Cultivation of Arthrospira platensis in Veterinary Hospital Wastewater Enhances Pigment Production and Reduces Antibiotic Resistance Genes
Authen Promariya, Sekbunkorn Treenarat, Nattaphong Akrimajirachoote, Wanat Sricharern, Wuttinun Raksajit

TL;DR
Arthrospira platensis can treat veterinary hospital wastewater by boosting pigment production and reducing antibiotic resistance genes.
Contribution
This study demonstrates the dual use of Arthrospira platensis for wastewater treatment and pigment production in veterinary hospital wastewater.
Findings
A. platensis cultivated in 25% VHW showed highest biomass and pigment production.
Cultivation reduced Proteobacteria dominance and key antibiotic resistance genes in wastewater.
The study shows A. platensis can simultaneously treat wastewater and produce valuable pigments.
Abstract
Veterinary hospital wastewater (VHW) contains high levels of nutrients, organic pollutants, and antibiotic resistance genes (ARGs), posing environmental risks. This study investigated the potential of Arthrospira platensis for treating VHW. A. platensis was cultivated in different wastewater concentrations for 8 days. Growth and pigment production were highest in 25% VHW, while higher concentrations inhibited performance. In 25% VHW, pigment levels increased significantly compared to the control. Metagenomic analysis showed that cultivation of A. platensis in wastewater altered the microbial community and reduced both harmful bacteria and ARGs. The dominant bacteria group, Proteobacteria, decreased markedly after A. platensis cultivation in 25% VHW, which was also accompanied by a reduction in key resistance genes. These findings demonstrate that A. platensis can simultaneously treat…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
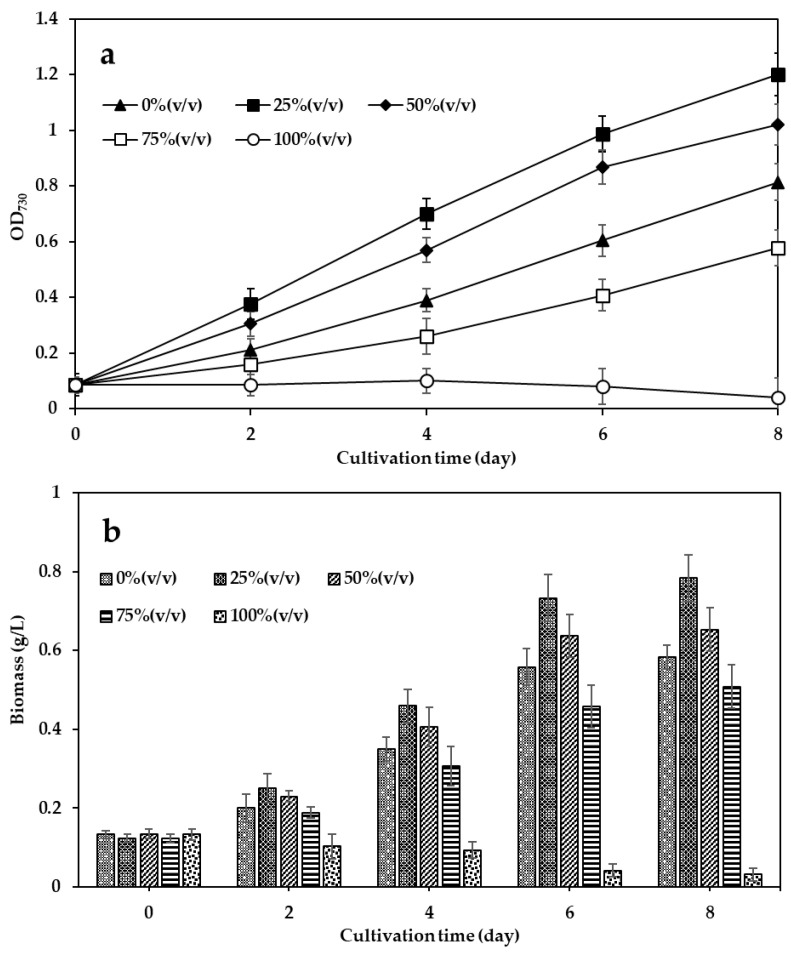 Figure 1
Figure 1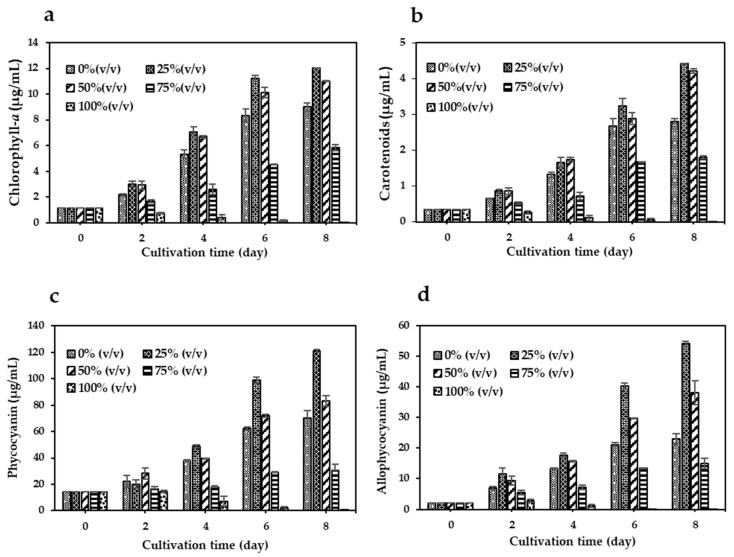 Figure 2
Figure 2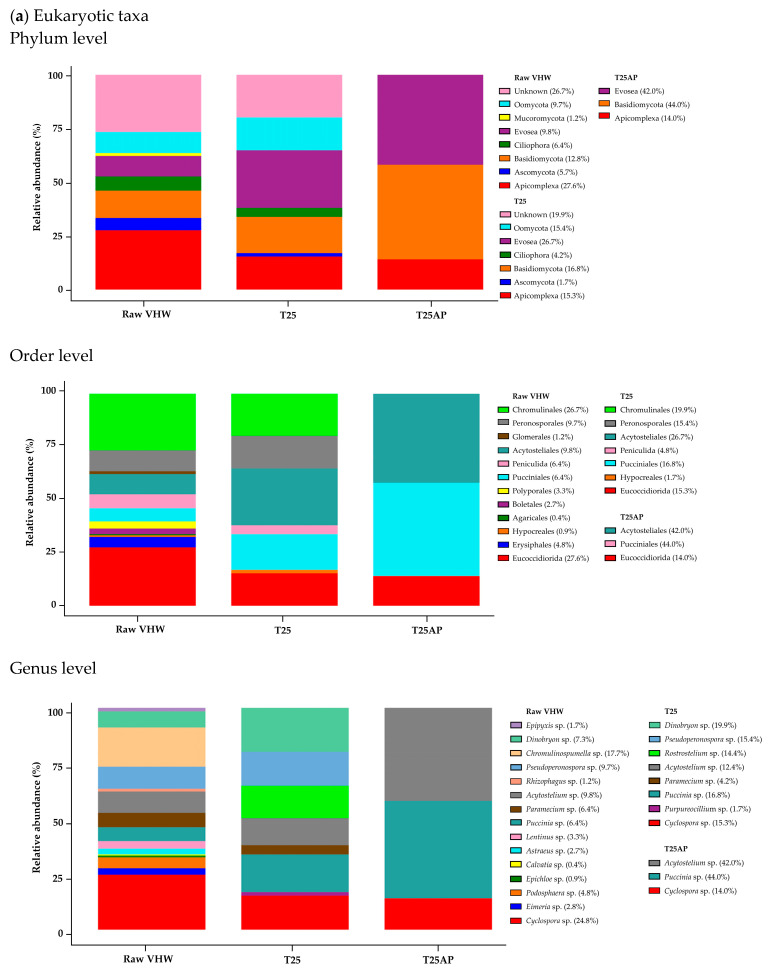 Figure 3
Figure 3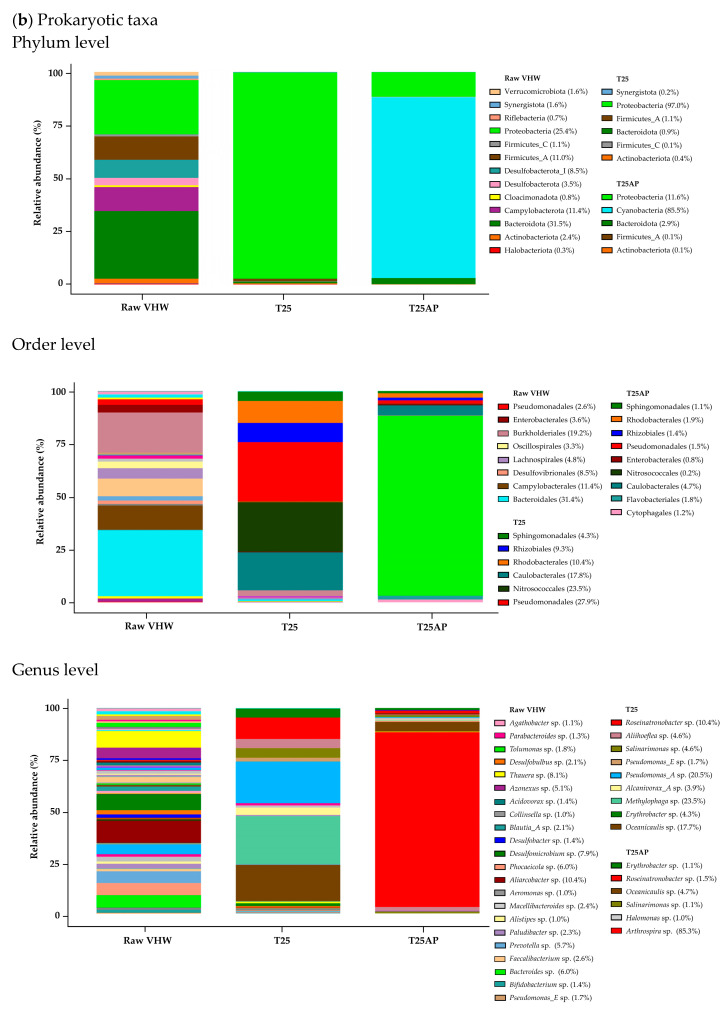 Figure 4
Figure 4 Figure 5
Figure 5- —Kasetsart University
- —National Science and Technology Development Agency (NSTDA)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAquaculture disease management and microbiota · Pharmaceutical and Antibiotic Environmental Impacts · Microbial Community Ecology and Physiology
1. Introduction
Veterinary hospitals are major sources of antibiotic contamination in wastewater, as broad-spectrum antibiotics are frequently prescribed for companion animals. A wide range of antibiotics has been detected in veterinary hospital wastewater (VHW), with quinolones being the most prevalent group, consistent with their extensive clinical use, followed by β-lactams, tetracyclines, metronidazoles, macrolides, trimethoprim, and sulfonamides, at concentrations ranging from approximately 31 to 15,701 ng/L [1]. This distribution pattern is comparable to reports from human hospital effluents, reflecting the overlap in antibiotic use between human and veterinary medicine [2,3,4]. Despite adherence to antimicrobial stewardship guidelines, drug residues are frequently present in veterinary clinic wastewater, posing environmental risks. These residues may persist through treatment processes and contribute to the development of resistant microorganisms [5,6]. Such bacteria, often carrying transferable resistance genes, can spread through water, waste streams, and areas contaminated by pets. Multidrug-resistant infections place a burden on veterinary care and threaten public health. Addressing this challenge effectively requires the implementation of on-site treatment solutions specifically designed for veterinary wastewater. Conventional wastewater treatment is generally ineffective at removing antibiotics due to their limited treatment capacity [7,8], while alternative methods such as physical separation and chemical degradation are often restricted by high operational costs and economic feasibility [9]. Consequently, there is growing interest in sustainable technologies capable of efficiently removing these contaminants to mitigate environmental and public health risks [10,11,12]. Recent research has focused on developing systems to reduce or eliminate antibiotics prior to discharge or reuse, with increasing attention being directed toward biological alternatives. In particular, microalgae-based systems have emerged as a promising, cost-effective, and environmentally friendly approach for removing various pharmaceuticals and personal care products [13,14].
Microalgal-bacterial consortia (MBC) systems exploit interactions between microalgae and bacteria to simultaneously remove organic matter, excess nutrients, and contaminants [15,16]. These interactions are diverse and complex, ranging from mutualistic symbiosis to competitive antagonism, and may also include commensalism and amensalism [17]. In MBC systems, antagonistic effects can arise when decaying microalgae consume oxygen or release inhibitory compounds, limiting oxygen availability for bacteria. Additionally, under mixotrophic conditions, microalgal consumption of organic matter can further reduce net oxygen production, thereby restricting bacterial activity. Conversely, under a symbiotic relationship, microalgae supply oxygen through photosynthesis to support bacterial nutrient utilization, while bacteria release carbon dioxide that microalgae use. Effective system performance depends on balancing these competing and cooperative dynamics through careful control of light, nutrient levels, and aeration [18,19,20]. Arthrospira sp. (commonly known as Spirulina) is particularly promising and has demonstrated strong potential for cultivation in various wastewater sources, including municipal [21], dairy farm [22], and swine wastewater [23]. It can significantly enhance treatment efficiency by utilizing carbon sources in wastewater to support its growth and biomass yield while simultaneously producing valuable bioproducts such as chlorophyll-a, phycocyanin, and carotenoids. Although previous studies have demonstrated the benefits of A. platensis in wastewater utilization, research on A. platensis–bacterial interactions during wastewater treatment, particularly regarding the removal of antibiotic-resistant bacteria and resistance genes, remains limited. Therefore, this study aimed to evaluate the potential of A. platensis to produce biomass and pigments in VHW while also assessing its ability to reduce antibiotic-resistant bacteria and resistance genes.
2. Materials and Methods
2.1. Wastewater Sampling and Quality Analysis
Wastewater samples were randomly collected from a veterinary hospital located at a university in Bangkok, Thailand, in July 2023. Samples were collected in glass bottles with screw caps and transported to the laboratory. Composite sampling was performed by collecting water at hourly intervals over a 3-h period, with each sample taken from the water surface at a depth of approximately 1–2 feet. Sampling points included the center, left edge, and right edge of the wastewater retention pond. Three subsamples of 500 mL each were collected and then combined to create a representative composite sample. The composite sample was filtered through 0.45 μm membrane filters to remove suspended solids and stored at −20 °C until analysis. Physicochemical parameters, including ammonia nitrogen (NH_3_–N), nitrile nitrogen, nitrate nitrogen (NO_3_^−^–N), total phosphate (PO_4_^3−^), biochemical oxygen demand (BOD), chemical oxygen demand (COD), total dissolved solids (TDS), total suspended solids (TSS), total Kjeldahl nitrogen (TKN), fats, oils, and grease (FOG), and pH, were analyzed according to the standard methods for the examination of water and wastewater (APHA/AWWA/WEF) by Central Laboratory (Thailand) Company Limited (Bangkok, Thailand) [24,25].
2.2. Microorganism and Culture Conditions
An inoculum was prepared by culturing the Arthrospira platensis IFRPD1182, obtained from the Institute of Food Research and Product Development, Kasetsart University, Bangkok, Thailand [26], in Zarrouk medium mixed with non-sterile VHW at 0% VHW (0 mL VHW + 100 mL medium), 25%VHW (25 mL VHW + 75 mL medium), 50%VHW (50 mL VHW + 50 mL medium), 75%VHW (75 mL VHW + 25 mL medium), and 100% (100 mL VHW + 0 mL medium). The Zarrouk medium contained (g/L): NaHCO_3_ 16.8, NaNO_3_ 2.5, K_2_SO_4_ 1.0, NaCl 1.0, K_2_HPO_4_ 0.5, CaCl_2_·2H_2_O 0.04, Na_2_EDTA·2H_2_O 0.03, MgSO_4_·7H_2_O 0.2, FeSO_4_·7H_2_O 0.01, supplemented with 1.0 mL/L of trace elements solution containing (g/L): H_3_BO_3_ 2.86, MoO_3_ 0.02, MnCl_2_·4H_2_O 1.81, CuSO_4_·5H_2_O 0.08, ZnSO_4_·7H_2_O 0.22, Co(NO_3_)2·6H_2_O 0.05). The medium was initially adjusted to pH 10 ± 0.5 using 5 N NaOH. Cultures were incubated for 8 days on a rotary shaker at 120 rpm, under continuous LED illumination of 40 µmol/m^2^/s, at 30 ± 2 °C.
2.3. Biomass and Pigment Production
Biomass concentration was measured spectrophotometrically (Thermo Fisher Scientific, Waltham, MA, USA), from the optical density of the culture, according to Duangsri [26]. The regression Equation (1) is
where X (g/L) is the biomass concentration and OD_730_ is the absorbance of the suspension at 730 nm, a wavelength chosen to minimize interference from pigments [27].
Chlorophyll-a (Chl-a) and carotenoids (Car) were quantified using the absorption coefficient (E_λ_) [28], following Equations (2) and (3) [29], respectively. In brief, a 1 mL culture sample was centrifuged at 8000× g for 7 min in a high-speed refrigerated microcentrifuge (TOMY, Tokyo, Japan), and the pellet was subsequently extracted with 99.9% methanol in the dark for 20 min at 4 °C. The absorbance of the extracts was measured at 665 nm (λ_max_ for Chl-a), 470 nm (λ_max_ for Car), and 720 nm using a Genesys 30 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA).
In addition, phycocyanin (PC) and allophycocyanin (APC) were quantified using Equations (4) [30] and (5) [31]. In brief, cells were suspended in phosphate buffer (pH 7), frozen in liquid nitrogen, stored at –20 °C, and disrupted using an ultrasonic homogenizer (Thermo Fisher Scientific, Waltham, MA, USA). The absorbance of extracts was measured at 620 nm (λ_max_ for PC) and 652 nm (λ_max_ for APC) with a Genesys 30 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA).
2.4. Shotgun Metagenomic Sequencing and Data Processing
Samples were processed using the ZymoBIOMICS^®^ shotgun metagenomic sequencing service (Zymo Research, Irvine, CA, USA). In brief, wastewater samples were filtered through a 0.2 μm membrane, and DNA was extracted using the ZymoBIOMICS^®^-96 MagBead DNA Kit (Zymo Research Corporation, Irvine, CA, USA). Sequencing libraries were prepared using either the KAPA™ Hyper-Plus Library Preparation Kit (Kapa Biosystems, Wilmington, MA, USA) or the Nextera^®^ DNA Flex Library Prep Kit (Illumina, San Diego, CA, USA), with up to 100 ng of input DNA and internal single-index 8 bp barcodes. Library quantification was performed using TapeStation^®^ (Agilent Technologies, Santa Clara, CA, USA) and quantitative PCR (qPCR), followed by sequencing on a HiSeq^®^ or NovaSeq^®^ platform (Illumina, San Diego, CA, USA). Raw sequencing reads were quality-trimmed using Trimmomatic-0.33 [32]. Taxonomic identification of bacterial and archaeal sequences was conducted using the complete GTDB database (R07-RS207) and pre-formatted GenBank databases (v.2022.03) via Sourmash.
2.5. Statistical Analysis
All data obtained in this study represent the means of three independent biological replicates, and the error bars represent the standard deviation (Mean ± SD, n = 3). The statistical analysis was analyzed by one way analysis of variance (ANOVA) and the significant difference (p < 0.05) were compared by Tukey’s HSD using SPSS version 22 (IBM, New York, NY, USA).
3. Results
3.1. Water Quality of VHW
Wastewater samples collected from the veterinary hospital were analyzed for key physicochemical parameters, as summarized in Table 1. The results indicated considerable contamination, particularly with nitrogenous compounds, organic pollutants, and solids.
3.2. Effect of VHW Proportions on Biomass and Pigment Production of A. platensis
As shown in Figure 1a, A. platensis cultured in 25% and 50% VHW exhibited growth patterns similar to 0% VHW but with notably higher OD_730_ values over the 8-day cultivation period. On day 8, biomass concentrations were 0.58 ± 0.04 g/L (0% VHW, control), 0.78 ± 0.05 g/L (25% VHW), 0.65 ± 0.05 g/L (50% VHW), and 0.50 ± 0.05 g/L (75% VHW). Biomass in the 25% VHW treatment was significantly higher than in the control (0% VHW) (p < 0.05), while the 50% and 75% VHW treatments did not differ significantly from the control. By contrast, the 100% VHW condition strongly inhibited growth, with marked suppression observed during the first two days and complete cessation by day 4 (Figure 1b).
Figure 2a–d show pigment production levels of chlorophyll-a (Chl-a), carotenoids (Car), phycocyanin (PC), and allophycocyanin (APC) in A. platensis cultures. All pigment levels in cultures with 25% VHW peaked on day 8, showing significant increases (p < 0.05) compared to 0% VHW: 1.3-fold for Chl-a (12.0 μg/mL), 1.5-fold for Car (4.4 μg/mL), 1.7-fold for PC (120 μg/mL), and 2.3-fold for APC (54 μg/mL). The 50% VHW also led to higher pigment levels than the control, though lower than those observed at 25% VHW. By contrast, cultures grown in 75% and 100% VHW exhibited reduced pigment content, with 100% VHW showing negligible pigment production.
3.3. Metagenomic Analysis of Wastewater Samples from a Veterinary Hospital
Shotgun metagenomic sequencing was employed to investigate microbial communities in VHW. Three sample types were examined: 100% VHW (raw VHW), 25% VHW cultured without A. platensis (T25), and 25% VHW cultured with A. platensis (T25AP). Taxonomic composition at the phylum, order, and genus levels was analyzed for both eukaryotic and prokaryotic communities. In raw VHW, the dominant eukaryotic phyla (≥10% relative abundance) were Basidiomycota (12.8%) and Apicomplexa (27.6%) (Figure 3a). Phyla with lower relative abundance included Evosea (9.8%), Oomycota (9.7%), Ascomycota (5.7%), and Ciliophora (6.4%). Under the T25 condition, the dominant eukaryotic phyla were Evosea (26.7%), Basidiomycota (16.8%), Oomycota (15.4%), and Apicomplexa (15.3%). By contrast, under the T25AP condition, Oomycota was no longer present, while Evosea (42.0%), Basidiomycota (44.0%), and Apicomplexa (14.0%) increased in relative abundance.
At the order level, dominant eukaryotic taxa (≥10% relative abundance) in raw VHW were Eucoccidiorida (27.6%) and Chromulinales (26.7%) (Figure 3a). Orders with lower relative abundance were Pucciniales (6.4%), Periculida (6.4%), Acytosteliales (9.8%), Peronosporales (9.7%), Erysiphales (4.8%), Boletales (2.7%), and Polyporales (3.3%). Under the T25 condition, dominant orders included Acytosteliales (26.7%), Chromulinales (19.9%), Pucciniales (16.8%), Peronosporales (15.4%), and Eucoccidiorida (15.3%), while Boletales and Polyporales were no longer detected. Following T25AP cultivation, Peronosporales and Chromulinales disappeared, while Acytosteliales (42.0%), Pucciniales (44.0%), and Eucoccidiorida (14.0%) increased in relative abundance.
At the genus level, dominant eukaryotic genera (≥10% relative abundance) in raw VHW included Cyclospora sp. (24.8%) and Chromulinospumella sp. (17.7%). Genera with lower abundance were Acytostelium sp. (9.8%), Pseudoperonospora sp. (9.7%), Puccinia sp. (6.4%), and Paramecium sp. (6.4%) (Figure 3a). Under the T25 condition, the dominant genera shifted to Dinobryon sp. (19.9%), Puccinia sp. (16.8%), Pseudoperonospora sp. (15.4%), Cyclospora sp. (15.3%), Rostrostelium sp. (14.4%), and Acytostelium sp. (12.4%). By contrast, under the T25AP condition, Dinobryon sp. and Pseudoperonospora sp. disappeared, while Cyclospora sp. (14.0%), Puccinia sp. (44.0%), and Acytostelium sp. (42.0%) showed pronounced increases in relative abundance.
In addition, in raw VHW, the dominant prokaryotic phyla (≥10% relative abundance) were Bacteroidota (31.5%), Proteobacteria (25.4%), Campylobacterota (11.4%), and Firmicutes_A (11.0%). Phyla with lower relative abundance included Desulfobacterota_I (8.5%), Desulfobacterota (3.5%), and Actinobacteriota (2.4%) (Figure 3b). Under the T25 condition, the dominant prokaryotic phylum was Proteobacteria (97.0%). By contrast, under the T25AP condition, Proteobacteria (11.6%), Bacteroidota (2.9%), and Firmicutes_A (0.1%) decreased markedly in relative abundance.
At the order level, dominant prokaryotic taxa (≥10% relative abundance) in raw VHW were Bacteroidales (31.4%), Burkhoderiales (19.2%), and Campylobacterales (11.4%) (Figure 3b). Orders with lower relative abundance were Desulfovibrionales (8.5%), Lachnospirales (4.8%), Eneterobacterales (3.6%), Oscillospirales (3.3%), and Pseudomonadales (2.6%). Under the T25 condition, dominant orders included Pseudomonadales (27.9%), Nitrosococcales (23.5%), Caulobacterales (17.8%), and Rhodobacterales (10.4%), while Rhizobiales (9.3%) and Sphingomonadales (4.3%) were detected at lower relative abundance. Following T25AP cultivation, all these orders decreased in relative abundance.
At the genus level, dominant prokaryotic genera (≥10% relative abundance) in raw VHW included Aliarcobacter sp. (10.4%). Genera with lower abundance were Bacteroides sp. (6.0%), Phocaeicola sp. (6.0%), Prevotella sp. (5.7%), Thauera sp. (8.1%), Azonexus sp. (5.1%), and Desulfomicrobium sp. (7.9%) (Figure 3b). Under the T25 condition, the dominant shifted to Methylophaga sp. (23.5%), Pseudomonas_A sp. (16.8%), Oceanicaulis sp. (17.7%), and Roseinatronobacter sp. (10.4%), while Thauera sp., Bacteroides sp., Desulfomicrobium sp., Azonexus sp., Phocaeicola sp., and Prevotella sp. disappeared. Following T25AP cultivation, Methylophaga sp. and Pseudomonas_A sp. were no longer present, while Oceanicaulis sp. (4.7%) and Roseinatronobacter sp. (1.5%) decreased in relative abundance.
3.4. Distribution of Antibiotic Resistance Genes (ARGs) Across Bacterial Genera
The heatmap in Figure 4 illustrates the distribution of ARGs across bacterial genera under two cultivation conditions: 25% VHW without A. platensis (T25) and 25% VHW with A. platensis (T25AP). The x-axis represents bacterial genera, each with columns for T25 and T25AP, while the y-axis lists 20 ARGs, including aph, oqxB, ant, aac, msr, mph, qac, vat, tet, erm, mef, dfr, sul, cat, lnu, and several beta-lactamase genes such as blaOXA, blaPBR, and blaNPS-1. Notably, Acinetobacter sp. under the T25 condition exhibited high expression of aph and moderate levels of oqxB and bla_OXA_, while these genes were less pronounced under T25AP. Methylophaga sp. displayed the strongest signal for oqxB under T25, suggesting a high prevalence of resistance genes. Pseudomonas_E sp. showed elevated oqxB under T25 and some mph expression under T25AP. Other genera, such as Halomonas sp., Phoceicola sp., and Thauera sp., demonstrated moderate levels of mph, erm, and tet, with slightly increased abundance under T25AP in some cases. By contrast, Bifidobacterium sp., Blautia_A sp., and Gemmiger sp. showed low or minimal ARG presence, indicating a limited role in resistance dissemination. Interestingly, Tolumonas sp. exhibited moderate aph abundance under both conditions. Overall, the heatmap reveals genus-specific ARG distribution patterns and highlights shifts in gene abundance under A. platensis treatment.
4. Discussion
This study investigated the potential of A. platensis as an alternative approach for removing ARGs while simultaneously producing valuable pigments from VHW. Antibiotics pose significant risks to both environmental and human health, even at low concentrations, and their overuse and misuse have contributed to the emergence of antibiotic-resistant bacteria, reducing the effectiveness of commonly prescribed antimicrobial therapies. Previous reports have demonstrated the occurrence of various classes of antibiotics in VHW, including quinolones, penicillin, tetracyclines, metronidazoles, macrolides, trimethoprim, and sulfonamides [1,7]. In the present study, however, antibiotic concentrations in VHW were not directly measured. Nevertheless, the antibiotics present are likely to include common veterinary drugs given their widespread therapeutic use in animals. The physicochemical profile of the VHW provides essential context for understanding the observed effects on A. platensis growth and pigment production (Table 1). Elevated levels of NH_3_–N (56.56 ± 3.23 mg/L) and TKN (76.13 ± 3.10 mg/L) indicate a nitrogen-rich environment, favorable for A. platensis cultivation, since nitrogen is required for protein synthesis and the biosynthesis of pigments such as phycobiliproteins and chlorophylls [33,34]. The low concentration of nitrate and undetectable nitrite suggest limited nitrification, possibly resulting from the anaerobic nature of the wastewater or the absence of active nitrifying bacteria at the time of sampling. At 25% VHW, the nitrogen load appears optimal, supporting biomass accumulation and pigment synthesis. This dilution likely provides sufficient nitrogen while minimizing toxicity. Conversely, inhibition observed at 75% and 100% VHW is likely due to excessive free ammonia, which at high concentrations disrupts cellular pH homeostasis and enzyme activity, impairing metabolic functions [35].
The high concentrations of organic pollutants, reflected by elevated BOD (71.50 ± 12.31 mg/L) and COD (160.97 ± 24.44 mg/L), indicate a considerable organic load that may indirectly affect microalgal growth. While A. platensis is not primarily heterotrophic, the presence of organic matter may promote the growth of competing heterotrophic bacteria in non-sterile cultures, resulting in nutrient competition or production of inhibitory metabolites. At high wastewater concentrations, such microbial competition and oxygen depletion could impair A. platensis viability. Additionally, the PO_4_^3−^ concentration (6.31 ± 1.43 mg/L) supports pigment biosynthesis and growth under light-sufficient conditions, as phosphorus is vital for ATP production and nucleic acid synthesis [36]. Excessive phosphorus in undiluted wastewater, however, may disrupt nutrient ratios (e.g., N:P), exacerbating stress responses [37]. The presence of FOG (9.59 ± 2.58 mg/L), together with moderate TDS (412.00 ± 21.57 mg/L) and TSS (25.06 ± 1.83 mg/L), may further explain the inhibition observed under concentrated VHW conditions. Lipophilic substances may coat microalgal cells, hindering nutrient uptake and light absorption, while suspended solids reduce light penetration, promoting shading effects in the culture medium.
This study demonstrates that diluted VHW can serve as a viable supplement to conventional media for cultivating A. platensis, enhancing both biomass and pigment production at optimal concentrations [38]. The 25% VHW treatment produced the highest biomass (0.78 ± 0.05 g/L) (Figure 1). These results suggest that moderate dilution provides adequate nitrogen and phosphorus for growth while minimizing toxicity from raw wastewater. Biomass at 50% VHW (0.65 ± 0.05 g/L) was slightly lower but still exceeded the control, indicating the adaptability of A. platensis to moderate wastewater concentrations. By contrast, growth in 100% VHW ceased by day 4, likely due to high organic loads or residual antibiotics impairing photosynthesis, enzymatic activity, and metabolism [37]. These findings are consistent with earlier studies reporting that high wastewater concentrations induce oxidative or chemical stress in microalgae, inhibiting growth [39].
Pigment production by A. platensis in response to varying VHW concentrations highlights its ability to utilize nutrients in diluted wastewater for metabolism and biosynthesis. At 25% VHW, pigment levels of chlorophyll-a, carotenoids, phycocyanin, and allophycocyanin increased significantly (Figure 2), indicating that moderate dilution supplies nutrients without triggering stress. On day 8, pigment levels peaked, with phycocyanin and allophycocyanin showing 1.7- and 2.3-fold increases, respectively, over the control, likely due to sufficient nitrogen and phosphorus availability. These results align with previous studies demonstrating enhanced pigment synthesis under low to moderate nutrient enrichment [40]. Higher VHW concentrations (75% and 100%) suppressed pigment production, with 100% VHW yielding negligible levels, likely due to toxic effects from residual pollutants [41]. Overall, the results emphasize the importance of partial wastewater dilution in facilitating nutrient uptake by A. platensis.
Shotgun metagenomic analysis revealed significant shifts in microbial community composition in VHW [42]. In raw VHW, the dominant eukaryotic phyla (≥10% relative abundance) were Basidiomycota (12.8%) and Apicomplexa (27.6%), while Bacteroidota (31.5%) and Proteobacteria (25.4%) dominated among prokaryotes (Figure 3). Under T25 conditions (25% VHW without A. platensis), eukaryotic phyla such as Evosea (26.7%) and Oomycota (15.4%) increased, while Basidiomycota (16.8%) and Apicomplexa (15.3%) decreased slightly. By contrast, the prokaryotic community shifted strongly toward Proteobacteria (97.0%), while Bacteroidota declined sharply (0.9%). Genus-level changes showed increases in Dinobryon sp., Puccinia sp., Pseudoperonospora sp., Cyclospora sp., Rostrostelium sp., and Acytostelium sp., while dominant prokaryotes included Methylophaga sp., Pseudomonas_A sp., Oceanicaulis sp., and Roseinatronobacter sp. Moreover, several moderate genera disappeared. This shift likely resulted from the alkaline pH (~10) and high salinity of the Zarrouk-based medium, which imposed osmotic stress and restricted bacterial growth. This medium relies on inorganic carbon sources, such as NaHCO_3_, and lacks the organic carbon required by many bacterial species, thereby suppressing sensitive groups. Exceptions include alkaliphilic or halotolerant bacteria, which may still grow under these conditions [43]. In addition, co-cultivation with A. platensis (T25AP) further reshaped the community, with Oomycota disappearing while Evosea (42.0%) and Basidiomycota (44.0%) increased in abundance. Among prokaryotes, Proteobacteria decreased to 11.6% and Bacteroidota to 2.9%. Certain eukaryotic genera (Dinobryon sp., Pseudoperonospora sp.) and prokaryotic genera (Methylophaga sp., Pseudomonas_A sp.) disappeared, while Cyclospora sp., Puccinia sp., and Acytostelium sp. markedly increased among eukaryotes. Similarly, Oceanicaulis sp. and Roseinatronobacter sp. decreased in abundance among prokaryotes. These results indicate that A. platensis exerts strong selective pressure on both eukaryotic and prokaryotic communities, likely through nutrient competition, enhanced oxygenation, and production of inhibitory compounds [44], highlighting its potential for suppressing undesirable or antibiotic-resistant microorganisms in wastewater treatment.
The distribution of ARGs across bacterial genera under T25 and T25AP conditions showed notable shifts in resistance profiles that may be attributed to antibiotic pressure. The presence and abundance of specific ARGs varied significantly depending on both the bacterial genus and treatment condition, suggesting a complex interplay between microbial composition, antibiotic exposure, and gene mobility [45,46,47]. One of the most striking observations was the high abundance of the aph gene in Acinetobacter sp. under the T25 condition, which decreased under T25AP. This pattern may reflect either a selective reduction in Acinetobacter sp. populations carrying the aph gene or suppression of expression due to competition with other microbial taxa under antibiotic pressure. The aph gene is known to confer resistance to aminoglycosides, and its strong signal under baseline conditions highlights the potential of Acinetobacter sp. as a reservoir of aminoglycoside resistance genes [48]. The concurrent detection of oqxB and blaOXA in Acinetobacter sp. further emphasizes its multidrug resistance potential. Methylophaga sp. exhibited the highest oqxB abundance under T25, which sharply declined under T25AP. The oqxB gene, often associated with efflux pumps conferring resistance to quinolones and other drugs, may be downregulated or lost due to competitive exclusion or horizontal gene transfer dynamics under antibiotic stress [49]. Interestingly, Pseudomonas_E sp. showed a similar pattern, with elevated oqxB levels in T25, supporting the idea that this gene is prevalent among Gram-negative opportunistic pathogens under non-stressed conditions. The T25AP treatment led to increased detection of several resistance genes across multiple genera, indicating a possible enrichment of ARG-harboring populations in response to antibiotic pressure. For instance, mph, erm, and tet genes, which typically confer resistance to macrolides, lincosamides, and tetracyclines [50], were more widely distributed or showed higher abundance in genera such as Thauera sp., Phoceicola sp., and Bifidobacterium sp. under T25AP. This suggests that these taxa may harbor inducible resistance mechanisms or have acquired ARGs through horizontal gene transfer when exposed to antibiotics.
The increased abundance of tet genes in Comamonas sp., Proteiniclasticum sp., and Phoceicola sp. under both T25 and T25AP conditions indicates that these genera may naturally harbor or maintain stable tetracycline resistance genes regardless of antibiotic exposure. On the other hand, some genera such as Gemmiger sp. and Blautia_A sp. showed minimal ARG presence, aligning with their classification as primarily gut-associated or anaerobic bacteria with limited antibiotic resistance potential [51]. Interestingly, Tolumonas sp. consistently expressed the aph gene under both treatments, possibly indicating stable aminoglycoside resistance or a niche that enables ARG maintenance even in the absence of selective pressure. This persistent presence may suggest intrinsic resistance or a conserved genetic element. It is also noteworthy that beta-lactam resistance genes, including blaOXA, blaLRC-1, and blaNPS-1, were detected at low levels. This may reflect the specificity of antibiotic selection pressure in T25AP, which might not have directly favored beta-lactam resistance, or the limited dissemination of these genes among the dominant bacterial populations in this experimental setup. Altogether, the data underscore how antibiotic exposure can reshape microbial community structure and enhance the dissemination or expression of specific ARGs. The variation in gene profiles between T25 and T25AP across bacterial genera emphasizes the need for targeted analyses to better understand resistance dynamics in microbial ecosystems.
These results indicate that A. platensis can play a key role in mitigating the spread of ARGs by altering environmental conditions and shaping microbial community composition. Although ARGs remained detectable, their abundance was reduced under A. platensis treatment, highlighting its potential in wastewater treatment. Future work should explore system scale-up, the mechanisms of antibiotic and ARG removal, and potential applications for the treated microalgal biomass.
5. Conclusions
This study demonstrates the potential of Arthrospira platensis for treating veterinary hospital wastewater. Cultivation in diluted wastewater not only enhanced biomass and pigment production but also significantly altered microbial communities and reduced antibiotic resistance gene abundance. These findings highlight A. platensis as a promising candidate for integrated wastewater treatment, offering both environmental and biotechnological benefits.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lucas D. Badia-Fabregat M. Vicent T. Caminal G. Rodríguez-Mozaz S. Balcázar J.L. BarcelóD. Fungal treatment for the removal of antibiotics and antibiotic resistance genes in veterinary hospital wastewater Chemosphere 201615230130810.1016/j.chemosphere.2016.02.11326991378 · doi ↗ · pubmed ↗
- 2Beaudoin A.L. Bollig E.R. Burgess B.A. Cohn L.A. Cole S.D. Dear J.D. Fellman C.L. Frey E. Goggs R. Johnston A. Prevalence of antibiotic use for dogs and cats in United States veterinary teaching hospitals, August 2020 J. Vet. Intern. Med.202337186410.1111/jvim.1681437526594 PMC 10473043 · doi ↗ · pubmed ↗
- 3Adebowale O.O. Jimoh A.B. Adebayo O.O. Alamu A.A. Adeleye A.I. Fasanmi O.G. Olasoju M. Olagunju P.O. Fasina F.O. Evaluation of antimicrobial usage in companion animals at a veterinary teaching hospital in Nigeria Sci. Rep.2023131819510.1038/s 41598-023-44485-w 37875528 PMC 10598005 · doi ↗ · pubmed ↗
- 4Verlicchi P. Al Aukidy M. Galletti A. Petrovic M. BarcelóD. Hospital effluent: Investigation of the concentrations and distribution of pharmaceuticals and environmental risk assessment Sci. Total Environ.201243010911810.1016/j.scitotenv.2012.04.05522634557 · doi ↗ · pubmed ↗
- 5Caddey B. Fisher S. Barkema H.W. Nobrega D.B. Companions in antimicrobial resistance: Examining transmission of common antimicrobial-resistant organisms between people and their dogs, cats, and horses Clin. Microbiol. Rev.202538 e 00146-2210.1128/cmr.00146-2239853095 PMC 11905369 · doi ↗ · pubmed ↗
- 6Thamlikitkul V. Tiengrim S. Thamthaweechok N. Buranapakdee P. Chiemchaisri W. Contamination by antibiotic-resistant bacteria in selected environments in Thailand Int. J. Environ. Res. Public Health 201916375310.3390/ijerph 1619375331590350 PMC 6801415 · doi ↗ · pubmed ↗
- 7Khan N.A. Ahmed S. Farooqi I.H. Ali I. Vambol V. Changani F. Yousefi M. Vambol S. Khan S.U. Khan A.H. Occurrence, sources and conventional treatment techniques for various antibiotics present in hospital wastewaters: A critical review Trends Anal. Chem.202012911592110.1016/j.trac.2020.115921 · doi ↗
- 8Sabri N.A. van Holst S. Schmitt H. van der Zaan B.M. Gerritsen H.W. Rijnaarts H.H.M. Langenhoff A.A.M. Fate of antibiotics and antibiotic resistance genes during conventional and additional treatment technologies in wastewater treatment plants Sci. Total Environ.202074114019910.1016/j.scitotenv.2020.14019932615424 · doi ↗ · pubmed ↗