Comparative Anatomical and Morphometric Analysis of Eustachian Tube Across Species
Rui Li, Yueqi Wang, Huaicun Liu, Xuan Fang, Quancheng Cheng, Man Li, Huiru Ding, Chao Wang, Ziyuan Wang, Baoshi Fan, Junxiao Jia, Yu Song, Zhen Zhong, Fei Shen, Weiguang Zhang, Junxiu Liu

TL;DR
This study compares the structure of the Eustachian tube across species and finds significant differences, which could help understand middle ear diseases.
Contribution
The study introduces a new anatomical approach and identifies NOX2's role in Eustachian tube inflammation.
Findings
Rodents have shorter Eustachian tubes with cartilage and bubbles, while miniature pigs have longer, conical structures.
NOX2 levels increased by 38.6% in inflamed mice, contributing to oxidative stress and inflammation.
Inflammatory factors IL-1β and COX2 increased, causing mucosal thickening and cell infiltration in inflamed Eustachian tubes.
Abstract
Background/Objectives: The Eustachian tube (ET) is a physiological channel connecting the middle ear with the external atmosphere. The ET plays a role in maintaining the pressure balance of the middle ear, protecting it from pathogen invasion, and cleaning secretions. Eustachian tube dysfunction (ETD) can lead to middle ear diseases in animals. The ET morphological structure are different across species. Therefore, we aim to compare the anatomical and morphological of ET across species. Methods: The combined skull base–nasal approach was used to anatomy ET. Hematoxylin-eosin, luxol fast blue myelin and immunohistochemical Staining were used to observe the morphology of ET. Results: There were significant differences in the size and structure of ET among species: the rodents ET (mouse: 1.152 ± 0.084 mm; rat: 3.738 ± 0.04355 mm) is characterized by cartilage and obvious bubbles; while the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
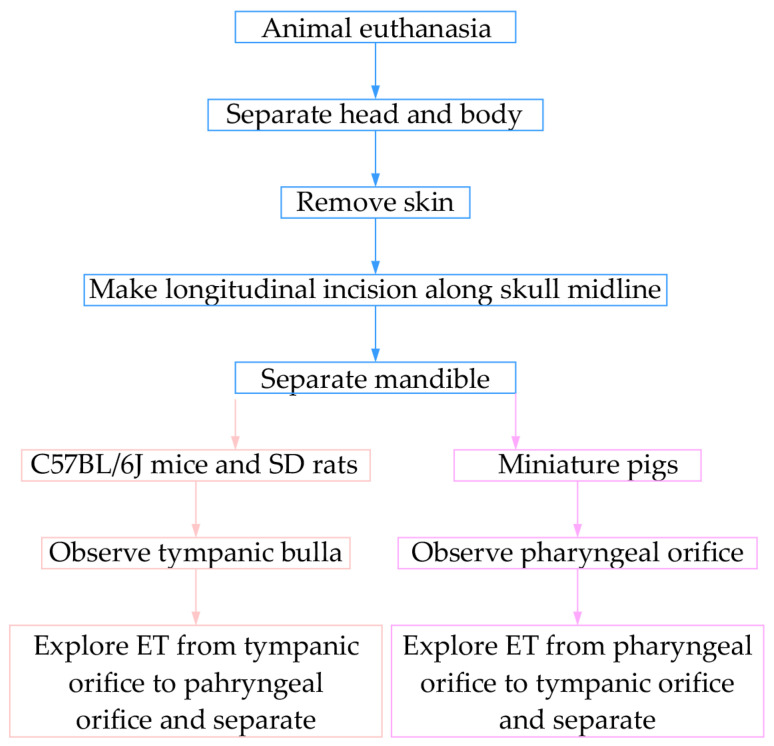 Figure 1
Figure 1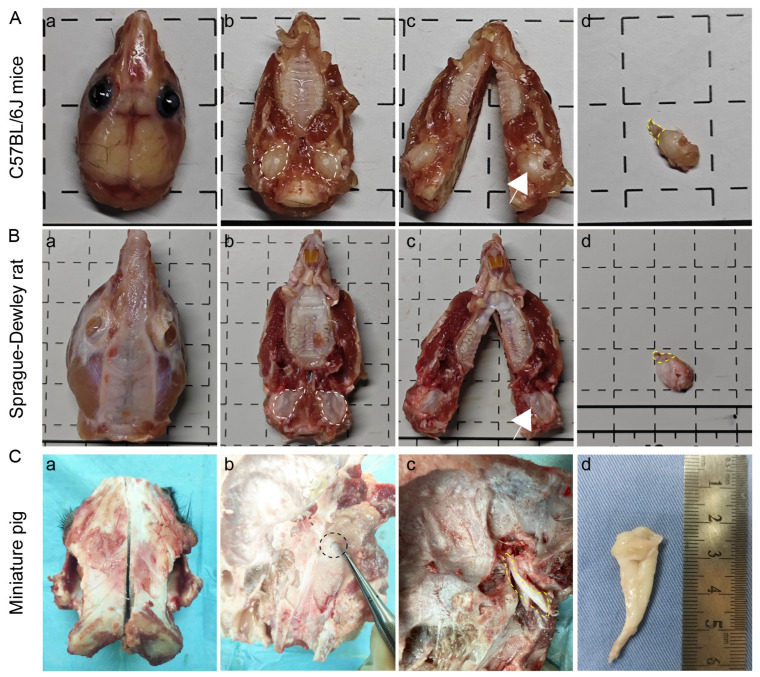 Figure 2
Figure 2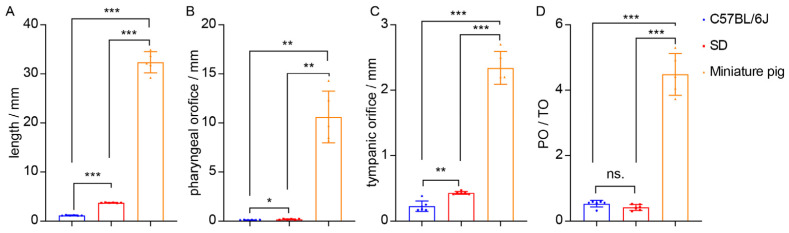 Figure 3
Figure 3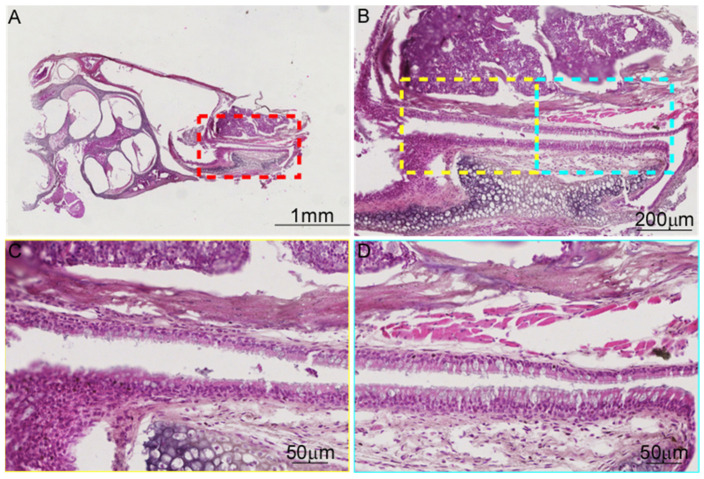 Figure 4
Figure 4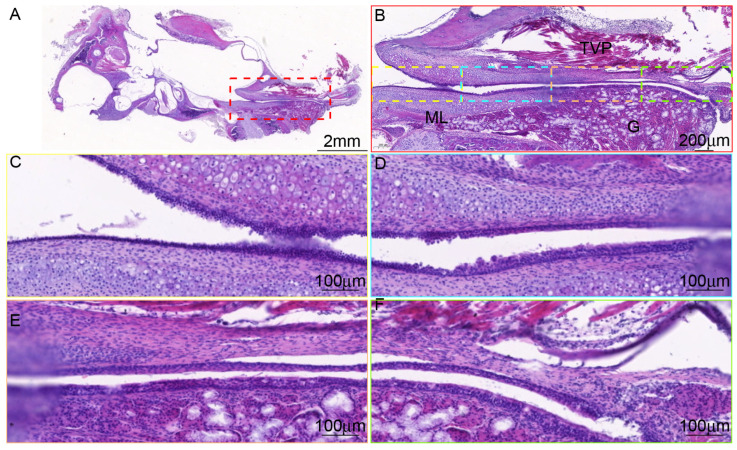 Figure 5
Figure 5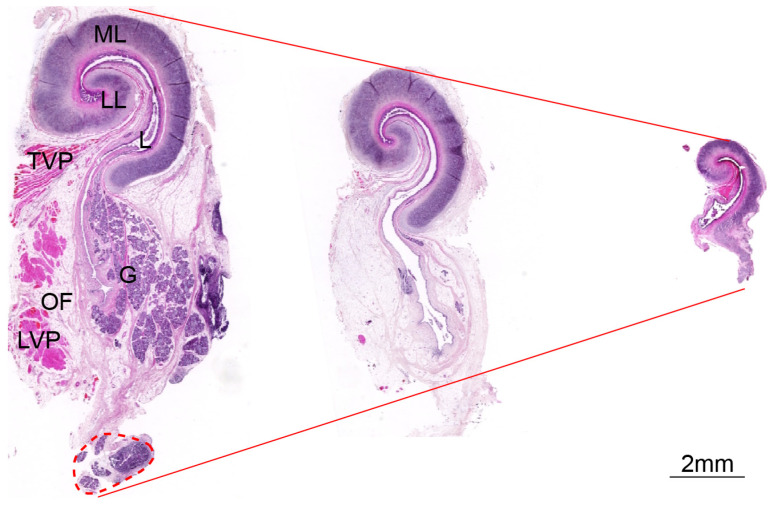 Figure 6
Figure 6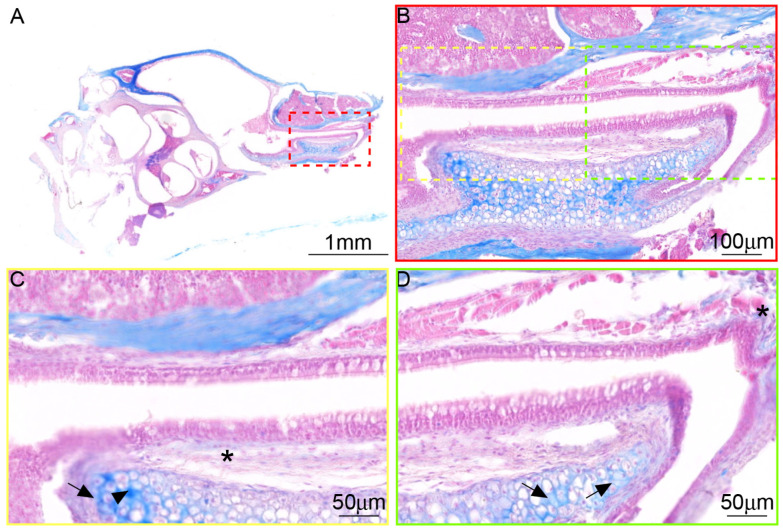 Figure 7
Figure 7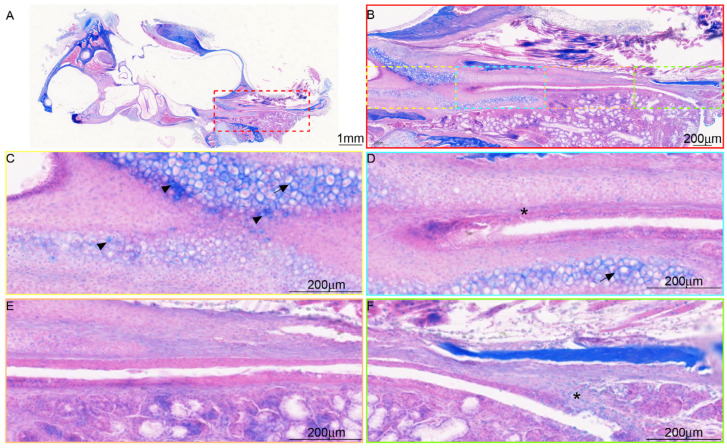 Figure 8
Figure 8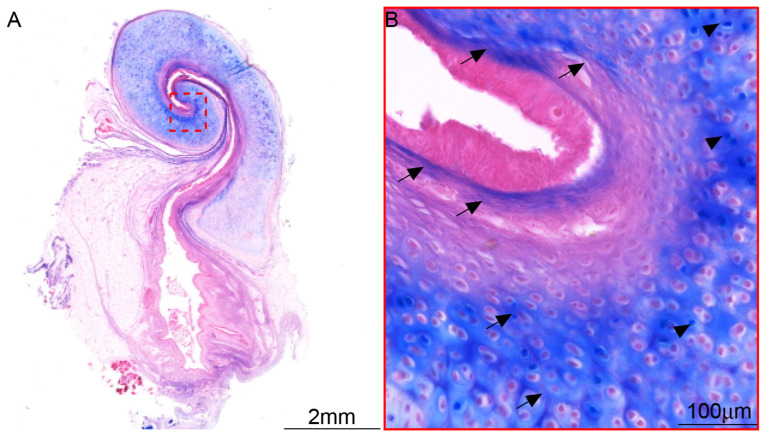 Figure 9
Figure 9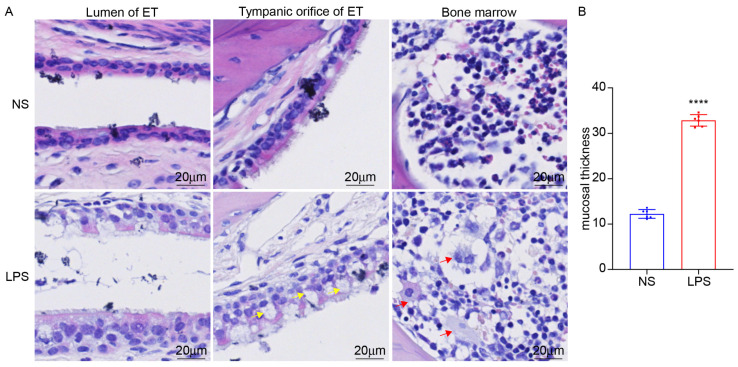 Figure 10
Figure 10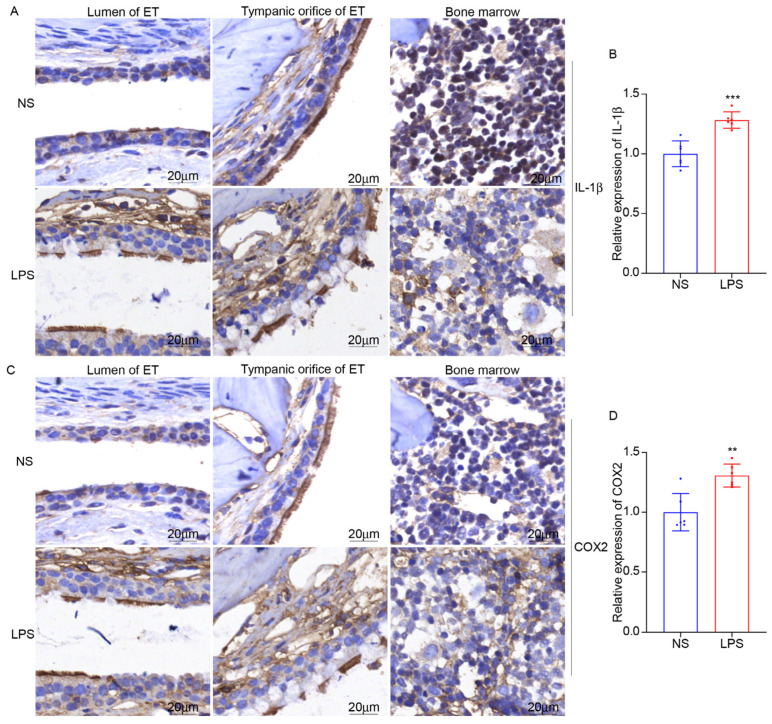 Figure 11
Figure 11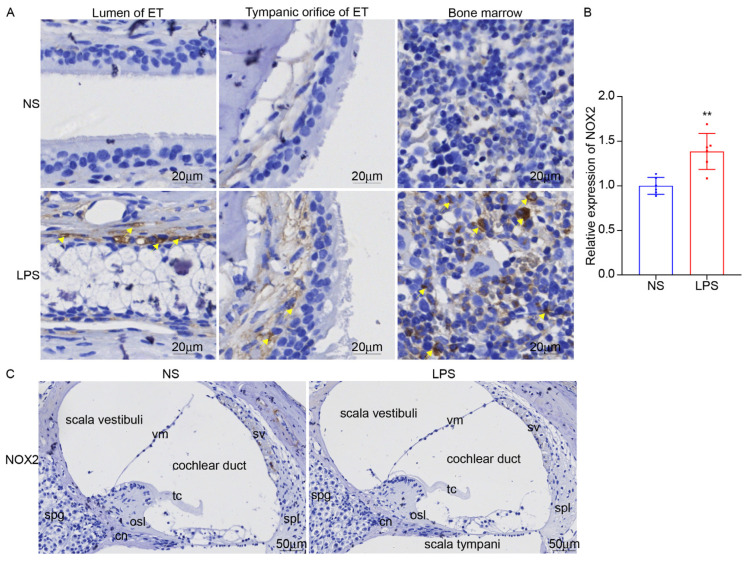 Figure 12
Figure 12- —the Key Technologies Research and Development Program of China
- —Beijing Natural Science Foundation-Haidian Cultivation Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEar Surgery and Otitis Media · Animal Vocal Communication and Behavior · Nasal Surgery and Airway Studies
1. Introduction
The Eustachian tube (ET) is the physiological channel connecting the middle ear with the external atmosphere. The incidence of Eustachian tube dysfunction (ETD) is 40% in children and 1–5% in adults [1,2], which is an important cause of protracted otitis media and hearing loss [3,4], and seriously affects the quality of life of patients [5]. The ET structure is the basis for maintaining ET function. The difficulty in obtaining human ET limited the development of ET research. Therefore, it is necessary to construct animal models. The structure of ET varies in size, structure, and tissue morphology across species. It is necessary to select the appropriate model animals. However, there is a lack of precise comparative studies on ET anatomical methods and tissue morphology across species. Therefore, it is difficult to select appropriate animal models for research.
The pathogenesis of ETD is unknown. At present, the clinical treatment of ETD is limited [6,7], and the treatment effect is poor. Therefore, the exploration of animal models of ET disease is helpful to further understand the pathogenesis of ETD [8,9]. Lipopolysaccharide (LPS) is an essential component of bacterial endotoxins and can activate macrophages and trigger the release of inflammatory cytokines in mammalian tissues. Recent studies have shown that intratympanic injection of LPS mimics acute otitis media (AOM) and can lead to morphological and functional changes in the inner ear [10,11]. However, the effects of intratympanic injection of LPS on the structure and function of ET remain unclear.
NADPH oxidase 2 (NOX2) is a key enzyme responsible for reactive oxygen species (ROS), acting as a pivotal mediator of oxidative stress, significantly influencing disease progression and treatment responses. The oxidative stress mediated by NOX2 plays a major role in the process of corneal injury and healing [12]. In Parkinson’s disease models, activation of NOX2 can mediate neuroinflammation, and inhibition of NOX2 can alleviate the neurotoxic effects associated with activated microglia [13]. This suggests that targeting NOX2 provides a compelling therapeutic strategy.
In this study, a combined skull base–nasal approach was designed for the dissection of ET tissue in mice, rats, and miniature pigs, which could completely remove ET tissue without the assistance of endoscopic instruments. Tissue staining was utilized to observe the differences in morphology and nerve fiber distribution of ET among species. To provide new ideas for the study of the pathogenesis of ETD from the perspective of comparative anatomy. At the same time, the expression changes in NOX2 in ET inflammation were explored. This serves as a reference for the pathological basis of ETD.
2. Methods
2.1. Animals and Anesthesia
Male healthy Bama miniature pigs (n = 3), aged 15–18 months, and weighing 25–30 kg were purchased from Peking University Third Clinical School of Medicine. Healthy, male Sprague-Dawley (SD) rats (n = 3), aged 6–8 weeks old, and weighing 200 g, and healthy, male C57BL/6J mice (n = 3), aged 6–8 weeks old, and weighing 20 g were purchased from the Peking University Health Science Center Department of Laboratory Animal Science. Animal experiments were reviewed and approved by the Experimental Animal Ethics Committee, Peking University Third Clinical School of Medicine (Date 21 November 2022, Approval No. SA2022576) and the Institutional Animal Care and Use Committee of Peking University Health Science Center (Date 26 March 2024, Approval No. DLASBD0161), and performed following the Chinese national guidelines for the care of laboratory animals. All efforts were made to minimize suffering.
Miniature pigs were given an intramuscular injection of Suta50+ (Virbac, Carros, France) cyrazine hydrochloride compound anesthetic. After the induction of anesthesia, tracheal intubation was performed, the ear vein was opened, and the respiratory anesthesia machine was set up with the following parameters: isoflurane, 3–3.5%; tidal volume, 12 mL/kg of body weight; and respiratory rate, 20 times/min. After entering a state of deep anesthesia (determined by a lack of response to forceps and syringe acupuncture on the abdominal skin), euthanasia was performed by rapid intravenous injection of 10% potassium chloride, and ECG monitoring was performed until the ECG waveform disappeared and the machine was withdrawn. Sprague-Dawley (SD) rats and C57BL/6J mice were deeply anesthetized with isoflurane and then sacrificed.
2.2. Anatomy
The animal’s head and body were divided, the skin was removed, and then the mandible was separated. A longitudinal incision was made through the midline of the skull. At this point, tympanic bulla was observed in mice and rats. The ET was explored from the tympanic orifice to the pharynx orifice and separated; in miniature pigs, the pharyngeal orifice was observed in the nasal cavity, and the ET was divided from the pharyngeal orifice to the tympanic orifice (Figure 1).
2.3. Hematoxylin-Eosin Staining (H&E)
The obtained ET was kept in formalin. Then, nitric acid decalcification solution (Beyotime Biotech, Shanghai, China) was added for decalcification, and then the samples were dehydrated, transparent, and paraffin embedded. Continuous sections were at a thickness of 50 μm (pig), 10 μm (mouse). A hematoxylin-eosin (H&E) staining kit (Solarbio, Beijing, China) was used for tissue staining.
2.4. Luxol Fast Blue Myelin Staining
The paraffin sections of ET were dewaxed in water. To the wash, 95% ethanol was added. The ET was stained at room temperature for 20 h in Luxol Fast Blue dyeing solution (Leagene Biotechnology, Beijing, China). Next, 95% ethanol was added to wash away excess dyeing liquid, and then the sections were rinsed with distilled water. Differentiation was performed with Luxol differentiation solution. Then, 75% ethanol was added. After rinsing with distilled water, the sections were dehydrated conventionally, cleared with xylene, and mounted with neutral balsam.
2.5. Immunohistochemical Staining (IHC)
Immunohistochemistry was performed on 5 μm paraffin sections following standard procedures. After deparaffination and antigen retrieval in citric acid buffer, sections were blocked with 3% H_2_O_2_ and permeabilized with 5% goat serum and 0.5% Triton X-100. Primary antibodies against IL-1β (Cusabio, Wuhan, China, 1:1000), COX2 (Proteintech, Wuhan, China, 1:1000), and NOX2 (Abcam, Shanghai, China, 1:1000) were applied overnight at 4 °C, followed by incubation with a universal HRP-polymer secondary antibody (ZSGB-BIO, Beijing, China). Signal was developed using DAB (ZSGB-BIO, Beijing, China), counterstained with hematoxylin, dehydrated, and mounted for microscopic observation.
2.6. Inflammatory Model
Mice were provided with an intratympanic injection (IT) of LPS (Sigma, Shanghai, China, 2.5 mg/kg, 5 mg/mL) or 10 μL saline after anesthesia for 48 h. Then, the ET was dissected using a combined nasal–cranial base approach, and further tests were conducted.
2.7. Statistical Analysis
Statistical analysis was performed by GraphPad Prism 8 software (GraphPad Software, San Diego, CA, USA). Data were presented as the mean ± standard deviation (SD) and were analyzed with the two-tailed Student t-test or Brown-Forsythe and Welch’s ANOVA test. p value of <0.05 was represented statistically significant.
3. Results
3.1. Comparative Anatomical Approaches for ET Across Species
Due to the distinct craniofacial anatomies of mice (Figure 2A), rats (Figure 2B), and miniature pigs (Figure 2C), species-specific ET dissection protocols were developed and compared. Compared with the miniature facial skull of rats and mice, the skulls of miniature pigs are thick and hard. The bony structure of the facial skull needs to be broken using a reciprocating saw, while the skull of rats and mice can be directly cut with scissors. In the exploration of the pharyngeal orifice and tympanic orifice of the ET, the tympanic orifice is obvious in miniature pigs. After removing the inner ear bone, the tympanic orifice could be observed by retrograde exploration along the ET pharyngeal orifice. However, the tympanic orifices of rats and mice are small and difficult to find, so the lateral wall of the pharynx needs to be pulled or probed from the tympanic orifice anterograde to the pharyngeal orifice.
3.2. There Is a Significant Difference in the Size of ET Across Species
The size of ET in C57BL/6J mice and SD rats was measured. The total length of ET in C57BL/6J mice is 1.05–1.28 (1.152 ± 0.084) mm. The maximum diameter of the pharyngeal orifice (PO) of ET is 0.10–0.14 (0.1133 ± 0.01633) mm, and the maximum diameter in the tympanic orifice (TO) is 0.17–0.38 (0.2283 ± 0.08085) mm. The total length of ET in SD rats is 3.68–3.80 (3.738 ± 0.04355) mm, the PO is 0.13–0.23 (0.18 ± 0.04) mm, and the TO is 0.41–0.47 (0.43 ± 0.0228) mm. The size of the ET in five ears from three miniature pigs was measured (one of the ETs was not completely removed). The total length of the ET in miniature pigs is 29.2–34.9 (32.340 ± 2.157) mm, and the PO of ET is 8.2–14.3 (10.60 ± 2.625) mm. The TO is 2.1–2.7 (2.34 ± 0.251) mm (Supplementary Table S1).
Comparative analysis shows that ET length, PO, and TO in miniature pigs are larger than rats, and that in rats are larger than mice (Figure 3A–C). There is no significant difference in the PO/TO value between rats and mice, and both have a PO/TO value less than 1, suggesting that the pharyngeal orifice is narrower than the tympanic orifice in rodents. The PO/TO value in miniature pigs is significantly higher than that in mice and rats, and the PO/TO value in miniature pigs is greater than 1, indicating that the ET tympanic orifice in miniature pigs is narrower than the pharyngeal orifice (Figure 3D).
3.3. There Are Differences in the Tissue Morphology of ET Across Species
The epithelial cells at the tympanic orifice are relatively flat, gradually transitioning to a columnar morphology toward the pharyngeal orifice. Goblet cells are sparsely distributed near the tympanic orifice, whereas both goblet cells and ciliated cells are mainly distributed near the pharyngeal orifice in mice (Figure 4), similar to human ET [14,15]. The epithelial changes from the tympanic orifices to the pharyngeal orifices were not significant, and mucus glands were found near the pharyngeal orifice in rats (Figure 5). The ET is surrounded by cartilage (Figure 4, Figure 5 and Figure 6). The lateral cartilage plate is narrow, and the medial cartilage plate is wide in miniature pigs (Figure 5). The ET cartilage at the pharyngeal orifice is shaped like the number “9”, and gradually becomes shorter toward the tympanic orifice that is shaped like “C”. ET is surrounded by muscle and adipose tissue. Cut along the bottom of the ET mucosa. The ET mucosa is smooth, the cartilage at the pharyngeal ostium is trumpet-shaped, and the cartilage at the tympanic ostium is connected with the tympanic cavity in a “V” shape (Figure 6).
3.4. The Distribution of Myelinated Nerve Fibers in ET Across Species
The nerve in ET is stained with Luxol fast blue solution in the paraffin sections, in C57BL/6J mice (Figure 7), SD rats (Figure 8), and miniature pigs (Figure 9). The comparative analysis revealed that in these three animals, the nerve fibers were mainly distributed in the ET cartilage and interwoven into a network. A small number of nerve fibers branched out in the ET submucosal tissue. The longitudinal section and transverse section of the nerve are visible. Nerves within the connective tissue of the ET lumen are aligned in the same direction as the collagen fibers, running parallel to the long axis of the lumen.
3.5. LPS Stimulation Can Induce ET Inflammation and the Expression of NOX2
We established the mouse model of ET inflammation by intratympanic injection of LPS for 48 h. The mucosa of ET was swollen, and inflammatory cells increased (Figure 10). Inflammatory factors IL-1β and COX2 in ET and the surrounding bone marrow were significantly increased (Figure 11). Intratympanic LPS stimulation can effectively induce ET inflammation in mice, which is an effective animal model of otitis media in C57BL/6J mice.
NADPH oxidase 2 (NOX2) plays a significant role in the generation of reactive oxygen species (ROS) in various physiological and pathological conditions. As a major source of ROS, NOX2 is involved in mediating oxidative stress and related damage in tissues. Inhibiting NOX2 can alleviate tissue damage in inflammatory disease models. NOX2 was significantly increased after LPS stimulation for 48 h in ET but not in the inner ear (Figure 12), suggesting that NOX2 has an important regulatory role in the inflammatory process of ET.
4. Discussion
The structure of ET shows significant interspecies differences across species, representing an adaptive selection of species to ecological pressures and physiological needs [8]. Rodents ET (mice/rats) are mainly of cartilage structure and have smaller pharyngeal openings. ET length in mice is 1.152 ± 0.084 mm, and the PO/TO is 0.5283 ± 0.1034. In rats, ET length is 3.738 ± 0.04355 mm, and the PO/TO is 0.4167 ± 0.08869, similar to mice. While in miniature pigs, ET length in mice is 32.340 ± 2.15700 mm with a smaller tympanic orifice, and the PO/TO is 4.482 ± 0.6361. Rodents (mice/rats) have obvious tympanic bulla structures, which are significantly different from miniature pigs. This may relate to the physiological need to fit rapid middle ear pressure regulation due to their cave-dwelling lifestyle. The short and narrow ET lumen and PO/TO ratio are not conducive to the rapid flow of air. To achieve a rapid balance of middle ear pressure, the tympanic bulla of rodents expands the middle ear volume, which is beneficial for maintaining middle ear pressure. The ET cartilage in miniature pigs presents a gradually conical structure towards the pharyngeal orifice, with a relatively close length to human ET [16].
The cartilaginous portion plays a critical role in the physiological function of ET by enabling its opening and closing [17]. The “9-shaped” cartilage plate of the miniature pig (Figure 6) is slightly different from the “C-shaped” of humans, but the functional regulatory structures, such as the tensor veli palatini muscle (TVP), levator veli palatini muscle (LVP), submucosal glands, and nerve distribution, are similar. The cartilage forms the top and inner, and outer walls of ET. The medial cartilage plate is wider compared to the lateral cartilage plate. The opening and closing of ET are regulated by TVP and LVP. Submucosal glands and mucosal folds can be seen in the middle and lower parts of the lumen [18]. The similarity mainly stems from the fact that both pigs and humans are terrestrial omnivores. They need to maintain the balance of middle ear pressure through frequent chewing and swallowing. In addition, the distribution of myelinated nerve fibers in ET is similar across species. The nerve fibers are mainly distributed in the cartilage. A small number of nerve fiber branches are distributed in the submucosal, suggesting that the nerves may regulate the activity of cartilage to maintain the opening and closing of ET. And we have summarized the ET structure across species (Supplementary Table S2).
ET morphological differences across species may determine the selection of model animals. ET animal models mainly originate from small and medium-sized animals such as rats, guinea pigs, and rabbits. They have similarities in morphology and structure with the human ET, but the structures are much smaller than humans. In recent years, scholars have discovered that the pig ET is similar in size and structure to the human one, which can better simulate ET cartilage [8]. We found that the mice/rats ET was more similar to the human ET bone part, which is similar to the human ET isthmus bone and cartilage, as well as the gradual narrowing of the tube cavity from the tympanic orifice to the isthmus. In addition, mice/rats ET mucosal epithelial cells were also similar to those of human ET. Goblet cells were mainly distributed in the pharyngeal orifice with dense cilia. The pig ET was more similar to the human ET cartilage part. Therefore, we hypothesized that the mice/rat might be a nice choice for studies related to bone or mucosa-related ETD, while the pig model might be suitable for research about cartilage-related ETD. Currently, the pig model is mainly used in the evaluation of balloon dilation [19]. Rodent animals are mainly used for studies related to the cleaning and protective functions of ET [20,21].
In our study, LPS was injected into mice ET to construct an ET inflammation model and explore the key molecules involved in ET inflammation. We found that NOX2 was elevated by about 38.6% after LPS stimulation, and led to ET IL-1β, and COX2 being elevated about 28.4% and 30.8%. NOX2 is a member of the NADPH oxidase family and participates in various pathological physiological processes by catalyzing the oxidation of NADPH to generate superoxide O^2−^. In macrophages, NOX2 is highly expressed and participates in immune regulation by catalyzing the NOX2/ROS pathway [22]. NOX2 can also promote the release of inflammatory factors by activating the MAPK signaling pathway [23,24]. NOX2 inhibition can reduce neural inflammation and maintain neuronal integrity [25,26]. A recent study found that NOX2 is a master regulatory molecule for the fragility of outer hair cells (OHC) in the cochlea of patients with frequent hearing loss. After NOX2 knockout, oxidative stress damage was significantly reduced. In this study, the expression of NOX2 increased in ET (Figure 12A,B), but it did not induce NOX2 expression in the inner ear (Figure 12C), suggesting that short-term LPS stimulation mainly causes NOX2-mediated oxidative stress damage to ET, but does not cause NOX2-related inner ear damage. Whether the long-term stimulation with LPS could lead to the expression of NOX2 in the inner ear remains to be explored further. Furthermore, this section mainly focuses on the morphological changes in ET. We will further explore the influence of LPS on ET function regulation in the future.
5. Conclusions
The combined skull base–nasal approach is an effective method to anatomy ET across species. Rodent species (mouse: 1.152 ± 0.084 mm, rat: 3.738 ± 0.04355 mm) tympanic bulla structures are useful for maintaining middle ear pressure. Miniature pig ET cartilage (32.34 ± 2.157 mm) gradually expands towards the pharyngeal orifice, which is similar to human cartilage structure. NOX2-mediated oxidative stress can promote the release of inflammatory factors and thicken the ET mucosa. This study provides evidence for ET sampling from different species and provides reference suggestions for the selection of model animals for different research purposes. Moreover, further exploration clarifying the key role of NOX2 in ETD is expected to provide new targets for ETD treatment and promote the advancement of ETD treatment from surgical treatment to molecular treatment.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ockermann T. Reineke U. Upile T. Ebmeyer J. Sudhoff H.H. Balloon dilatation eustachian tuboplasty: A clinical study Laryngoscope 20101201411141610.1002/lary.2095020564474 · doi ↗ · pubmed ↗
- 2Oehlandt H. Pulkkinen J. Haavisto L. Balloon dilation of the eustachian tube in chronic eustachian tube dysfunction: A retrospective study of 107 patients J. Int. Adv. Otol.20221849550010.5152/iao.2022.2137936349671 PMC 9682686 · doi ↗ · pubmed ↗
- 3Alshehri S. Musleh A. The role of eustachian tube dysfunction in recurrent chronic otitis media: A cross-sectional study of anatomical and functional variations Healthcare 2025137710.3390/healthcare 1301007739791684 PMC 11720005 · doi ↗ · pubmed ↗
- 4Bhatta R. Mishra S.K. Santana-Hernández D. Paudel A. Maharjan S. Kandel R. Shah R. Khadka K. Budhathoki M. Guragain B. Prevalence and causes of hearing impairment: A cross-sectional study in karnali province, nepal J. Laryngol. Otol.202513967168010.1017/S 002221512500019240151035 PMC 12377974 · doi ↗ · pubmed ↗
- 5Schilder A.G. Bhutta M.F. Butler C.C. Holy C. Levine L. Kvaerner K. Norman G. Pennings R. Poe D. Silvola J. Eustachian tube dysfunction: Consensus statement on definition, types, clinical presentation and diagnosis Clin. Otolaryngol.20154040741110.1111/coa.1247526347263 PMC 4600223 · doi ↗ · pubmed ↗
- 6Hamrang-Yousefi S. Ng J. Andaloro C. Eustachian tube dysfunction Disclosure: Jimmy Ng declares no Relevant Financial Relationships with Ineligible Companies. Disclosure: Claudio Andaloro Declares no Relevant Financial Relationships with Ineligible Companies Stat Pearls Publishing LLC.Treasure Island, FL, USA 2023
- 7Fallon K. Remenschneider A. Understanding eustachian tube dysfunction JAMA Otolaryngol. Head Neck Surg.20251518910.1001/jamaoto.2024.347439509097 · doi ↗ · pubmed ↗
- 8An F.W. Yuan H. Guo W. Hou Z. Cai J. Luo C. Yu N. Jiang Q. Cheng W. Liu W. Establishment of a large animal model for eustachian tube functional study in miniature pigs Anat. Rec.20193021024103810.1002/ar.2409830779320 · doi ↗ · pubmed ↗