The Role of miRNAs in Chemotherapy-Induced Cardiotoxicity
Maria Anastasiou, Evangelos Oikonomou, Panagiotis Theofilis, Maria Gazouli, Amanda Psyrri, Flora Zagouri, Gerasimos Siasos, Dimitrios Tousoulis

TL;DR
This paper reviews how microRNAs (miRNAs) are involved in chemotherapy-induced heart damage and could help detect and treat it early.
Contribution
The paper systematically summarizes miRNAs associated with chemotherapy-related cardiac dysfunction and their potential as biomarkers and therapeutic targets.
Findings
Several miRNAs, including miR-1, miR-29, and miR-30, are linked to apoptosis and oxidative stress in chemotherapy-induced cardiotoxicity.
miRNAs like let-7, miR-34a, and miR-499 are associated with fibrosis and extracellular matrix remodeling in cardiac dysfunction.
miR-130a is highlighted as a potential therapeutic agent for chemotherapy-related cardiac dysfunction.
Abstract
Cardiotoxicity is one of the most important adverse events of chemotherapy regimens, especially of anthracyclines. Different mechanisms are associated with chemotherapy-related cardiac dysfunction (CTRCD): oxidative stress, mitochondrial dysfunction, inhibition of topoisomerase 2 beta, abnormal iron metabolism, apoptosis, and fibrosis. Even after years of investigation, the early detection and prevention of cardiac impairment after chemotherapy through biomarkers remains an unmet need. The differential expression of microRNAs (miRs) in plasma at different timepoints (baseline, stable intervals during and at the end of chemotherapy) has been associated with CTRCD. Namely, some miRs, such as let-7, miR-29 and miR-30 family, miR-1 clusters, miR-34a, miR-126, miR-130a, miR-140, miR-320a, and miR-499, could play prognostic and/or diagnostic roles in CTRCD. Key miRs involved in apoptosis and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
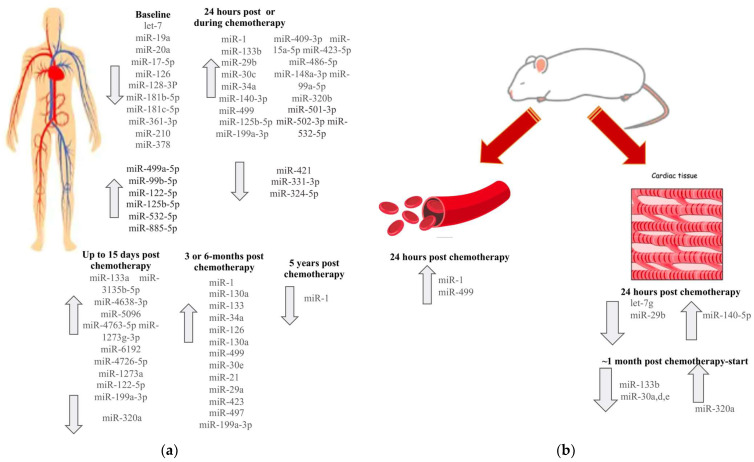 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsChemotherapy-induced cardiotoxicity and mitigation · Cardiac Ischemia and Reperfusion · Cancer-related cognitive impairment studies
1. Introduction
As survival rates of cancer patients have improved, the medical community increasingly focuses on managing adverse effects of anticancer therapies to enhance cancer survivors’ quality of life. One significant long-term adverse event is chemotherapy-related cardiac dysfunction (CTRCD), as antineoplastic agents such as anthracyclines and trastuzumab, while effective against cancer, can also cause cardiac impairment. Identifying prognostic factors of chemotherapy-induced cardiotoxicity is crucial to detect patients likely to develop CTRCD early, thereby preventing irreversible cardiac damage. Cardioprotective therapeutic strategies have been developed to mitigate this risk, including pharmacological interventions such as beta-blockers, angiotensin-converting enzyme inhibitors, angiotensin receptor blockers, mineralocorticoid receptor antagonists, statins, and dexrazoxane [1,2,3].
MicroRNAs (miRs) are small (18–25 nucleotides) non-coding, single-stranded RNA parts that are stable in the circulation, and some of them are involved in molecular mechanisms and pathways associated with CTRCD [4]. For this reason, they may serve as candidate biomarkers with prognostic value for the early detection of CTRCD. Most of the trials are focused on anthracycline-induced cardiotoxicity and the mechanisms that lead to cardiac injury and impairment. Anthracyclines are used as part of multimodal treatment against different types of cancer, such as breast cancer and sarcomas. Although they show a cardiotoxicity incidence of 10–16%, it has also been reported up to 57% of asymptomatic cardiac impairment [5,6,7]. In this narrative review, we examine the role of miRs in the molecular mechanisms underlying CTRCD, summarize clinical studies linking circulating miR profiles to cardiotoxicity risk, and explore their potential as biomarkers and therapeutic targets for improving early detection, risk stratification, and management of CTRCD.
2. Methods
We conducted a comprehensive literature search to identify studies investigating the role of miRs in CTRCD. Searches were performed in PubMed and Google Scholar databases through May 2025. The search strategy combined relevant keywords such as “chemotherapy-induced cardiotoxicity”, “CTRCD”, “cardiac dysfunction”, “microRNAs”, “miR biomarkers”, and “anthracyclines”.
3. Mechanisms of CTRCD and miRNAs’ Correlation
The chemotherapy regimens most associated with CTRCD are anthracyclines, particularly doxorubicin. MiRs play a key regulatory role in gene expression, influencing several pathophysiological mechanisms involved in CTRCD. These mechanisms and their correlations with specific miRs are summarized in Table 1.
3.1. Oxidative Stress
Oxidative stress is one of the most critical and extensively studied mechanisms of anthracycline-induced cardiac dysfunction. The iron-mediated excessive production of reactive oxygen species (ROS) in cardiomyocytes by anthracyclines renders cells vulnerable to oxidative stress, resulting in the degradation of cellular proteins, lipids, DNA, and membrane lipid peroxidation [8,9,10]. On the other hand, the dysregulation of antioxidant enzymes and pathways in cardiac and endothelial cells leads to redox imbalance and exacerbates oxidative stress. Key antioxidant enzymes—including peroxidase, catalase, and superoxide dismutase—are markedly reduced in cardiac tissue following doxorubicin exposure through mechanisms such as Nrf2 suppression and downregulation of the cAMP/PKA/SIRT1 pathway [11,12].
ROS production, antioxidant defense, and cell death pathways in cardiac cells are tightly regulated by various miRs. MiR-21 modulates key genes involved in redox homeostasis, such as SOD2 and PTEN, thereby influencing the balance between ROS generation and cardiac injury [13,14]. Chemotherapy-induced downregulation of miR-200a, together with miR-144, weakens antioxidant defenses through Nrf2 suppression and related mechanisms [15]. Similarly, miR-140-5p targets Nrf2 and Sirt2, further promoting myocardial oxidative stress [16]. Indirectly, miR-34, miR-146a, and miR-126 help to mitigate oxidative stress by suppressing anti-apoptotic proteins, reducing pro-inflammatory signaling, and enhancing angiogenesis, respectively [17,18,19,20]. Finally, miR-210 supports cardiac and endothelial cell survival by attenuating ROS levels and cellular sensitivity to oxidative injury [21,22].
3.2. Disruption of Mitochondrial Function
Mitochondria are the primary organelles responsible for ROS production, and their dysfunction is a major contributor to oxidative stress. They are abundant in cardiomyocytes and represent a key target of anthracyclines. As anthracyclines accumulate within cardiac mitochondria, they interact with iron and cardiolipin, leading to excessive ROS generation and mitochondrial dysregulation [23,24,25]. By targeting genes that are involved in mitochondrial biogenesis and function, miRs participate in mitochondrial regulation. For example, miR-181c regulates the expression of mitochondrial genes, such as mt-COX1, leading to mitochondrial ROS production [26]. MiR-155 regulates mitochondrial damage via the Nrf2/HO-1 signaling pathway, affecting the high glucose-induced cardiac fibrosis [27]. Furthermore, miR-30 family members are reduced as a response to doxorubicin treatment, disrupting mitochondrial integrity and increasing apoptosis [28].
MiR-130a exacerbates FUNDC1-mediated mitochondrial fission while suppressing mitochondrial fusion via MFN1/MFN2, leading to fragmented mitochondria and ROS accumulation [29,30]. Beyond chemotherapy-induced stress, miR-130a also amplifies mitochondrial injury in ischemia–reperfusion models [30,31]. Targeted inhibition of miR-130a may therefore represent a potential therapeutic strategy for CTRCD. Moreover, mitochondrial homeostasis is closely linked to apoptosis, and several miRs—including miR-423 and miR-499—regulate pro-apoptotic signaling through pathways such as Wnt/β-catenin and p53 [32,33].
3.3. Inhibition of Topoisomerase 2 Beta (Top2β)
The inhibitory effect of anthracyclines on Top2β has been extensively studied as a key mechanism of anthracycline-induced cardiotoxicity. When inhibited by anthracyclines, Top2β promotes DNA intercalation and double-strand breaks in cardiomyocytes, leading to apoptosis and mitochondrial dysfunction [34,35,36]. While this pathway is primarily associated with cancer resistance, it also serves as an indicator of cardiotoxicity risk. Notably, miR-9 appears to reduce TOP2B expression and may contribute to chemotherapy resistance [37].
3.4. Abnormal Iron (Fe) Metabolism
Mitochondria are the primary intracellular sites of iron (Fe) storage and regulation. A family of mitochondrial iron transporters, including MFRN1 and MFRN2, controls the transfer of Fe into the mitochondria [38]. Minotti et al. demonstrated that a metabolite of doxorubicin binds the Fe–sulfur complex of IRP-1, inducing Fe depletion. Concurrently, ROS accumulation inactivates IRP-2, promoting the binding of IRPs to iron regulatory elements, which ultimately leads to intracellular Fe accumulation [39]. Conversely, the mitochondrial exporter ABCB8 helps maintain Fe balance by facilitating its export from mitochondria. Doxorubicin disrupts this process by reducing ABCB8 protein levels in cardiomyocytes, thereby increasing mitochondrial Fe content [24,40]. Collectively, these alterations drive Fe overload, mitochondrial dysfunction, and oxidative stress.
Among the miRs involved in Fe homeostasis, miR-210 and miR-34a are the most significant. Fe deficiency and hypoxia activate miR-210 expression, positioning it as a regulator of Fe metabolism. MiR-210 decreases transferrin receptor 1 and iron-sulfur cluster scaffold protein expression, thereby influencing Fe uptake and mitochondrial function [41]. This regulatory action prevents Fe overload and subsequent cell toxicity. In contrast, miR-34a has been linked to Fe-mediated oxidative injury induced by anthracyclines [18]. Modulating these miRs, or restoring Fe balance, may represent promising strategies for cardioprotection against anthracycline-induced toxicity.
3.5. Apoptosis
Multiple pathways contribute to anthracycline-induced cardiomyocyte apoptosis. Both intrinsic and extrinsic apoptotic cascades are activated in response to anthracyclines. The intrinsic pathway is primarily regulated by the anti-apoptotic Bcl-2 protein family, which modulates Bax/Bak and caspase-3 activity. This leads to mitochondrial outer membrane permeabilization and subsequent cytochrome c release [42]. The p53 pathway, also part of intrinsic apoptosis, is strongly upregulated by doxorubicin [43,44]. In contrast, the extrinsic pathway is mediated by death receptors such as Fas, TNF-α, and TRAIL, as well as caspase-8 and -10 [44,45,46]. Additional signaling cascades, including PI3K/AKT/mTOR, ASK1, and p53, further contribute to apoptosis and autophagy regulation [47,48].
Several miRs are involved in modulating these apoptotic pathways by targeting pro- and anti-apoptotic proteins. For example, miR-21, miR-1, and miR-499 regulate caspase-9 in cancer cells, Bcl-2 in cardiomyocytes, and apoptosis-related genes in breast cancer, respectively [44]. Let-7b-5p mimics enhance apoptosis by upregulating Bcl-2 family members in keratinocyte cells [49], while miR-29b directly targets the pro-apoptotic protein Bax [50]. Similarly, reduced let-7g expression in doxorubicin-treated models downregulates Bcl-xl, thereby promoting apoptosis [51]. Let-7a suppresses pro-apoptotic genes, such as Fas ligand and Bax, in chemotherapy-treated ovarian cells [52]. In addition, miR-130a upregulation increases caspase-3 activation, thereby driving extrinsic apoptosis, while simultaneously reducing PPARγ-mediated apoptosis and inflammation [29]. Conversely, miR-133b and miR-146a exert protective effects against anthracycline-induced cardiotoxicity by reducing cardiomyocyte apoptosis and dampening pro-inflammatory signaling [53].
3.6. Fibrosis
Myocardial fibrosis is a common mechanism underlying CTRCD, driven by chronic inflammation, structural remodeling, and the disruption of the normal myocardial architecture. Activated cardiac fibroblasts promote excessive deposition of collagen and other extracellular matrix proteins in the myocardium, as a consequence of oxidative stress, mitochondrial dysfunction, calcium dysregulation, and apoptosis [54,55,56]. Matrix metalloproteases (MMPs), such as MMP-1 and MMP-2 are also impaired by anthracyclines [57]. In addition, pro-inflammatory cytokines (e.g., interleukin-6 and TNF-α), together with TGF-β and SMAD3 signaling, are upregulated following chemotherapy exposure and play key roles in fibrosis progression [58].
MiRs regulate chemotherapy-induced fibrosis through multiple pathways. For instance, miR-29b is upregulated in plasma after anthracycline treatment and has been implicated in extracellular matrix remodeling and fibrotic changes [59]. Similarly, miR-29a modulates collagen synthesis and correlates with troponin T levels [60]. In contrast, miR-155 promotes high glucose–induced cardiac fibrosis by modulating the Nrf2/HO-1 signaling pathway [27]. Upregulation of miR-21 in response to acute myocardial injury enhances PTEN inhibition and MMP-2 expression, thereby driving fibroblast proliferation and myocardial fibrosis [61]. MiR-34a also contributes to fibrosis by inhibiting SIRT1 and other cardioprotective factors against remodeling [62]. Finally, miR-126 and miR-423 have been linked to fibrotic changes through their roles in vascular injury and myocardial remodeling, respectively [60].
4. Association of miRs with CTRCD
The most common miRs associated with CTRCD are Let-7f, miR-1, miR-29, miR-34a, miR-126, miR-130a, miR-133, miR-30e and miR-140-5p, miR-320, and miR-499. Table 2 summarizes clinical trials in cancer patients that identified miRs with potential roles as biomarkers of CTRCD. However, the definition of cardiotoxicity varies across these studies, with some including subclinical markers such as troponin elevation or myocardial strain abnormalities, underscoring the complexity of diagnosing early cardiac dysfunction.
4.1. Let-7 Family
In human, let-7a, b, c, d, e, f, g, i, miR-98 and miR-202 are members of the let-7 family [79]. Some of them are differentiated after doxorubicin therapy and in cardiac impairment and fibrosis [80,81,82,83]. Let-7f and let-7g appear to be particularly involved in chemotherapy-induced cardiotoxicity. In a study of 363 breast cancer patients treated with anthracyclines and cyclophosphamide followed by docetaxel as neoadjuvant therapy, baseline let-7f levels were associated with subsequent cardiac impairment and showed a negative correlation with other cardiac biomarkers [66]. Similarly, in 179 patients with triple-negative breast cancer who received the same regimen, let-7f expression demonstrated prognostic value for a lower risk of cardiotoxicity [67]. In preclinical models, let-7g was significantly downregulated in rat heart tissue following exposure to 18 mg/kg doxorubicin, and its levels correlated with troponin T, heart rate, and pulse pressure [51].
4.2. miR-1 Clusters
MiR-1 and miR-133 form two clusters essential for skeletal and cardiac muscle development and function and are strongly linked to acute myocardial infarction [84,85,86], but the results regarding their contribution in early detection of cardiotoxicity are controversial. Within 24 h of anthracycline infusion, miR-1 upregulation showed a stronger correlation with cardiotoxicity than troponin I [65,87]. Leger et al. reported similar findings, with miR-1 elevation detected as early as 6 h after anthracycline exposure and persisting for up to 18 h [59]. Both miR-1 and miR-133 were upregulated at 3 and 6 months, correlating positively with troponins I and T, in 17 breast cancer patients receiving chemotherapy and trastuzumab [73]. Conversely, in a follow-up of more than 5 years, Totoń-Żurańska et al. found reduced miR-1 levels in cancer survivors compared with healthy controls [86]. Importantly, miR-133a levels allowed discrimination of cardiotoxicity in breast cancer patients treated with doxorubicin [68].
In mouse models, miR-1 was upregulated within 2 h of doxorubicin infusion in animals with reduced ejection fraction [87]. Findings on miR-133b are inconsistent: two studies in rats and cardiomyocytes reported downregulation [88,89], whereas breast cancer patients showed increased expression during doxorubicin treatment [65]. Functionally, miR-133b overexpression reduces fibrosis and limits apoptosis and hypertrophy in cardiomyocytes [88,90]. Similarly, miR-133a regulates proliferation, hypertrophy, and survival, positioning both miR-133a and miR-133b as potential biomarkers and therapeutic targets of cardiotoxicity.
4.3. miR-29 Family
The miR-29 family, particularly miR-29b, plays a pivotal role in cardiac remodeling, fibrosis, and apoptosis and is closely associated with CTRCD. In pediatric and young adult patients, miR-29b was the second miR upregulated within 6 h after anthracycline treatment, correlating with both anthracycline dose and high-sensitivity troponin T levels [59]. Similarly, increased miR-29a levels correlated with elevated cardiac troponin I 6 months after initiation of doxorubicin therapy. By contrast, in doxorubicin-treated rats, myocardial miR-29b was downregulated, promoting apoptosis and cardiac dysfunction [50]. Mechanistically, miR-29b regulates multiple collagens and extracellular matrix genes [91] and its downregulation may control remodeling and fibrosis after CTRCD [92]. Elevated miR-29a levels in patients with hypertrophic cardiomyopathy further support its role in cardiac fibrosis and matrix regulation—processes also relevant to CTRCD [93,94].
4.4. miR-30 Family
The miR-30 family consists of miR-30a, miR-30b, miR-30c, miR-30d, and miR-30e, and its role in CTRCD is well documented, particularly in doxorubicin exposure. In both in vivo and in vitro models, miR-30a, miR-30d, and miR-30e were downregulated through GATA-6, leading to cardiac toxicity. Overexpression of miR-30 family members reduced apoptosis and ROS levels in cardiomyocytes, underscoring their cardioprotective function [89,95,96]. Loss of miR-30 diminishes its protective effects, such as suppression of pro-apoptotic signaling, regulation of β-adrenergic pathways, and preservation of mitochondrial integrity [95,97]. Clinically, Zhou et al. observed a gradual increase in circulating miR-30c in 80 non-small cell lung cancer patients receiving chemotherapy plus bevacizumab. Elevated levels correlated with cardiotoxicity during treatment, but not after therapy completion [72].
4.5. miR-34a
MiR-34a expression rises significantly in the blood of breast cancer patients after initial doxorubicin administration and remains elevated even one year after treatment completion [70,75,78]. It is noteworthy that the study by Pierre Frères et al. employs a distinct sample collection timepoint compared to other clinical studies, with measurements taken approximately 8 days prior to surgery, and this timing varies individually for each patient [78]. Lakhani et al. reported similar findings in HER2-positive breast cancer patients, where miR-34a upregulation correlated with troponin I elevation during therapy [60]. A striking increase in miR-34a was also observed after two cycles of epirubicin [62]. Preclinical studies link miR-34a to early cardiac injury, although it does not consistently reflect changes in left ventricular ejection fraction [60,78].
4.6. miR-126
MiR-126 is one of the most extensively studied miRs, with established roles in inflammation, angiogenesis, carcinogenesis, and metastasis [98]. Reduced plasma levels of miR-126 have been documented in cardiovascular conditions including acute myocardial infarction, chronic kidney disease–related endothelial dysfunction, and stroke, supporting its prognostic value [99,100,101,102]. In cardiotoxicity, baseline miR-126 levels were significantly lower in breast cancer patients who developed cardiotoxicity following neoadjuvant epirubicin therapy, and negatively correlated with troponin I expression [66]. Conversely, high baseline miR-126 levels predicted lower risk of cardiac impairment in patients with resectable triple-negative breast cancer [67]. Lakhani et al. further reported gradual miR-126 upregulation correlating with both troponin I and T during chemotherapy [60].
4.7. miR-130a
MiR-130a, highly expressed in cardiac tissue, regulates development and apoptosis [103,104]. In a cohort of 72 breast cancer patients treated with anthracyclines and trastuzumab, 17% developed cardiotoxicity within 15 months, reflected by elevated baseline and treatment-associated miR-130a levels [64]. Preclinical studies suggest that miR-130a downregulation may serve as a therapeutic strategy against doxorubicin-induced cardiac injury [29].
4.8. miR-140
MiR-140-3p is one of the miRs upregulated in doxorubicin-treated breast cancer patients with abnormal LVEF [70]. MiR-140-5p is also increased in vivo and in vitro models after doxorubicin exposure by direct regulation of oxidative stress through Nrf2 and Sirt2 [16,105]. It is also involved in bevacizumab-mediated cardiotoxicity in cardiomyocytes targeting VEGFA signal pathway [106].
4.9. miR-320a
MiR-320a is the family member most strongly associated with cardiotoxicity. In doxorubicin-treated hearts, it regulates cardiotoxic responses through VEGFA signaling, similarly to miR-140-5p. In contrast, circulating miR-320a was reduced in chemotherapy patients, including those receiving anthracyclines. Its regulation also influences cardiac microvascular density and endothelial function by modulating proliferation and apoptosis [107].
4.10. miR-499
MiR-499 increases within 2 h of doxorubicin treatment in mice with impaired cardiac function. In young patients, it is among the earliest and most dose-dependent miRNAs elevated following anthracycline therapy [59]. In breast cancer patients, miR-499 upregulation at 6 months post-doxorubicin initiation correlated with elevated cardiac troponin T [60]. Conversely, in cardiomyocytes and animal models, miR-499-5p was downregulated, and its restoration protected against chemotherapy-induced cardiac damage by regulating p21 and inhibiting apoptosis [32].
5. Discrepancies in miR Expression Patterns
For several miRs, we observed discrepancies between tissue and blood expression. For instance, while miR-29 and miR-499 are downregulated in cardiomyocytes following doxorubicin exposure, their elevated serum levels may reflect release from damaged cardiac cells into the circulation [32,50,59,60]. Conversely, miR-320a was reduced in the circulation of patients with acute myelogenous leukemia after doxorubicin treatment, but upregulated in the hearts and cardiomyocytes of doxorubicin-treated mice [107]. Figure 1 illustrates these divergent expression patterns, highlighting the contrasting dynamics of circulating versus tissue-specific miRNAs in both human and murine models, and underscoring their potential as circulating biomarkers for monitoring cardiotoxicity.
6. Cardiotoxicity and Correlated miRs with Other Anticancer Regimes
Beyond anthracyclines, several other anticancer agents, including trastuzumab, bevacizumab, cyclosporin A, and fluorouracil (5-FU), can also induce cardiotoxicity. Among these, cardiac impairment is one of the most frequent adverse effects of 5-FU, underscoring the importance of identifying potential therapeutic biomarkers. In cardiomyocytes, 5-FU reduces miR-199-5p expression, leading to cardiac injury, whereas restoring miR-199-5p levels mitigates 5-FU–induced cardiotoxicity [108]. Similarly, miR-377 is overexpressed in cyclosporin A–treated cardiomyocytes, promoting apoptosis [99]. Upregulation of miR-1254 and miR-579 has been strongly correlated with symptomatic bevacizumab-induced cardiotoxicity, independent of acute myocardial infarction [71]. In addition, Nourmohammadi et al. reported downregulation of miR-29 and miR-30b-5p in cyclosporin A–treated rats compared with controls, linking this dysregulation to progressive fibrosis and hypertrophy [109].
7. Clinical Implication
Early detection, risk stratification, and potential therapeutic use are the most important clinical roles of miRs in CTRCD. It is well-investigated that elevations in troponin detected shortly after chemotherapy administration reflect acute and direct myocardial cell injury or necrosis and correlate with later reductions in LVEF [110,111,112]. On the contrary, B-type natriuretic peptide (BNP) levels tend to rise more gradually and remain elevated during and after chemotherapy, indicating ongoing or chronic cardiac dysfunction and heart failure and poorer cardiac outcomes over the longer term [112,113,114].
Several miRNAs, including miR-34a, miR-29b, miR-1, miR-133a, and miR-499, show consistent early upregulation after anthracycline exposure, correlating with conventional biomarkers and LVEF reduction [32,50,59,60,65,68,78]. MiR-423, in particular, has strong predictive value for cardiac injury and heart failure [78,115,116]. Risk stratification can also be guided by baseline miRNA levels: let-7f, miR-126, and miR-130a have demonstrated predictive value for cardiotoxicity risk [64,66]. Both miR-130a and miR-29a exhibit high sensitivity for detecting cardiac remodeling after chemotherapy or anti-HER2 therapy [64,78]. Therapeutically, modulation of miRNAs offers a promising strategy. Preclinical studies suggest that targeting specific miRNAs—such as miR-1, miR-30e, miR-34a, let-7, miR-21, miR-130a, and miR-499—can attenuate chemotherapy-induced cardiotoxicity [32,62,117,118,119].
Overall, circulating miRNAs represent minimally invasive and sensitive biomarkers for the early detection of subclinical CTRCD, enabling the timely identification of patients at risk before overt cardiac dysfunction occurs. Integrating longitudinal miRNA profiling with established biomarkers and cardiac imaging could provide a more comprehensive and precise risk assessment. Moreover, machine learning and systems biology approaches hold potential for defining unique miRNA expression patterns or signatures specific to CTRCD.
8. Conclusions
Several miRNAs show strong potential as biomarkers for addressing the unmet need for the early detection of CTRCD. The diverse pathways leading to CTRCD highlight the importance of differential miRNA regulation. Among the most frequently implicated are let-7f, miR-1, miR-29, miR-34a, miR-126, miR-130a, and miR-133. Standardization of detection methods and validation in larger, multicenter cohorts will be essential for clinical translation. Ultimately, combining multiple miRNAs into a biomarker panel may provide the most effective prognostic tool for early detection of CTRCD and timely implementation of cardioprotective strategies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kourek C. Touloupaki M. Rempakos A. Loritis K. Tsougkos E. Paraskevaidis I. Briasoulis A. Cardioprotective Strategies from Cardiotoxicity in Cancer Patients: A Comprehensive Review J. Cardiovasc. Dev. Dis.2022925910.3390/jcdd 908025936005423 PMC 9409997 · doi ↗ · pubmed ↗
- 2Lyon A.R. López-Fernández T. Couch L.S. Asteggiano R. Aznar M.C. Bergler-Klein J. Boriani G. Cardinale D. Cordoba R. Cosyns B. 2022 ESC Guidelines on cardio-oncology developed in collaboration with the European Hematology Association (EHA), the European Society for Therapeutic Radiology and Oncology (ESTRO) and the International Cardio-Oncology Society (IC-OS)Eur. Heart J.2022434229436110.1093/eurheartj/ehac 24436017568 · doi ↗ · pubmed ↗
- 3Haj-Yehia E. Michel L. Mincu R.I. Rassaf T. Totzeck M. Prevention of cancer-therapy related cardiac dysfunction Curr. Heart Fail. Rep.202522910.1007/s 11897-025-00697-x 39969700 PMC 11839799 · doi ↗ · pubmed ↗
- 4Bartel D.P. Micro RN As: Genomics, biogenesis, mechanism, and function Cell 200411628129710.1016/S 0092-8674(04)00045-514744438 · doi ↗ · pubmed ↗
- 5Gerodias F.R. Tan M.K. De Guzman A. Bernan A. Locnen S.A. Apostol-Alday A. Ybanez E.J. Magno J.D. Lim A. Junia A. Anthracycline-Induced Cardiotoxicity in Breast Cancer Patients: A Five-Year Retrospective Study in 10 Centers Cardiol. Res.20221338039210.14740/cr 144236660064 PMC 9822673 · doi ↗ · pubmed ↗
- 6Mata Caballero R. Serrano Antolín J.M. Jiménez Hernández R.M. Talavera Calle P. Curcio Ruigómez A. Del Castillo Arrojo S. Graupner Abad C. Cristóbal Varela C. Alonso Martín J.J. Incidence of long-term cardiotoxicity and evolution of the systolic function in patients with breast cancer treated with anthracyclines Cardiol. J.20222922823410.5603/CJ.a 2020.006232329038 PMC 9007491 · doi ↗ · pubmed ↗
- 7Kremer L.C. van der Pal H.J. Offringa M. van Dalen E.C. Voûte P.A. Frequency and risk factors of subclinical cardiotoxicity after anthracycline therapy in children: A systematic review Ann. Oncol.20021381982910.1093/annonc/mdf 16712123328 · doi ↗ · pubmed ↗
- 8Xu X. Persson H.L. Richardson D.R. Molecular pharmacology of the interaction of anthracyclines with iron Mol. Pharmacol.20056826127110.1124/mol.105.01338315883202 · doi ↗ · pubmed ↗