Wolbachia Surface Protein (wsp) Gene Sequencing of Strains A and B in Native Aedes albopictus of Mérida, Yucatán
Henry Puerta-Guardo, Yamili Contreras-Perera, Silvia Perez-Carrillo, Azael Che-Mendoza, Karina Jacqueline Ciau-Carrillo, Manuel Parra-Cardeña, Iram Rodriguez-Sanchez, Mayra A. Gomez-Govea, Anuar Medina-Barreiro, Guadalupe Ayora-Talavera, Norma Pavia-Ruz, Abdiel Martin-Park

TL;DR
This study confirms the presence of Wolbachia strains A and B in native Aedes albopictus mosquitoes in Yucatán, offering potential for using Wolbachia to control arbovirus transmission.
Contribution
The first report of Wolbachia wsp gene sequencing in native Aedes albopictus populations in Yucatán, Mexico.
Findings
Wolbachia strains A and B were confirmed in Aedes albopictus from Mérida using wsp gene sequencing.
High nucleotide sequence homology was observed between Wolbachia from Mérida and global strains A and B.
The stability of Wolbachia strains in Aedes albopictus across geographical regions was demonstrated.
Abstract
The mosquito Aedes (Stegomyia) albopictus is an invasive species that has spread across nearly all continents and is increasingly associated with the transmission of Aedes-borne human viruses (ABVs), such as dengue (DENV), chikungunya (CHIKV), yellow fever (YFV), and Zika (ZIKV). Wolbachia-based strategies are currently among the most innovative alternative biological methods to control vector populations. Wolbachia is an endosymbiotic bacterium known to interfere with the reproductive mechanisms of vectors and, consequently, with virus replication. Previous studies have demonstrated that Ae. albopictus is naturally associated with two Wolbachia strains, A and B. This study provides additional wsp gene DNA sequencing evidence confirming that native populations of Ae. albopictus in Yucatán—an arbovirus-endemic region—naturally harbor both Wolbachia strains A and B. These findings present…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
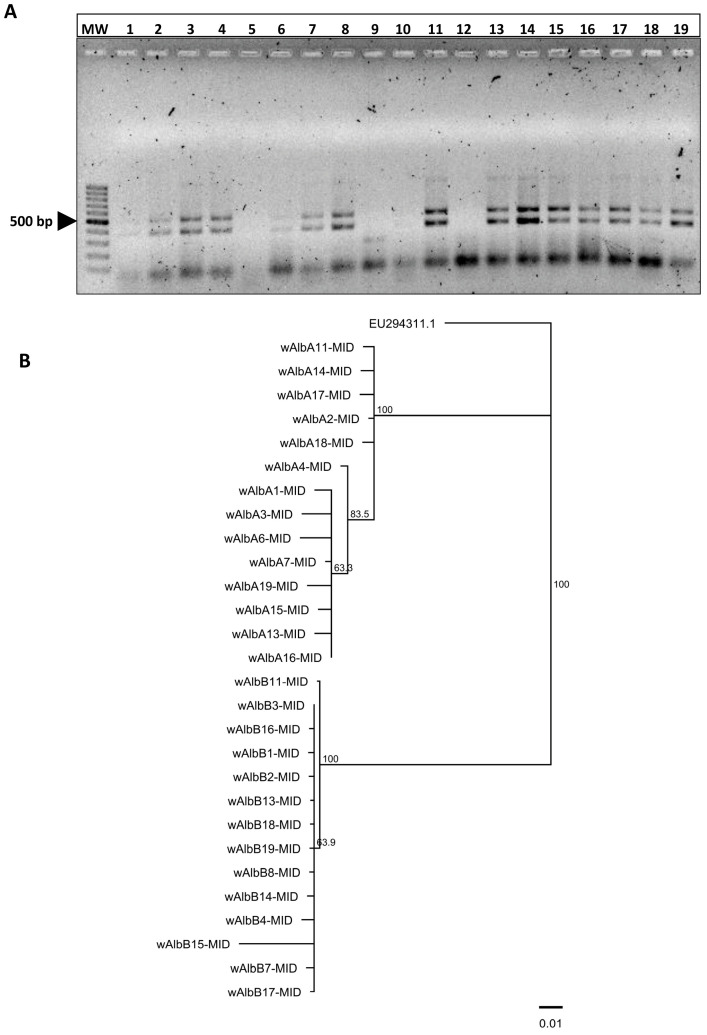 Figure 1
Figure 1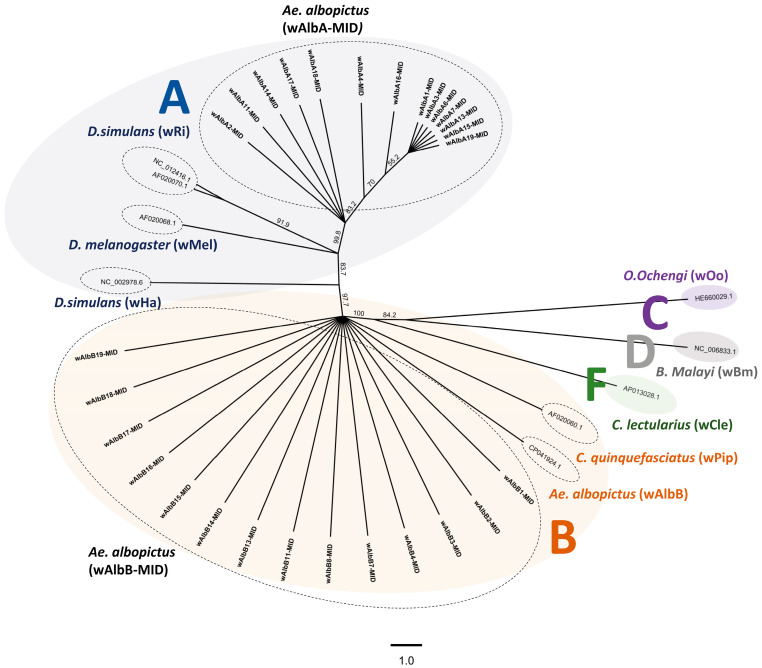 Figure 2
Figure 2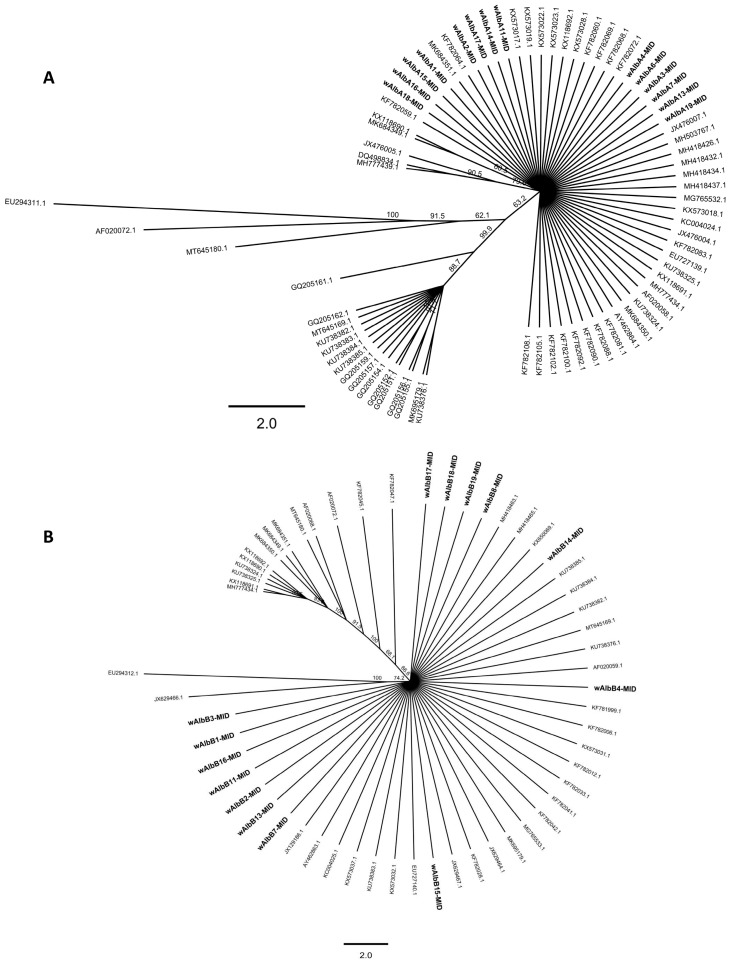 Figure 3
Figure 3- —Fondo Mixto Consejo Nacional de Ciencia y Tecnología (CONACYT)
- —(México)—Gobierno del Estado de Yucatán)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect symbiosis and bacterial influences · Mosquito-borne diseases and control · Insect and Pesticide Research
1. Introduction
Wolbachia is a diverse genus of obligate intracellular Gram-negative α-Proteobacteria (order: Rickettsiales) that can be maternally transmitted. Wolbachia have been suggested to infect at least 20% of arthropod species, where an estimated 40–65% are insects, among which 28% are mosquito species, as well as filarial nematodes of mammals and plants [1,2]. All Wolbachia strains are classified as a single species, Wolbachia pipientis, phylogenetically divided into 16 clades named supergroups, denoted from A to Q, mainly based on multilocus sequence typing (MLST) analysis as well as on amino acid sequence analyses of the Wolbachia Surface Protein (wsp) [3,4,5].
Wolbachia displays tropism for somatic and reproductive tissues of arthropod hosts, where they can manipulate their reproductive abilities by inducing cytoplasmic incompatibility (CI) [6,7]. CI confers a reproductive advantage to Wolbachia-infected females over uninfected females, with subsequent persistence and spread of Wolbachia in mosquito populations [8]. This reproductive manipulation has attracted significant interest as it plays a critical role in host biology, ecology, and evolution, as well as in the development of a symbiont-based Wolbachia-based method termed the Incompatible Insect Technique (IIT) for the control of insects of medical and agricultural importance [9].
Additionally, Wolbachia endosymbionts have been evaluated for their ability to suppress Aedes-borne human diseases (ABDs), including dengue, Zika, and chikungunya [10,11,12,13]. Along with Aedes (Stegomyia) aegypti, considered the main vector for many of these arboviral diseases, the commonly known Asian Tiger mosquito Ae. albopictus is a competent vector of ABDs [14,15]. Ae. albopictus is an aggressive biting mosquito that has invaded and colonized many countries in the Americas, Europe, Africa, and the Pacific [16,17,18].
Unlike Ae. aegypti, Ae. albopictus has been shown to naturally harbor Wolbachia, primarily strains from supergroups A and B [19,20,21,22]. Worldwide, although various programs using the release of Wolbachia-infected mosquitoes to suppress/replace natural mosquito populations have been carried out in several countries, including Australia, Brazil, Colombia, Mexico, Indonesia, and Vietnam [23,24,25], in the Americas, there are a handful of studies describing the presence of Wolbachia in wild populations of Aedes mosquitoes, including Ae. albopictus [26,27,28,29,30]. Wolbachia strains and their subgroups present in Ae. albopictus can serve as markers for inferring the geographic origin of mosquito populations, since both the Aedes spp. reservoir and bacteria strains often have specific geographic distributions, reflecting dispersal patterns and evolutionary retreats [1,2,5,19,20]. Therefore, analyzing the distribution of these strains can identify phylogenetic relationships and trace the biogeographic history of Ae. albopictus populations in different regions.
The wsp gene is commonly used as a marker for screening Wolbachia presence/infection, as well as for strain typing and phylogenetic analyses [3,4,31]. In this study, we obtained nucleotide sequences of the wsp gene obtained from Ae. albopictus that were positive for Wolbachia strains A and B according to PCR, and conducted DNA sequencing analyses to confirm the presence of these two strains and their relationship to other Wolbachia strains previously reported in Ae. albopictus and other mosquito species worldwide. In Mexico, the Ministry of Health has incorporated a Wolbachia-based strategy for the replacement of Aedes aegypti populations as part of the national dengue and arboviral disease control plan. It is anticipated that the use of Wolbachia-infected Aedes mosquitoes, including Ae. aegypti and Ae. albopictus, will expand in the coming years, not only in Mexico but also in other Latin American countries and globally. Therefore, it is essential to expand the understanding of the Wolbachia bacteria circulating in local mosquito populations, as this will facilitate the design of future Wolbachia-based intervention strategies for the control and management of mosquito-borne diseases in endemic areas.
2. Materials and Methods
2.1. Mosquito Sample Collection and Identification
In 2019 from April to December, a total of 45 Ae. albopictus adult mosquitoes were collected in distinct suburban areas of Mérida, Yucatán (abbreviated hereafter as MID) using outdoor BG-sentinel traps, as previously described in [30,32]. The identity of all mosquitoes was established based on standard taxonomic keys [33]. Field collections were performed every week from April to December of 2019 in three suburban areas located at the periphery of the city of Merida in the Peninsula of Yucatan: San Pedro Chimay (20°51′55″ N 89°34′46″ O), Hacienda Tahdzibichen (20°53′06″ N 89°35′52″ O), and Tekik de Regil (hereafter Tekik; 20°48′59″ N 89°33′39″ O). The average altitude of the localities is 9 m above sea level, with annual average temperatures ranging from 26 °C to 27 °C (36 °C max–18 °C min), relative humidity of 70–75%, and two distinct climate phases during the year: a rainy season, from May/June to October, with a rainfall of 882.5 mm, and a dry season, from November to April, with rainfall of 167.9 mm. The sociodemographic features of these localities include an average of 1200 inhabitants per locality with an average of 6 households and 31 inhabitants per hectare. These areas share similar urban and ecological characteristics, such as type of housing, and share large vegetated backyards with vegetation (coverage > 60%) [30].
2.2. DNA Extraction and Wolbachia PCR Amplification
Total genomic DNA was extracted from individual mosquitoes (n = 19, female = 11; male = 8) using a Blood and Tissue DNeasy Kit© (Qiagen, Hilden, Germany) according to the manufacturer’s instructions, with some in house modifications, as previously described in [30]. Total extracted DNA was eluted using nuclease-free water and quantified by spectrophotometry using a Nanodrop One Microvolume UV-Vis system (Thermo Scientific, Madison, WI, USA). PCR amplification of the Wolbachia DNA genome was performed in PCR reactions of 15 µL containing a DNA template (200 ng per reaction), PCR buffer (10×), MgCl_2_ (50 mM), dNTPs mix (10 mM), Taq DNA polymerase (5 U/µL), RNAse/DNAse-free water, and forward and reverse primers (10 µM) to amplify the DNA genome from Wolbachia strains A (wAlbA) and B (wAlbB) as follow: forward primers named 328F (5′-CCAGCAGATACTATTGCG-3′) and 183F (5′-AAGGAACCGAAGTTCATG-3′) for the wAlbA and wAlbB strains, respectively. For both strains, the only reverse primer 691R (5′-AAAAATTAAACGCTACTCCA-3′) was used as previously described in [30,34,35]. The PCR amplification program was performed using the following parameters: initial denaturation at 95 °C for 5 min; 40 cycles of denaturation at 95 °C for 1 min, Tm annealing at 55 °C for 1 min and extension at 72 °C for 1 min; and final extension at 72 °C for 3 min. All PCR amplifications were performed using a Mastercycler epgradient S PCR thermal cycler (Eppendorf AG, Hamburg, Germany). The presence of Wolbachia in Ae. albopictus was screened based on the amplification of 300–600 bp fragments. PCR amplicons were separated on agarose gel at 1.5% and visualized using a ChemiDoc^TM^ MP Imaging system with Image Lab software V 2.0.1 9 (Bio-Rad Laboratories, Hercules, CA, USA).
2.3. Wolbachia wsp Gene DNA Sequencing Analyses
Double-positive PCR amplicons [n = 30; walbA(+) = 15; wAlbB(+) = 15] were enzymatically cleaned up using ExoSAP-IT™ PCR Product Cleanup Reagent (Thermo Scientific) and then submitted for standard Sanger DNA sequencing (500 ng per sample) to PSOMAGEN, Inc. (formerly Macrogen Corp., Rockville, MD, USA), using the two sets of forward primers 328F and 182F, and one reverse primer, 691R (5 pmol/μL), as described above. A total of 60 linear DNA sequences were obtained from 19 analyzed samples using three sets of primers. The obtained partial sequences were processed for editing and analyzed using Geneious software version 6.1 (Biomatters. available at http://www.geneious.com accessed on 1 June 2023). Alignment between reference sequences and sequences in the study group was edited and aligned using the MUSCLE tool (v5.0-5.3, https://drive5.com/software.html accessed on 1 June 2023) supported by Geneious software version 6.1. To do so, a pairwise MUSCLE alignment analysis of all raw experimental nucleotide sequences [328F vs. 691R (n = 30), and 182F vs. 691R (n = 30), (n = 60 total sequences)] was performed against a group of reference sequences (n = 100), obtained from the GenBank dataset of the National Center for Biotechnology Information (NCBI), for the wsp gene of the wAlbA and wAlbB strains. This process allowed us to remove poorly aligned positions and to obtain non-ambiguous sequence alignments to be used in further analyses. A final consensus sequence was generated for each group of samples (n = 28), hereafter identified as wAlbA-MID (n = 14) and wAlbB-MID (n = 14). These consensus sequences were processed through the Basic Local Alignment Search Tool (BLAST, https://blast.ncbi.nlm.nih.gov/) to identify regions of similarities between biological sequences existing in the GenBank. Sequencing analyses were performed to compare Wolbachia sequences in the study group with those nucleotide sequences of Wolbachia strains A (n = 78) and B (n = 58) reported for Ae. albopictus in the GenBank database.
Rooted and unrooted maximum likelihood phylogenetic trees of the wsp gene were built using Geneious software version 6.1 to show the relationship between the wsp gene of representative Wolbachia strains detected in Ae. albopictus of Yucatán and distinct Wolbachia strains representing different supergroups described to infect Ae. albopictus worldwide. The Jukes–Cantor genetic distance substitution model was used for Bayesian analysis to infer the evolutionary history of the Wolbachia nucleotide sequences included in the study [36]. The bar indicates a branch length of 0.01. Bootstrap values were obtained from 1000 replicates (Figure 1). The GenBank accession numbers for each nucleotide sequence used in the study are included in each figure (Figure 2 and Figure 3). Finally, to provide complementary information to the wsp gene DNA sequencing data and allow a more detailed description of the potential intra-strain variability of Wolbachia strains A and B in Ae. albopictus from Yucatán, we analyzed distinct genetic diversity indices, including haplotype richness (number of haplotypes), haplotype diversity (Hd), nucleotide diversity (π), and mean genetic distance (RStudio. V4.5.1, https://cran.r-project.org/bin/windows/base/ accessed on 1 June 2023).
3. Results and Discussion
3.1. Detection of Wolbachia Strains A and B in Field-Collected Aedes albopictus from Yucatán
In previous studies, we demonstrated the occurrence of Wolbachia infection (~42%, 19/45) in field-caught Ae. albopictus collected in the suburban areas of the municipality of Mérida, Yucatán [30]. In this study, 15 out of 19 Ae. albopictus specimens showed PCR amplifications for both set of primers specific to the molecular detection of Wolbachia strains A and B of Wolbachia (molecular weights between 400 and 600 bp) (Figure 1A). These fragments belonged to supergroups A and B (Table 1). The analyzed sequences belonging to the Wolbachia strain group A (wAlbA-MID) had variable lengths of 185 and 314 nucleotides (n = 14). On the other hand, the length of the sequences belonging to the Wolbachia strain group B (wAlbB-MID) varied from 252 to 377 nucleotides length (n = 14) (Supplementary Materials).
Furthermore, multiple alignment of several representative nucleotide sequences of the study group [wAlbA-MID (n = 4), wAlbB-MID (n = 4)] and the nucleotide sequence of the wsp gene of the reference strain Wolbachia pipientis (accession: AF020070.1) confirmed these similarities and the location of the amplified nucleotide sequences from Ae. albopictus of Yucatán within the wsp gene of Wolbachia (Supplementary Figure S1A,B). Additionally, these results confirmed that Ae. albopictus harbors two Wolbachia strains from supergroups A and B.
Based on the sequence of the wsp gene, other studies have identified high variability between different Wolbachia isolates, which can be used to resolve the sequencing relationships of different Wolbachia strains [37,38]. Here, we performed a pairwise alignment (MUSCLE) analysis of nucleotide sequences from the wsp gene fragments amplified by PCR in Ae. albopictus of Yucatán. This analysis identified significant similarities within each study group: the wAlbA-MID group (n = 14) showed identity percentages ranging from 98.9 to 100%, and the wAlbB-MID group (n = 14) showed identities ranging from 96.9 and 100% (Supplementary Tables S1 and S2). The results of the sequence alignment, as well as consensus sequences for wAlbA-MID and wAlbB-MID, are presented in Supplementary Figures S2 and S3.
As expected, a comparison between the two groups (wAlbA-MID vs. wAlbB-MID) revealed limited similarities, ranging from 69.4% to less than 78% (Supplementary Table S3), clearly indicating the presence of two Wolbachia strain supergroups in Ae. albopictus. Further analysis using the tree builder tool (Geneious software) generated a cladogram that distinctly shows two separated clusters, corresponding to the wAlbA-MID group and the wAlbB-MID group (Figure 1B).
These results together support and confirm that the wAlbA-MID and wAlbB-MID strains represent two related (in terms of sequencing) but distinct groups of Wolbachia strains circulating in wild-caught Ae. albopictus of Yucatán [19,20,22,30,34]. A preliminary analysis of distinct genetic diversity indices revealed that the number of haplotypes (haplotype richness) was higher among Wolbachia strain A (7 of 14) compared to strain B (4 of 14), indicating that strain A is richer in variants. In terms of haplotype diversity (Hd), which reflects the probability that two randomly chosen sequences differ, Wolbachia strain A showed high diversity (Hd = 0.81), whereas strain B showed moderate diversity (Hd = 0.63). Similarly, nucleotide diversity (π), which measures the average number of nucleotide differences per site between two sequences, was slightly higher in Wolbachia strain A (π = 0.0063) compared to strain B (π = 0.0039), indicating that both strains are highly conserved, with strain A being slightly more variable. However, this observed genetic variability within Wolbachia strains A and B of Ae. albopictus from Yucatan must be interpreted with caution. This study was limited by the collection of mosquitoes from a single location, resulting in a small sample size (n = 45) and a limited number of nucleotide sequences (n = 19). These constraints likely reduced the resolution of intra-population genetic variability. To better understand the extent of Wolbachia genetic diversity, future studies should involve more comprehensive field collection. This includes increasing the number of sampled mosquitoes and expanding sampling efforts to include additional urban and suburban locations, ideally with replication across ecological and geographic gradients.
Molecular characterization and nucleotide sequence relationship between Wolbachia strains in Ae. albopictus of Yucatan. (A) PCR amplification and sequencing of a DNA fragment of the wsp gene of Wolbachia strains A and B in Ae. albopictus. Amplicons with lengths of ~300–400 and ~500–600 base pairs (bp) were separated by electrophoresis on agarose gel (1.5%) (lanes 1–19). DNA Marker: 100 bp (lane 1). (B) Rooted maximum likelihood phylogenetic tree of the wsp gene of 28 Wolbachia strains detected in Ae. albopictus of Yucatan. Strains are designated as wAlbA-MID (n = 14) and wAlbB-MID (n = 14), which include the initial for Wolbachia (w) followed by the abbreviated names of their host species (Ae. albopictus: Alb), the Wolbachia supergroup that these strains may belong to (A and B), an assigned sample number (1–14), and the collection site (Merida: MID). The tree was inferred using Neighbor-Joining consensus tree Nucleotide alignment (MUSCLE) of all edited Wolbachia nucleotide sequences included in the study group and rooted with the Wolbachia endosymbiont outer surface protein precursor (wsp) gene of the Ostrinia furnacalis strain (wfurA) used as an outgroup (accession: EU294311.1). The Jukes–Cantor genetic distance substitution model was used for Bayesian analysis, built using Geneious Tree Builder (Geneious Sofware v6.1.8). The numbers on the branches indicate percentage bootstrap support for major branches obtained from 1000 replicates. Only bootstrap values above 60% that support greedy clustering are shown. The bar indicates a branch length of 0.01.
3.2. Nucleotide Comparison Analyses with Wolbachia Strains in Aedes albopictus
Consensus tree analyses of the wAlbA-MID (n = 14) and wAlbB-MID (n = 14) strains, along with reference Wolbachia strains from different supergroups (A, B, C, D, and F; see Table 2), revealed distinct phylogenetic clustering. The wAlbA-MID strains grouped closely with members of supergroup A, including the wRi and wHa strains previously identified in Drosophila simulans [39,40], and wMel identified from D. melanogaster [41]. In contrast, wAlbB-MID strains clustered with supergroup B strains such as wAlbB from Ae. albopictus [42], as well as wPip from Culex quinquefasciatus [43] (Figure 2). As shown in Figure 2, both groups showed low sequence identity (<35%) with Wolbachia strains from unrelated supergroups, including the wCle strain from Cimex lectularius (Cimicidae) [44], the wBm strain from Brugia malayi (Filariidae), and the wOo strain from Onchocerca ochengi (Onchocercidae) [45,46]. A detailed comparison of representative nucleotide sequence identities, based on alignment analyses, is provided in Supplementary Table S4. These analyses further confirm the distinct phylogenetic identities of the two Wolbachia strain groups detected in Ae. albopictus of Yucatán.
The Wolbachia strains in Aedes albopictus of Yucatan belong to supergroups A and B. Unrooted maximum likelihood phylogenetic tree showing the relationship between the wsp gene of Wolbachia strains detected in Ae. albopictus of Yucatan (wAlbA-MID, n = 14; wAlbB- MID, n = 14) (in bold) and distinct Wolbachia strains representing different Wolbachia supergroups described to infect Ae. albopictus elsewhere. Wolbachia supergroup affiliations are given in colored circles and indicated by the letter inside the circle next to the host species names as follows: Drosophila simulants Riverside (wRi) (accession: AF02070.1., NC_012416.1) and Hawaii (wHa) strains (accession: AF020068.1), and D. melanogaster (wMel) strain (accession: NC_002978.6) from supergroup A; Culex quinquefasciatus (wPip) (accession: AF020060.1) and Ae. albopictus (wAlbB) strains from supergroup B (accession: CP041924.1); Onchocerca ochengi (wOo) strain of supergroup C (accession: HE660029.1); Brugia malayi (wBm) strain representing supergroup D (accession: NC_006833.1), and Cimex lectularius (wCle) strain in supergroup F. Taxon labels correspond to Wolbachia strain names as well as Genbank accession numbers. Nucleotide sequences belonging to the study group as well as their species of origin are depicted in bold. Primarily greedy clustering was used as the consensus method. The numbers on clades correspond to bootstrap values, presented as percentages, from 1000 itinerations, as well as consensus support (%) between clades. The scale bar corresponds to inferred evolutionary changes and indicates a branch length of 1.0. The Jukes–Cantor genetic distance substitution model was used for Bayesian analysis. The tree was inferred using Neighbor-Joining consensus tree Nucleotide alignment (MUSCLE) of all edited Wolbachia nucleotide sequences and built using Geneious Tree Builder (Geneious Sofware v6.1.8). The numbers on branches indicate the percentage bootstrap support for major branches obtained from 1000 replicates. Only bootstrap values of 60% or more obtained by the consensus method are shown. The bar indicates a branch length of 0.01.
We also compared the two wAlbA-MID and wAlbA-MID groups of nucleotide sequences (Figure 1) with Wolbachia sequences from supergroups A (n = 78) and B (n = 58) found in Ae. albopictus from various geographical regions worldwide (Figure 3A,B). Pairwise multiple sequence alignment revealed that the wAlbA-MID group exhibits high sequence identities (>98–100%) with Wolbachia strains of supergroup A detected in Ae. albopictus from countries including India (JX476004.1, JX476007.1), the USA (e.g., AF020058.1), Malaysia (e.g., KX573028.1, KC004024.1, MH418426.1), Pakistan (MH503767.1), Taiwan (AY462864.1), México (e.g., MK684349.1, KX118691.1), Italy (EU727139.1), China (KU738324.1), and Sri Lanka (MH777434.1) (Figure 3A, Table 3).
Nucleotide sequence comparison of wAlbA and wAlb-B MID with Wolbachia strains belonging to supergroups A and B found in Ae. albopictus worldwide. Unrooted phylogenetic tree layout for comparison of wsp gene sequences obtained from all Wolbachia strains detected in Ae. albopictus of Yucatan: (A) wAlbA-MID (n = 14) and (B) wAlbB-MID (n = 14) (in bold) and other nucleotide sequences of Wolbachia strains detected in Ae. albopictus reported in GenBank [supergroup A (n = 78); supergroup B (n = 58)]. Strains of the study group are designated as wAlbA-MID or wAlbB-MID, as explained in Figure 1B. The tree was inferred using Neighbor-Joining consensus tree Nucleotide alignment (MUSCLE) of all edited Wolbachia nucleotide sequences included in the study group and other Wolbachia sequences of Ae. albopictus worldwide. The Jukes–Cantor genetic distance substitution model was used for Bayesian analysis, and the phylogeny tree was built using Geneious Tree Builder (Geneious Sofware v6.1.8). The taxon labels correspond to the Wolbachia strain names in the study group as well as the GenBank accession numbers. The numbers on branches indicate percentage bootstrap support for major branches obtained from 1000 replicates. Only bootstrap values above 60% or more obtained by the consensus method are shown. The bar indicates a branch length of 2.
Interestingly, lower sequence similarities (<80%) were observed between Wolbachia strains A from Brazil (e.g., GQ205162.1), India (e.g., JX476005.1), China (e.g., KU738376.1), México (e.g., MK695179.1), and Singapore (e.g., MT645169.1). These strains were separately grouped into a distinct clade.
For the wAlbB-MID group, the multiple alignment analyses revealed high sequence similarity (>98%) when compared Wolbachia strain B previously described in various countries, including India (JX629464.1), Malaysia (JX129186.1, KF781999.1), Pakistan (KX650069.1), the USA (MG765533.1), Italy (EU727140.1), Taiwan (AY462863.1), China (KU738369.1), and Singapore (MT645169.1) (Figure 3B, Table 4). In contrast, fewer sequences, such as those reported in Mexico (e.g., KX118691.1, MK684351.1) and China (e.g., KU738324.1), showed less than 90% sequence similarity (<80%) with the wAlbB-MID group of Merida.
Overall, our results suggest that most of the Wolbachia strains naturally occurring in populations of Ae. albopictus of Mérida are very similar to other Wolbachia strains belonging to supergroups A and B of Ae. albopictus described worldwide. Interesting, nucleotide sequences belonging to the wAlbA-MID group showed lower sequence identities (<80%) compared to Wolbachia strains previously identified as part of supergroup A in Brazil (e.g., GQ205162.1). In contrast, the results for Wolbachia strains in supergroup B were more consistent with previous studies from Asia (Sri Lanka, India, China, Malaysia, and Thailand), which reported that Ae. albopictus and, as well as other Aedes-related species such as Ae. Quadrivittatus, harbored Wolbachia strain B with strong bootstrap support [50,51,52,53,54]. Additional genetic markers such as MLST, including additional Wolbachia genes (e.g., gatB, coxA, hcpA, ftsZ, and fbpA), will enable more in-depth phylogenetic analyses to better define the genetic diversity among Wolbachia strains in Ae. albopictus and other Aedes species. It is well established that Ae. albopictus naturally carries either one strain (wAlbA or wAlbB) or both strains simultaneously (wAlbA and wAlbB) [46,50].
Despite this, Ae. albopictus continues to expand as an important vector for arbovirus transmission [14,15,16,17]. From a vector control perspective, this poses a challenge for the use of Ae. albopictus in Wolbachia-based control programs, which aim to suppress mosquito populations or reduce arbovirus infections in humans. Notably, recent evidence shows that Ae. albopictus artificially infected with four different Wolbachia strains (wMel, wMelPop, wRi, and wPip) have established stable lines with diverse CI patterns and reduced vector capacity for arbovirus transmission [55,56]. These findings support the feasibility of applying mass rearing and integrated SIT to control Ae. albopictus, as has already been implemented for Ae. aegypti worldwide.
4. Conclusions
Here, we confirmed the presence of two Wolbachia strains (supergroups A and B) in field-collected Ae. albopictus from suburban areas of Mérida, Yucatán, through wsp gene sequencing. These results align with previous reports of Ae. albopictus naturally harboring Wolbachia A and B worldwide, showing high sequence homology with strains from Asia and North America, but lower similarity to strains from Brazil, suggesting regional differentiation. Historical records indicate that Ae. albocpictus was introduced in the Americas in 1983 via used-tire shipments to the USA, and later detected in Brazil (1986) and Mexico (1988) [57]. The distribution of related and unrelated Wolbachia strains across distant regions likely reflects mosquito dispersal through natural migration or human activities, such as trade. These findings suggest historical connectivity among mosquito populations, with phylogenetic analysis offering insights into the biogeographic history of Ae. albopictus.
In Merida, Ae. albopictus has only recently been identified in urban and periurban areas of this municipality [32], limiting specimen availability for this study. Despite this constraint, our work provides the first description of Wolbachia gene sequencing in Aedes mosquitos of Yucatán. These results establish a foundation for future research with larger sample sizes and broader collection areas to assess the abundance, distribution, and diversity of Wolbachia strains in Aedes populations across Yucatán but also Mexico.
Although wsp genes are widely used in phylogenetic studies [58], single-gene analyses may be limited by recombination among Wolbachia strains [59]. Our complementary nucleotide diversity (π) analyses showed that strain A was slightly more diverse than strain B (π = 0.0063 vs. 0.0039), though both exhibited low variability, consistent with closely related endosymbionts such as Wolbachia A and B. Future studies incorporating additional markers (e.g., gltA, groEL, ftsZ) and multilocus sequence typing (MLST) [60,61] will better characterize Wolbachia diversity in Yucatán and México.
In conclusion, our results provide baseline evidence for the presence of Wolbachia strains A and B in wild Ae. albopictus populations of Yucatán. These findings can inform future Wolbachia-based intervention strategies, leveraging locally circulating Wolbachia strains to reduce mosquito populations and mitigate arboviral transmission in endemic regions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hilgenboecker K. Hammerstein P. Schlattmann P. Telschow A. Werren J.H. How many species are infected with Wolbachia?—A statistical analysis of current data FEMS Microbiol. Lett.200828121522010.1111/j.1574-6968.2008.01110.x 18312577 PMC 2327208 · doi ↗ · pubmed ↗
- 2Werren J.H. Baldo L. Clark M.E. Wolbachia: Master manipulators of invertebrate biology Nat. Rev. Microbiol.2008674175110.1038/nrmicro 196918794912 · doi ↗ · pubmed ↗
- 3Zhou W. Rousset F. O’Neil S. Phylogeny and PCR-based classification of Wolbachia strains using wsp gene sequences Proc. Biol. Sci.199826550951510.1098/rspb.1998.03249569669 PMC 1688917 · doi ↗ · pubmed ↗
- 4Baldo L. Dunning Hotopp J.C. Jolley K.A. Bordenstein S.R. Biber S.A. Choudhury R.R. Hayashi C. Maiden M.C.J. Tettelin H. Werren J.H. Multilocus sequence typing system for the endosymbiont Wolbachia pipientis Appl. Environ. Microbiol.2006727098711010.1128/AEM.00731-0616936055 PMC 1636189 · doi ↗ · pubmed ↗
- 5Lefoulon E. Bain O. Makepeace B.L. d’Haese C. Uni S. Martin C. Gavotte L. Breakdown of coevolution between symbiotic bacteria Wolbachia and their filarial hosts Peer J 20164 e 184010.7717/peerj.184027069790 PMC 4824920 · doi ↗ · pubmed ↗
- 6O’Neill S.L. Giordano R. Colbert A.M. Karr T.L. Robertson H.M. 16S r RNA phylogenetic analysis of the bacterial endosymbionts associated with cytoplasmic incompatibility in insects Proc. Natl. Acad. Sci. USA 1992892699270210.1073/pnas.89.7.26991557375 PMC 48729 · doi ↗ · pubmed ↗
- 7Sinkins S.P. Wolbachia and cytoplasmic incompatibility in mosquitoes Insect. Biochem. Mol. Biol.20043472372910.1016/j.ibmb.2004.03.02515242714 · doi ↗ · pubmed ↗
- 8Xi Z. Khoo C.C. Dobson S.L. Wolbachia establishment and invasion in an Aedes aegypti laboratory population Science 200531032632810.1126/science.111760716224027 · doi ↗ · pubmed ↗