Characterization of Antimicrobial Resistance and Hypervirulent Traits of Klebsiella variicola Isolates Collected in South Korea
Dokun Lee, Dokyun Kim, Hye Gyung Bae, Won-Jong Jang, Seok Hoon Jeong, Kyungwon Lee

TL;DR
This study examines antimicrobial resistance and virulence traits in Klebsiella variicola isolates from South Korea, finding low resistance rates but some multidrug-resistant strains.
Contribution
The study provides the first characterization of antimicrobial resistance and hypervirulent traits in K. variicola isolates from South Korea.
Findings
Most isolates were susceptible to all tested antimicrobials.
One isolate carried the blaKPC-2 gene on an IncX3 plasmid.
Ten isolates showed a mucoid phenotype, mainly with KL60 and KL61 capsular types.
Abstract
Introduction: Klebsiella variicola, a member of Klebsiella pneumoniae complex, has emerged as an opportunistic pathogen for human infection; however, antimicrobial resistance and hypervirulent characteristics of K. variicola have rarely been investigated in South Korea. Methods: We analyzed 76 clinical K. variicola isolates collected from 12 hospitals between September 2022 and October 2023. Bacterial identification was performed by MALDI-TOF MS. Antimicrobial susceptibility was tested by disk diffusion tests. Resistance determinants and virulence traits were investigated, and whole-genome sequencing was performed for hypermucoviscous or carbapenem-resistant K. variicola isolates. Results: Most (89.5%, 68/76) were susceptible to all 18 antimicrobials tested in this study, and 3 isolates harbored blaCTX-M-15. One isolate carried blaKPC-2 on its IncX3 plasmid, which is closely related to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
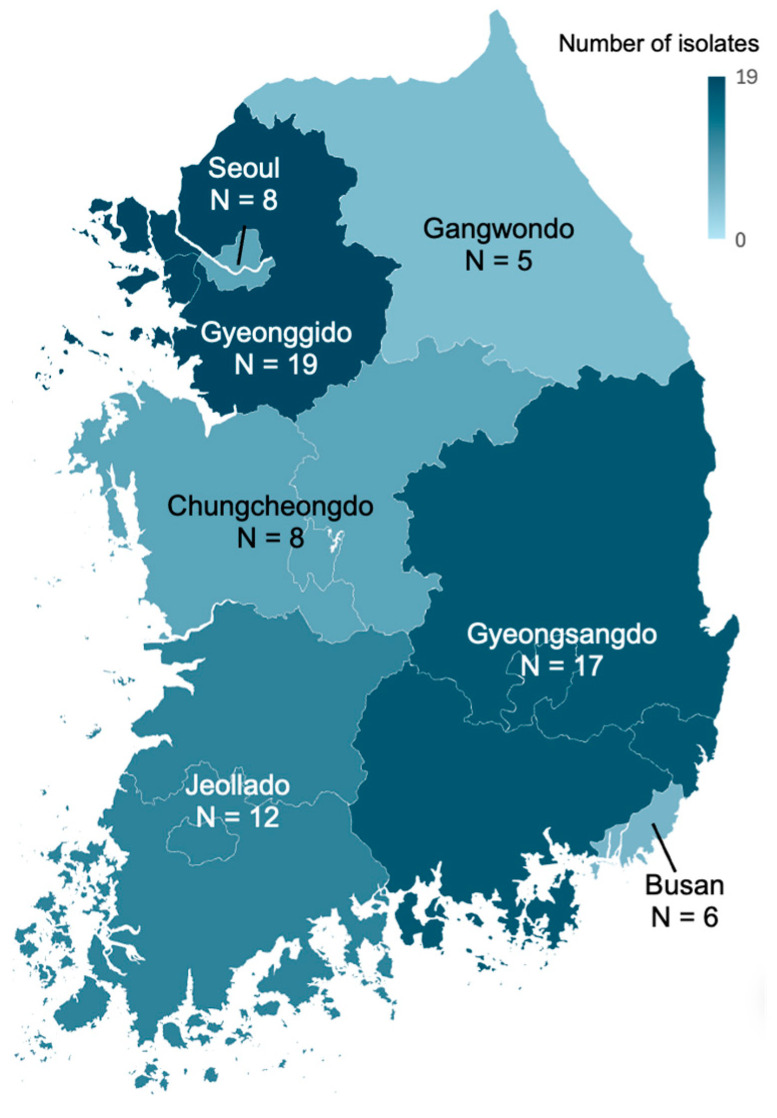 Figure 1
Figure 1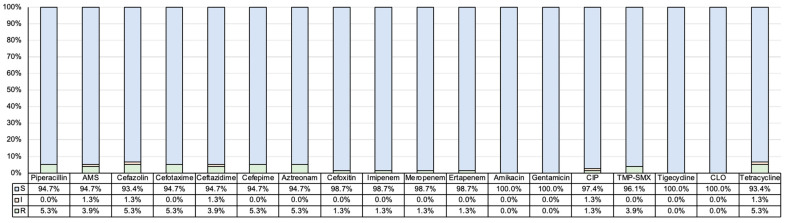 Figure 2
Figure 2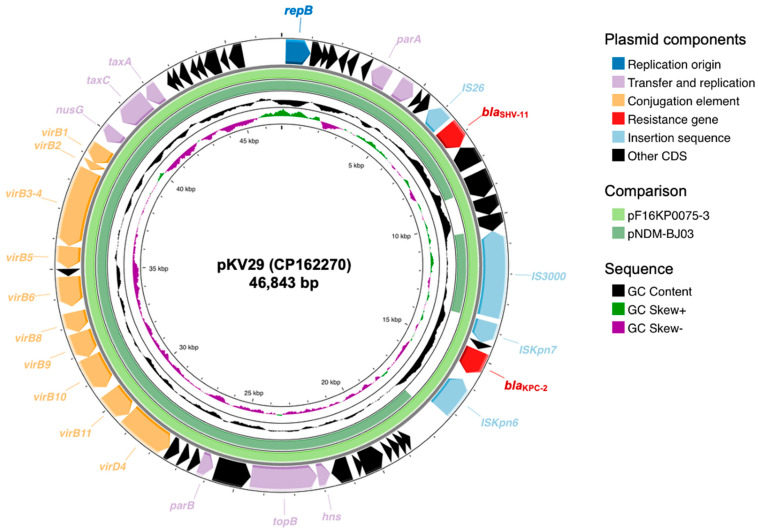 Figure 3
Figure 3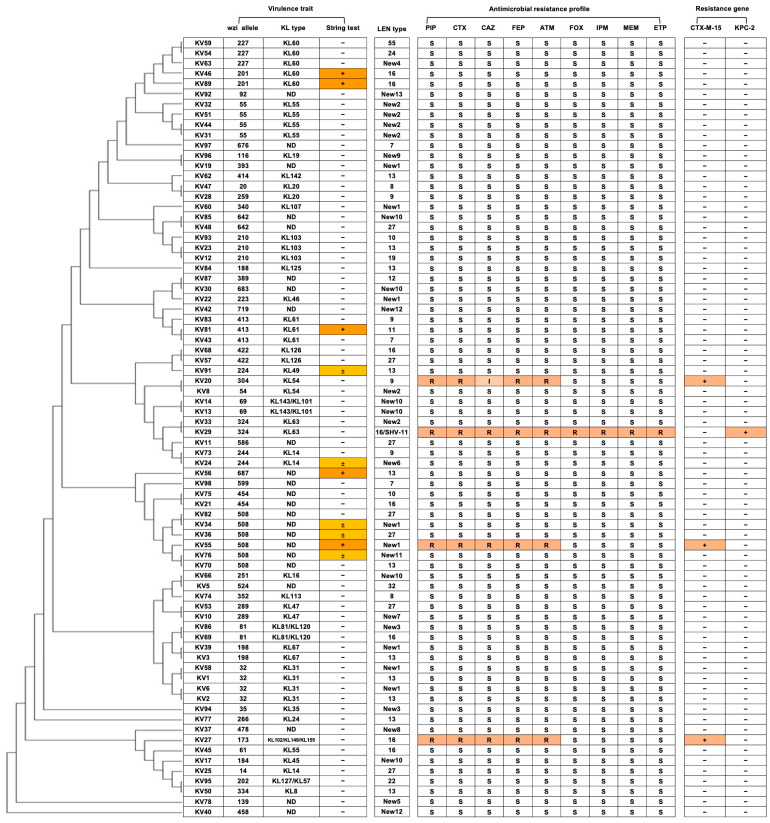 Figure 4
Figure 4- —Seoul Clinical Laboratories
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Listeria monocytogenes in Food Safety · Nephrotoxicity and Medicinal Plants
1. Introduction
The Klebsiella pneumoniae complex includes several species including K. pneumoniae, K. quasipneumoniae, K. variicola, K. qusivariicola, and K. africanensis. K. pneumoniae is one of the most important pathogens for both community-associated and hospital-originated infections, and emergence of hypervirulent (HV) and multidrug resistant (MDR) K. pneumoniae strains has been reported worldwide [1,2]. Furthermore, an outbreak of carbapenem-resistant and hypervirulent K. pneumoniae strains carrying a pLVPK-like virulence plasmid has been reported in China, which resulted in a fatal outcome for the infected patients [3].
K. variicola is another member of the Klebsiella pneumoniae complex, which was previously identified as the KpIII phylogroup of K. pneumoniae [4]. K. variicola exhibits biochemical characteristics similar to those of the other K. pneumoniae complex members, including positivity for urease, ortho-nitrophenyl-beta-galactoside, and the Voges-Proskauer test, while being negative for indole and ornithine decarboxylase [5], which causes misidentification among the K. pneumoniae complex members. For the accurate identification of K. variicola isolates, various approaches, including typing of chromosomal intrinsic β-lactamase, housekeeping gene analysis, single-nucleotide polymorphism-based average nucleotide identity analysis, and proteomic assay have been suggested [4], and Matrix-Assisted Laser Desorption Ionization Time-of-Flight Mass Spectrometry (MALDI-TOF MS) showed reliable discrimination accuracy among the K. pneumoniae complex members [6].
K. variicola causes various kinds of human infections, including bloodstream infection, urinary tract infection, and pneumonia [7]. Considering the previous misidentification of K. variicola to K. pneumoniae in the era before MALDI-TOF MS application, the public burden of K. variicola has been underscored [8]. Recently, extended-spectrum β-lactamase (ESBL-)-producing K. variicola have been reported in the US, Norway, and China [3,9,10]. Furthermore, carbapenemase-producing K. variicola strains including KPC-2, NDM-1 and NDM-5 have been identified [11]. Similarly to K. pneumoniae, hypervirulent K. variicola strains with ESBL and carbapenemase-production also emerged in China [12]. Hypervirulent K. pneumoniae complex has been associated invasive liver abscess syndrome, and poor clinical outcome has been reported when accompanied with multidrug resistance [13]. Despite the increasing concern about the dissemination of hypervirulent and MDR K. variicola strains, molecular epidemiologic research about K. variicola isolates in South Korea is limited to date.
The aim of this study is to characterize the antimicrobial susceptibility profile, related resistance determinants, and the virulence factors of K. variicola collected in South Korea.
2. Results
2.1. Collection of K. variicola Isolates
During the study period, a total of 76 K. variicola isolates were collected (Table 1 and Figure 1). About three-fourths of isolates (72.4%, n = 55) were collected from female patients, and 69.7% (n = 53) were collected from elderly patients over 60. The most common specimen type was urine (52.6%, n = 46), followed by abscess (22.4%, n = 17), blood (7.9%, n = 6), and sputum (6.6%, n = 5).
2.2. Antimicrobial Resistance Profiles and β-Lactamase Genotype of K. variicola Isolates
The resistance rate of K. variicola isolates was less than 10% in all antimicrobials tested in this study (Figure 2). Among the 76 K. variicola clinical isolates, 68 isolates (89.5%) exhibited susceptibility to all 18 antimicrobials. Only four isolates (5.3%) showed nonsusceptibility to the third-generation cephalosporins, including both cefotaxime and ceftazidime, and one of them showed carbapenem resistance.
The chromosomal SHV-OKP-LEN β-lactamase genotyping revealed that most of K. variicola isolates possessed blaLEN, except for one isolate (KV29) which carried blaSHV-11. Among the 76 blaLEN-possessing K. variicola isolates, 26 distinct LEN genotypes, including 13 novel allelic types, were identified. The most common LEN genotype was LEN-13 (n = 11), followed by LEN-New1 (n = 8), LEN-27 (n = 7), and LEN-16, LEN-New2, and LEN-New10 (n = 6 each).
Three cefotaxime-resistant K. variicola harbored blaCTX-M-15, and one cefotaxime- and ertapenem-resistant isolate (KV29) possessed blaKPC-2 in its plasmid (Figure 3). This plasmid (pKV29, GenBank accession: CP162270) belonged to the IncX3 family plasmid with 46,843 bp, and harbored conjugative type IV secretion system gene clusters, and an additional β-lactamase gene, blaSHV-11. pKV29 showed only a 7 bp difference with pF16KP0075-3 (GenBank accession: CP052170) identified in carbapenem-resistant K. pneumoniae clinical isolates in Korea [14], and the core elements of the plasmid showed >99% similarity with pNDM-BJ03 (GenBank accession: JX104760.1) [15].
2.3. Capsular Type and Virulence Traits of K. variicola
The capsular typing based on the wzi sequence revealed that 76 K. variicola isolates belonged to 51 different wzi allelic types (Figure 4). The most common capsular type was wzi508 (n = 6), followed by wzi32-KL31 (n = 4), wzi55-KL55 (n = 4), wzi210-KL103 (n = 3), wzi227-KL60 (n = 3), and wzi413-KL61 (n = 3). Ten K. variicola isolates exhibited weakly positive (n = 5) or positive (n = 5) reactions in the string test. Among the five string test-positive isolates, two belonged to wzi201-KL60, each one to wzi413-KL61, wzi687, and wzi508. Three other wzi508 isolates and each one of wzi224-KL49 and wzi244-KL14 exhibited weakly positive in the string tests. Most of hypermucoviscous isolates were susceptible to most of antimicrobials, and only one isolate (KV55) harbored the blaCTX-M-15 gene. Whole-genome sequencing revealed that all hypermucoviscous K. variicola isolates possessed virulence trait-related genes for iron acquisition (kfu), adhesin (mrk), and urease (ureA); however, common capsule production-associated genes such as magA, rmpA, or rmpA2, or salmochlelin/yersiniabactin-related gene including iroB or irp2, were not identified in any isolate. In addition, both wzi201-KL60 isolates possessed a IncF1B-type plasmid.
3. Discussion
K. variicola is a member of the K. pneumoniae complex, and automated bacterial identification systems have often caused misidentification as K. pneumoniae due to the similarity of biochemical reactions [8]. Whole-genome sequencing could be a standard for species-level discrimination within the K. pneumoniae complex, but its application in routine clinical microbiology remains limited. Through the clinical application of MALDI-TOF MS and the use of expanded reference databases, accurate species-level discrimination within the K. pneumoniae complex has become possible in clinical microbiology laboratories [6], and recent studies revealed that the actual prevalence of K. variicola infection had been underestimated [16]. Previous studies have suggested that the family of the chromosomal beta-lactamase genes could be genetic markers for species-level identification of the K. pneumoniae complex: blaSHV for K. pneumoniae, blaOKP for K. quasipneumoniae, and blaLEN for K. variicola [4]; however, the acquisition of plasmid harboring blaSHV could lead to misidentification, as seen in KV29 in this study.
Multidrug-resistant K. variicola isolates have been reported in China, Japan, European countries, and the USA [1,3,6,11], although those reports remain relatively rare. The first case of ESBL-producing K. variicola outbreak was identified in Mexico, of which the genotype of ESBL was SHV type [17]. The CTX-M-type ESBL-producing K. variicola has been identified in the US and Japan [10,16]. In our study, four isolates were identified as resistant to cefotaxime, and three of them were blaCTX-M-15-harboring K. variicola isolates, which indicates that the MDR K. variicola is still rare in South Korea and the ESBL genotype of K. variicola is still limited. The CTX-M-15 is the most common ESBL identified in K. pneumoniae isolates in Korea, accounting for approximately four-fifths of ESBL-producing K. pneumoniae, while E. coli shows a more diverse distribution of CTX-M genotypes [2]. The blaCTX-M-15-harboring K. variicola isolate, which possessed chromosomal blaLEN-16, was reported in the US, similar to the KV-27 strain in this study [10]. Carbapenemase-producing K. variicola is still rare, but KPC-2- or NDM-9-producing K. variicola strains have been reported in the US, China, India, and Chile, mostly related to ST69 and ST93 K. variicola strains [7]. Considering that K. variicola and K. pneumoniae belong to the same complex, it might be hypothesized that they could share a similar resistance gene profile or exhibit species-specific associations of mobile genetic elements at the transposon or plasmid level. One blaKPC-2-carrying isolate was identified in this study, and the plasmid structure was almost identical to that previously reported in the clinical K. pneumoniae strain collected from the patient with bloodstream infection in Korea [14]. These findings might suggest the potential for interspecies dissemination of resistance determinants within the K. pneumoniae complex.
Hypermucoviscous or hypervirulent traits of the K. pneumoniae complex have been increasingly recognized as an important pathogen for liver abscess syndrome [13]. The definition of hypervirulence in the K. pneumoniae complex has not been firmly established. However, several genetic biomarkers have been suggested, including capsule production-associated genes such as plasmid-borne rmpA and siderophore-related genes including iroB or irp2 [18]. In addition, the mucoid phenotype is often regarded as a common characteristic of hypervirulent strains. While hypervirulent K. pneumoniae has been investigated broadly, the reports of hypervirulent K. variicola remain scarce. A hypermucoviscous K. variicola strain belonging to the KL114 capsular type harboring a 343 kb plasmid (pKV8917) was reported in 2020 [19], and carbapenem-resistant and hypervirulent K. variicola KL16 strains were identified in China [3]. In this study, the capsular typing revealed 51 different wzi allelic types, which suggests heterogenic distribution rather than the presence of predominant clone. Two wzi201-KL60 strains showed a hypermucoviscous phenotype, and three out of four wzi508 strains were weakly positive or positive in the string tests. The reports about wzi201-KL60 are very rare, and the KL60 strain has been reported to exhibit diverse structural (penta- to heptasaccharides) and biochemical capsule compositions, especially enriched in glucose [20], and a hypervirulent carbapenemase-producing KL60 K. pneumoniae clone has been introduced in China [3], while the wzi508 K. variicola strain was never been reported in data. Further surveillance and functional investigation of emerging clones with diverse virulence traits should be performed.
One limitation of this study is that this study was performed in a single country with a relatively short collection period, resulting in a limited number of multidrug-resistant and hypermucoviscous isolates. Therefore, the representativeness of global or national epidemiology could not be explored. Further investigations with larger numbers of K. variicola isolates and expanding the sampling to the hospital environment, livestock, or wastewater are needed to clarify the nationwide and long-term epidemiology of this species.
4. Materials and Methods
4.1. Bacterial Isolates
K. variicola isolates from September 2022 and October 2023 from 12 hospitals were collected from Seoul Clinical Laboratories. All K. variicola isolates collected from clinical specimens during the study period were included in this study, and duplicated isolates were excluded. The sentinel hospitals are located in the 8 different districts of South Korea.
4.2. Species Identification and Antimicrobial Susceptibility Tests
Bacterial identification was performed by Bruker Biotyper (Bruker Daltonics, Billerica, MA, USA). Briefly, pure colonies were applied to a metal plate, and a matrix solution was inoculated. The protein spectra according to m/z ratio were acquired and were compared to the database version 13. Antimicrobial susceptibility tests of K. variicola isolates were performed by disk diffusion methods on Mueller-Hinton agar (Difco Laboratories, Detroit, MI, USA) following the CLSI guideline [21], with the following antimicrobials: piperacillin, ampicillin–sulbactam, cefazolin, cefotaxime, ceftazidime, cefepime, aztreonam, cefoxitin, ertapenem, imipenem, meropenem, trimethoprim–sulfamethoxazole, tigecycline, amikacin, tetracycline, chloramphenicol, ciprofloxacin, and gentamicin. Escherichia coli ATCC 25922 was used as quality control.
4.3. β-Lactamase Genotype
The chromosomal SHV-OKP-LEN β-lactamase genotyping was performed in all K. variicola isolates by PCR and sequencing with the primers (forward: 5′-CCGGGTTATTCTTATTTGTCGCT-3′; reverse: 5′-TAGCGTTGCCAGTGCTCG-3′) previously described [22]. PCR was carried out under the following conditions: initial denaturation at 94 °C for 5 min; 35 cycles of 94 °C for 30 s, 61 °C for 30 s, and 72 °C for 30 s; followed by a final extension at 72 °C for 7 min. Amplicons were sequenced by the same primers. Putative ESBL or carbapenemase producers were selected based on the resistance profile of beta-lactam antimicrobials according to the EUCAST guideline [23]. PCR and sequencing were performed for the determination of ESBL genotypes for CTX-M type and SHV, and carbapenemase genotypes for KPC, NDM, VIM, IMP, and OXA-48. The genotypes were confirmed by comparing them to the Beta-Lactamase Database (last updated on 29 March 2025; http://www.bldb.eu).
4.4. Determination of Hypervirulent Trait
String test was performed to identify the hypermucoviscous phenotype of K. variicola isolates, which is one of the key indicators of hypervirulent strains. Briefly, K. variicola isolates were inoculated on a blood agar plate (Synergy Innovation Co., Gyeonggido, Republic of Korea) and incubated overnight at 37 °C. Bacterial colonies were touched and gently pulled upward with inoculation loops. The formation of a viscous string measuring 3–5 mm was interpreted as weakly positive, and a string longer than 5 mm was considered positive. A hypervirulent K. pneumoniae strain identified as K1 capsular type in our previous study was used as a positive control [24]. For the investigation of the capsular types, PCR and sequencing of the wzi gene was performed with the primers (forward: 5′-GTGCCGCGAGCGCTTTCTATCTTGGTATTCC-3′; reverse: 5′-GAGAGCCACTGGTTCCAGAAYTTSACCGC-3′). The PCR conditions of wzi were as follows: initial denaturation at 94 °C for 5 min; 30 cycles of denaturation at 94 °C for 30 s, annealing at 55 °C for 40 s, and extension at 72 °C for 30 s; and final extension at 72 °C for 5 min. The wzi allelic types of K. variicola isolates were identified by comparing the 447 bp partial sequences of the wzi gene against the K. pneumoniae wzi database (http://bigsdb.pasteur.fr/klebsiella/, accessed on 16 September 2025). Based on the wzi allelic types, capsular type (KL type) was determined, and unassigned allelic types were designated as ‘not determined (ND)’.
4.5. Whole-Genome Sequencing of K. variicola Isolates
Microbial whole-genome sequence (WGS) analysis was performed for hypermucoviscous K. variicola isolates exhibiting weakly positive or positive in the string tests, and one carbapenem-resistant isolate. Genomic DNA of K. variicola isolates was extracted using MagMAX^TM^ Microbiome Ultra Nucleic Acid Isolation Kit (Thermo Fisher Scientifics, Waltham, MA, USA), following the manufacturer’s instructions. Briefly, 200 μL of bacterial suspension was mixed with 800 μL of lysis buffer and vortexed at 2500 rpm. Lysed samples were then vortexed for 10 min, and centrifuged at 14,000× g for 2 min. Subsequently, 40 μL of proteinase K was added, and the mixture was incubated for 20 min. After followed the four washing steps: two with wash buffer and RNase 10 μL, and two with 80% ethanol. Finally, extracted DNA was eluted in 80 μL of elution solution. The concentration of extracted DNA was measured by Qubit fluorometer (Thermo Fisher Scientifics). Libraries were prepared with extracted DNA using SMRTbell Express Template Prep Kit 3.0 (Pacific Biosciences of California, Menlo Park, CA, USA). Briefly, repair and A-tailing were performed with 46 μL of extracted DNA and 14 μL of Repair master mix, and adapter ligation was performed at 20 for 30 min with 4 μL of adapter mix, 30 μL of ligation mix, and 1 μL of ligation enhancer, followed by nuclease treatment and additional bead purification. Microbial genome was sequenced with a SMRT cell 1 M on the PacBio Sequel II system (Pacific Biosciences of California, Menlo Park, CA, USA). Raw sequence reads were assembled using PBMM2 (https://github.com/PacificBiosciences/pbmm2, last updated in February 2025, accessed on 16 September 2025), and the assembled contigs were annotated by PROKKA [25].
4.6. In Silico Analysis of Whole Genome Sequencing Data
Multilocus sequence typing (MLST) and core genome (cgMLST) of the WGS data were determined by Ridom SeqSphere version 9.0.3. Briefly, allelic types of nine housekeeping genes of the MLST scheme and 2358 core genome of the cgMLST scheme were identified using fastq files of the bacterial whole genome, and sequence type (ST) and clonal group were determined based on the allelic profiles. Virulence factors including kfuA, kfuB, kfuC, entB, cf29a, fimH, mrkD, pks, ureA, and allS were identified by comparing with the Institut Pasteur database (http://bigsdb.pasteur.fr, accessed on 16 September 2025). The plasmid replicon genes were identified by Plasmid Finder using the fastq file of the bacterial whole genome, with the database updated in January 2023 (https://cge.food.dtu.dk/services/PlasmidFinder/, accessed on 16 September 2025) [26]. The NCBI Basic Local Alignment Search Tool was used to compare the structure of plasmids, and the plasmid map was visualized using the Proksee online tool (https://proksee.ca/) [27].
4.7. Statistical Analysis
The comparisons between the groups were performed by Fisher’s exact test in categorical variables, and by the Mann–Whitney U test in continuous variables. All statistical analyses were performed using R software version 4.3.1 with the package ‘moonbook’.
5. Conclusions
This study presents the antimicrobial resistance and virulence traits of clinical K. variicola isolates in South Korea, suggesting the emergence of multidrug-resistant K. variicola strains harboring blaCTX-M-15 and blaKPC-2, and hypermucoviscous strains, such as wzi201-KL60 and wzi501 strains. Although most K. variicola isolates remained susceptible to most antimicrobials, the interspecies dissemination of resistance determinants within the K. pneumoniae complex highlights a potential threat for further dissemination. Continuous molecular surveillance is needed to understand the clinical impact and evolutionary dynamics of K. variicola.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kazmierczak K.M. Karlowsky J.A. de Jonge B.L.M. Stone G.G. Sahm D.F. Epidemiology of Carbapenem Resistance Determinants Identified in Meropenem-Nonsusceptible Enterobacterales Collected as Part of a Global Surveillance Program, 2012 to 2017 Antimicrob. Agents Chemother.202165 e 020002010.1128/AAC.02000-2033972241 PMC 8218680 · doi ↗ · pubmed ↗
- 2Kim D. Yoon E.J. Hong J.S. Choi M.H. Kim H.S. Kim Y.R. Kim Y.A. Uh Y. Shin K.S. Shin J.H. Major Bloodstream Infection-Causing Bacterial Pathogens and Their Antimicrobial Resistance in South Korea, 2017–2019: Phase I Report from Kor-GLASS Front. Microbiol.20211279908410.3389/fmicb.2021.79908435069503 PMC 8770956 · doi ↗ · pubmed ↗
- 3Gu D. Dong N. Zheng Z. Lin D. Huang M. Wang L. Chan E.W. Shu L. Yu J. Zhang R. A fatal outbreak of ST 11 carbapenem-resistant hypervirulent Klebsiella pneumoniae in a Chinese hospital: A molecular epidemiological study Lancet Infect. Dis.201818374610.1016/S 1473-3099(17)30489-928864030 · doi ↗ · pubmed ↗
- 4Rodríguez-Medina N. Barrios-Camacho H. Duran-Bedolla J. Garza-Ramos U. Klebsiella variicola: An emerging pathogen in humans Emerg. Microbes Infect.2019897398810.1080/22221751.2019.163498131259664 PMC 6609320 · doi ↗ · pubmed ↗
- 5Rodrigues C. Passet V. Rakotondrasoa A. Diallo T.A. Criscuolo A. Brisse S. Description of Klebsiella africanensis sp. nov., Klebsiella variicola subsp. tropicalensis subsp. nov. and Klebsiella variicola subsp. variicola subsp. nov Res. Microbiol.201917016517010.1016/j.resmic.2019.02.00330817987 · doi ↗ · pubmed ↗
- 6Rodrigues C. Passet V. Rakotondrasoa A. Brisse S. Identification of Klebsiella pneumoniae, Klebsiella quasipneumoniae, Klebsiella variicola and Related Phylogroups by MALDI-TOF Mass Spectrometry Front. Microbiol.20189300010.3389/fmicb.2018.0300030581423 PMC 6294014 · doi ↗ · pubmed ↗
- 7Barrios-Camacho H. Aguilar-Vera A. Beltran-Rojel M. Aguilar-Vera E. Duran-Bedolla J. Rodriguez-Medina N. Lozano-Aguirre L. Perez-Carrascal O.M. Rojas J. Garza-Ramos U. Molecular epidemiology of Klebsiella variicola obtained from different sources Sci. Rep.201991061010.1038/s 41598-019-46998-931337792 PMC 6650414 · doi ↗ · pubmed ↗
- 8Fontana L. Bonura E. Lyski Z. Messer W. The Brief Case: Klebsiella variicola-Identifying the Misidentified J. Clin. Microbiol.201957110.1128/jcm.00826-18PMC 632246030602547 · doi ↗ · pubmed ↗