with decreased levels of miR-199b-5p and miR-30a-3p, both of which are established tumor suppressor miRNAs in breast cancer [65,66,67,68,69,70,71,72,73]. This suggests that SERPINE1 dysregulation may be reinforced at multiple regulatory levels, and our findings indicate it as a novel target of miR-199b-5p and miR-30a-3p, highlighting potential avenues for therapeutic intervention.
Although the identified genes did not display pronounced heterogeneity in expression or prognosis between different breast cancer subtypes, this lack of variation highlights their potential as ‘subtype-independent’ biomarkers. Rather than distinguishing between subtypes, these genes may reflect a core dysregulation of the Hippo pathway across breast cancers. Such markers may complement existing subtype-specific classifiers by offering universal prognostic and therapeutic insights, applicable irrespective of molecular subtype.
Our study has several limitations. First, the cohort consisted exclusively of Polish women with T1N0M0 tumors, which may restrict generalizability to other populations. Second, the numbers of patients in the non-luminal HER2-positive and TNBC subgroups were relatively small compared with luminal subtypes, which may affect the robustness of subgroup-specific findings. Third, protein expression was validated using ELISA rather than Western blot; while ELISA provides quantitative data, it does not allow visualization of full-length protein bands. Fourth, functional validation of the predicted miRNA–mRNA interactions (e.g., luciferase reporter assays, knockdown/overexpression experiments) was not performed, and future studies are required to confirm these regulatory relationships.
5. Conclusions
This study reveals a conserved pattern of Hippo pathway dysregulation across all major breast cancer subtypes. We demonstrated that STK4, RASSF6, and FGF1 were consistently downregulated, whereas BIRC5, and SERPINE1 were upregulated. These changes reflect both impaired upstream Hippo activation and enhanced downstream oncogenic signaling. Notably, STK4 and SERPINE1 appear to be regulated by miRNAs, including miR-522-3p, miR-199b-5p, and miR-30a-3p. By focusing on subtype-independent events, our findings highlight the Hippo pathway as a common axis of dysregulation in breast cancer. This suggests that Hippo-related genes and their miRNA regulators could serve as universal biomarkers or therapeutic targets, complementing subtype-specific treatment approaches.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bray F. Laversanne M. Sung H. Ferlay J. Siegel R.L. Soerjomataram I. Jemal A. Global Cancer Statistics 2022: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries CA Cancer J. Clin.20247422926310.3322/caac.2183438572751 · doi ↗ · pubmed ↗
- 2Gąska I. Czerw A. Pajewska M. Partyka O. Deptała A. Badowska-Kozakiewicz A. Budzik M. Sygit K. Wojtyła-Buciora P. Drobnik J. The Cost of Breast Cancer: Economic and Social Perspective Cancers 202517301210.3390/cancers 1718301241008856 PMC 12468891 · doi ↗ · pubmed ↗
- 3Roy M. Fowler A.M. Ulaner G.A. Mahajan A. Molecular Classification of Breast Cancer PET Clin.20231844145810.1016/j.cpet.2023.04.00237268505 · doi ↗ · pubmed ↗
- 4Fu M. Hu Y. Lan T. Guan K.-L. Luo T. Luo M. The Hippo Signalling Pathway and Its Implications in Human Health and Diseases Signal Transduct. Target. Ther.2022737610.1038/s 41392-022-01191-936347846 PMC 9643504 · doi ↗ · pubmed ↗
- 5Ma S. Meng Z. Chen R. Guan K.-L. The Hippo Pathway: Biology and Pathophysiology Annu. Rev. Biochem.20198857760410.1146/annurev-biochem-013118-11182930566373 · doi ↗ · pubmed ↗
- 6Sebio A. Lenz H.-J. Molecular Pathways: Hippo Signaling, a Critical Tumor Suppressor Clin. Cancer Res.2015215002500710.1158/1078-0432.CCR-15-041126384319 PMC 4644679 · doi ↗ · pubmed ↗
- 7Ghaboura N. Unraveling the Hippo Pathway: YAP/TAZ as Central Players in Cancer Metastasis and Drug Resistance EXCLI J.20252461263710.17179/excli 2025-835140630468 PMC 12235270 · doi ↗ · pubmed ↗
- 8Misra J.R. Irvine K.D. The Hippo Signaling Network and Its Biological Functions Annu. Rev. Genet.201852658710.1146/annurev-genet-120417-03162130183404 PMC 6322405 · doi ↗ · pubmed ↗