Effectiveness of Curcumin on Oxidative Stress in Goat Semen: Explorations Regarding Semen Quality, Sperm Apoptosis, Ultrastructure, and Markers of Oxidative Stress
Zhaoxiang An, Wenjuan Xun, Hanlin Zhou, Guanyu Hou, Liguang Shi

TL;DR
Curcumin at 25 μM improves goat semen quality by reducing oxidative stress and protecting sperm structure and function.
Contribution
This study demonstrates curcumin's protective effects on goat sperm under oxidative stress through multi-omic and ultrastructural analyses.
Findings
Curcumin at 25 μmol/L improved sperm motility, membrane integrity, and acrosome stability while reducing ROS and apoptosis.
Curcumin elevated antioxidant enzyme activities (GPX, CAT, SOD) and restored lipid and metabolic pathways in sperm.
Curcumin preserved sperm ultrastructure and reduced mitochondrial and chromatin damage, supported by gene expression changes.
Abstract
Oxidative stress induces reactive oxygen species (ROS) accumulation, which compromises sperm DNA integrity, cellular homeostasis, and semen quality in Hainan black goats. This study aimed to mitigate ROS-mediated sperm damage by examining the protective effects of curcumin on metabolic regulation and sperm structural integrity. Semen samples were treated in vitro with varying concentrations of curcumin (5, 25, 50 μmol/L) under oxidative stress conditions. The intermediate concentration (25 μmol/L) was most effective at enhancing sperm quality. Following treatment, sperm motility, membrane integrity, and acrosome stability were significantly improved (p < 0.05), while ROS levels and apoptosis rates decreased. Antioxidant enzyme activities—glutathione peroxidase (GPX, p < 0.05), catalase (CAT, p < 0.05), and superoxide dismutase (SOD, p < 0.05)—were markedly elevated. Metabolomic analysis…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
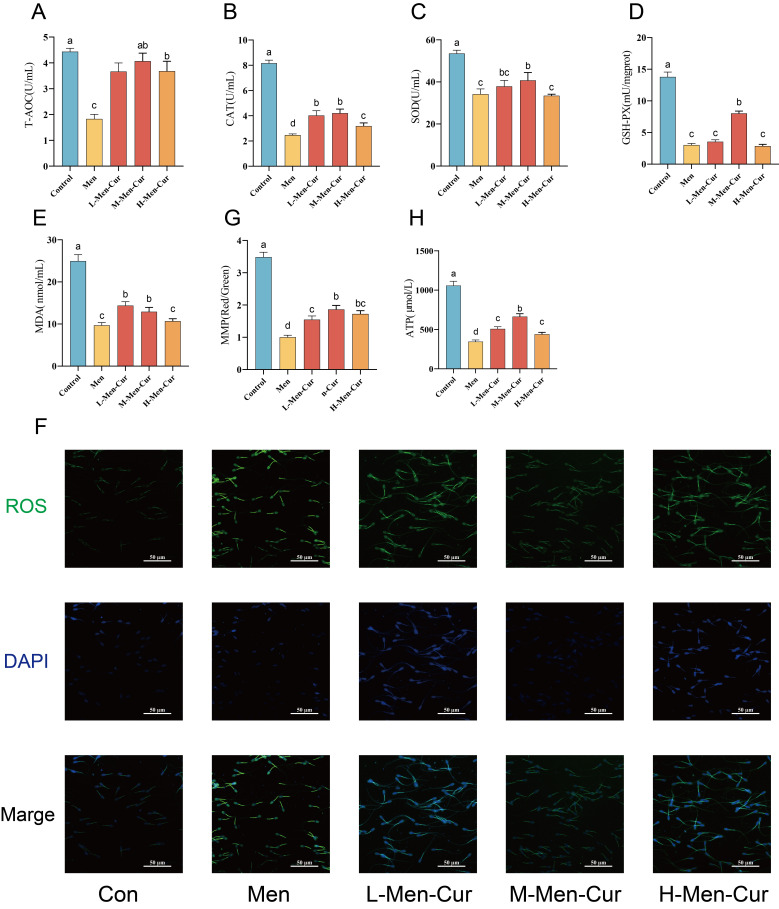 Figure 1
Figure 1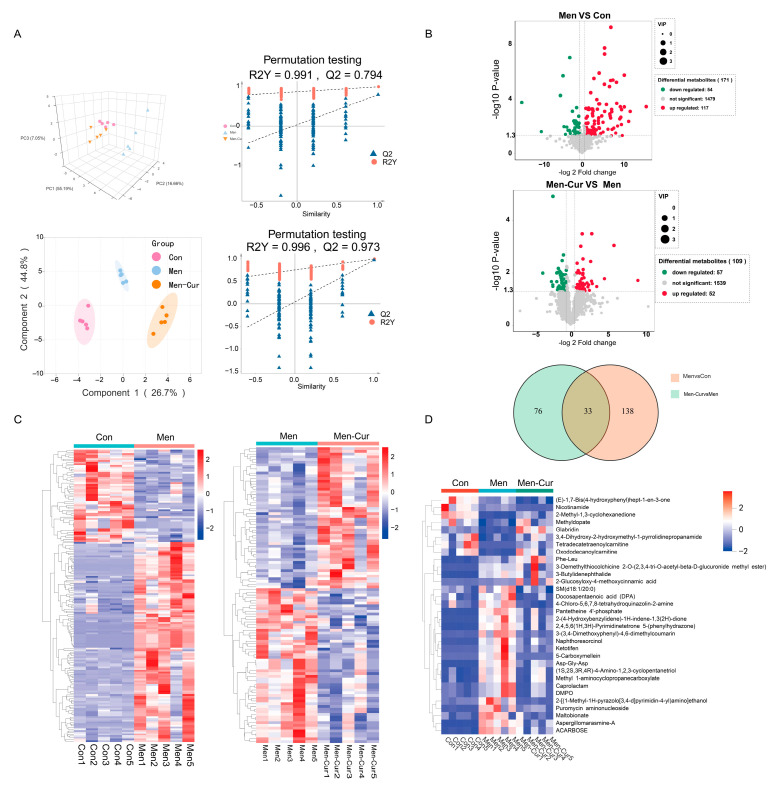 Figure 2
Figure 2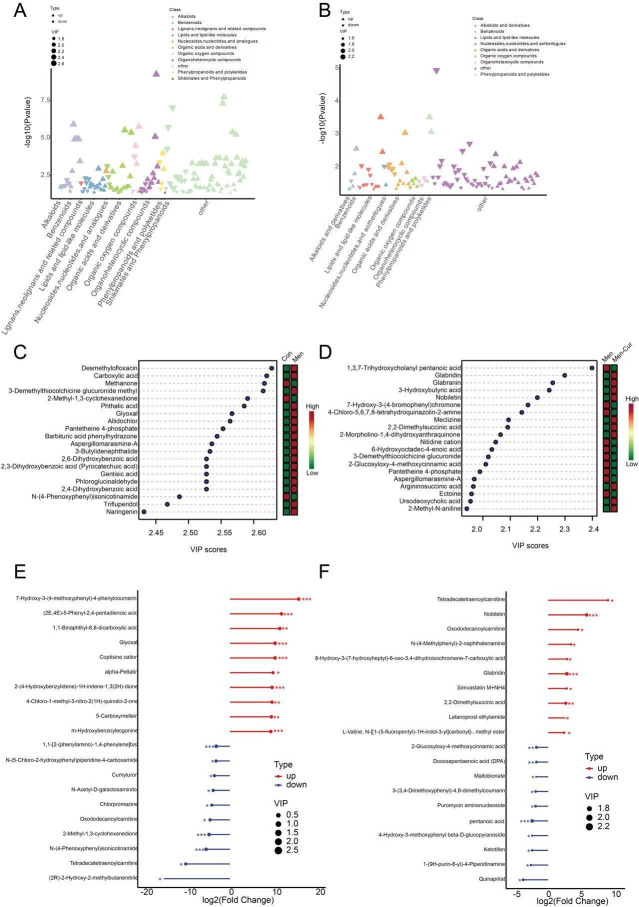 Figure 3
Figure 3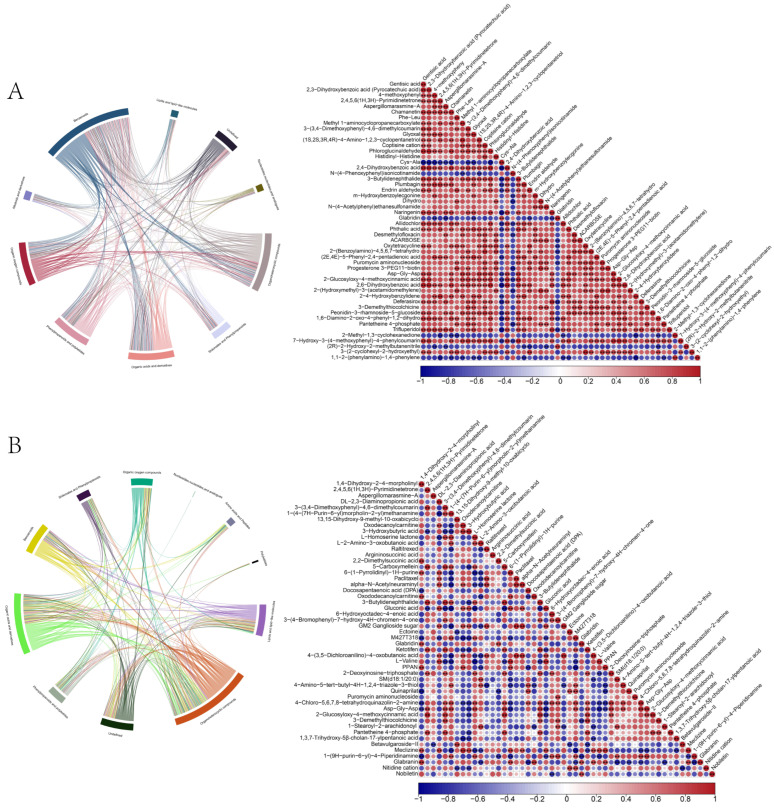 Figure 4
Figure 4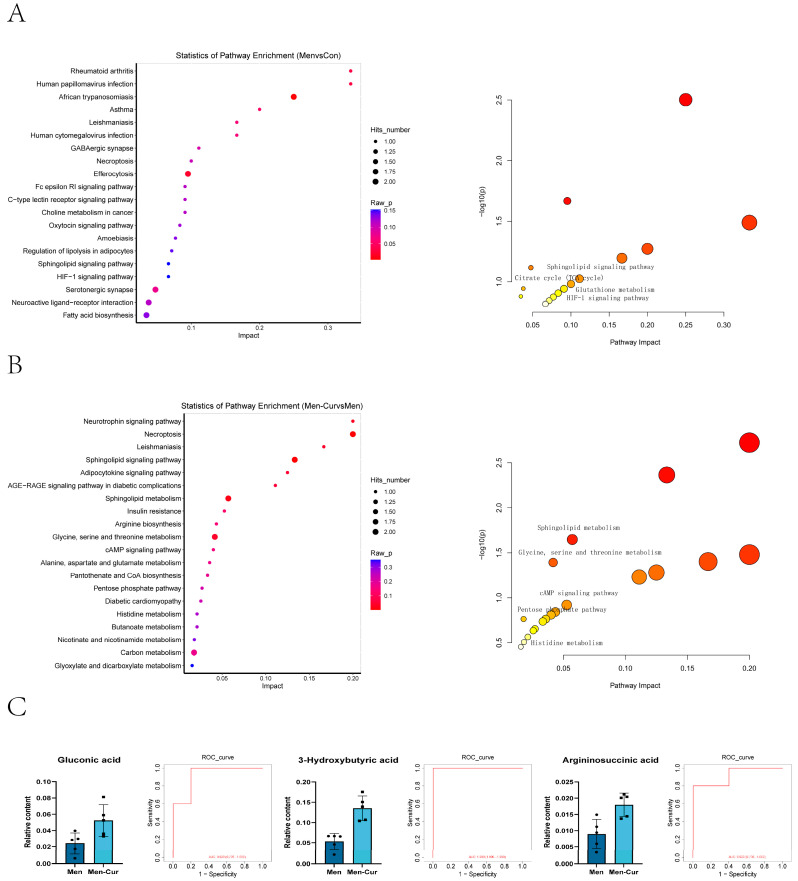 Figure 5
Figure 5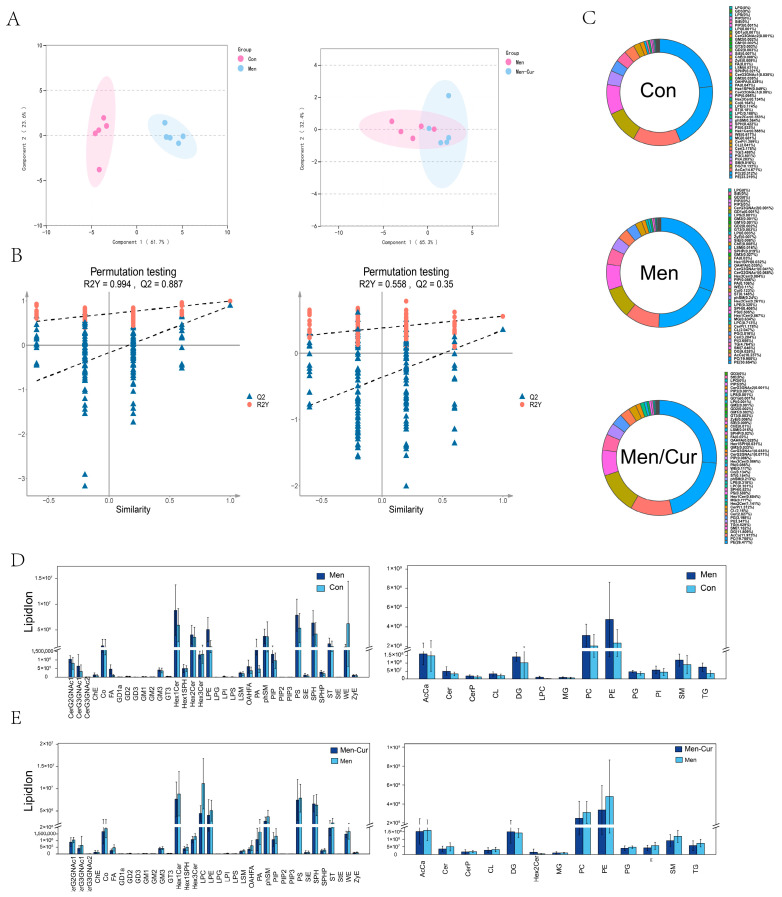 Figure 6
Figure 6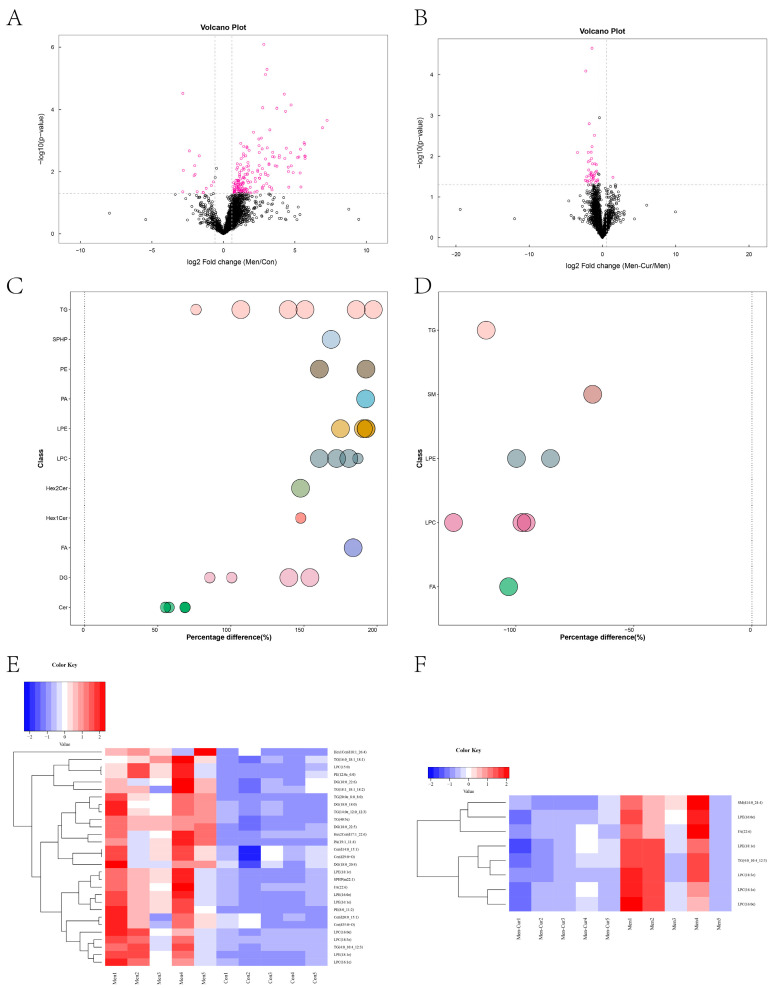 Figure 7
Figure 7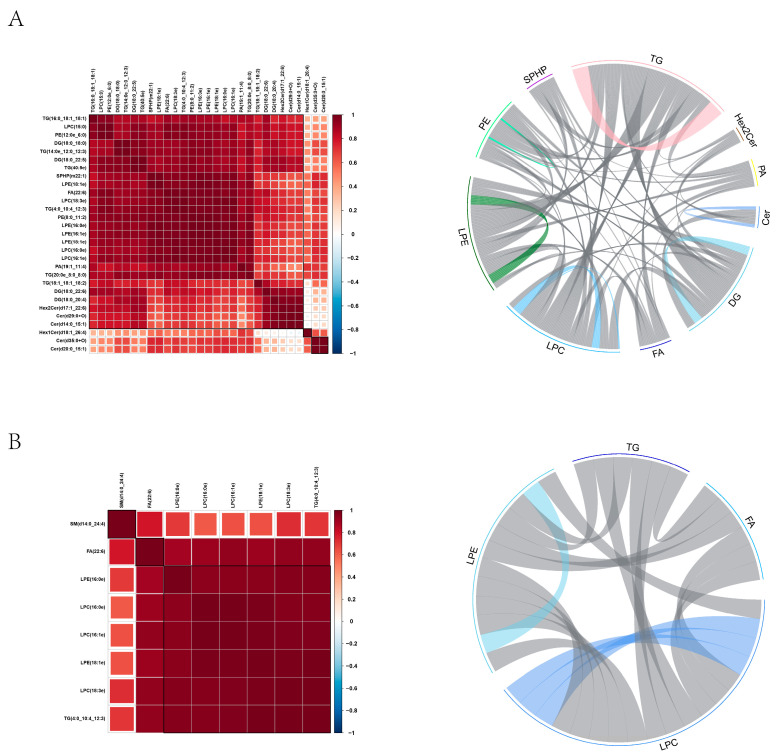 Figure 8
Figure 8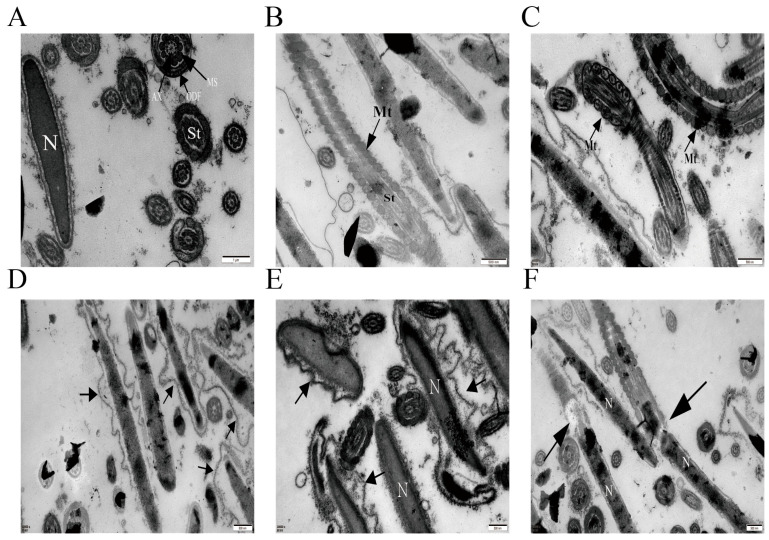 Figure 9
Figure 9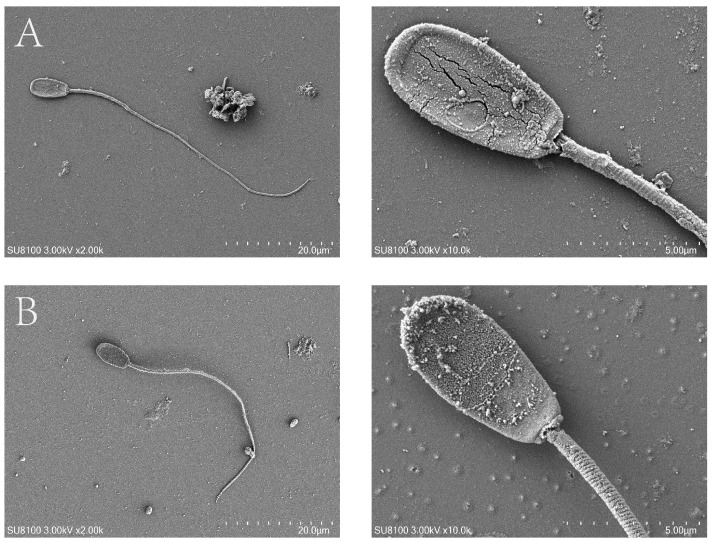 Figure 10
Figure 10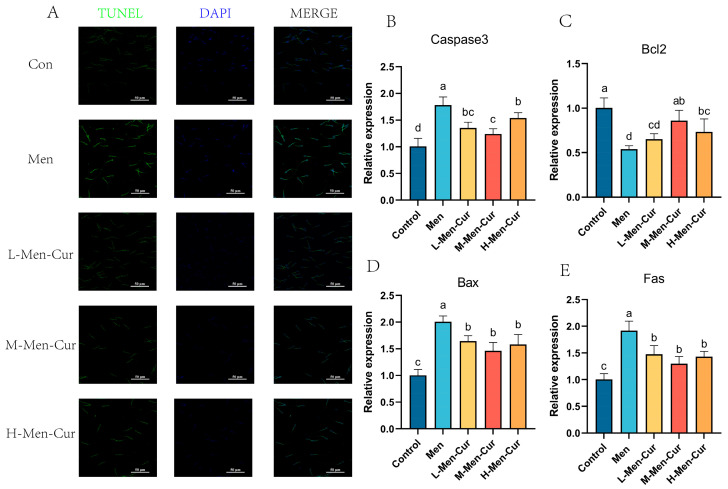 Figure 11
Figure 11- —Central Public-interest Scientific Institution Basal Research Fund
- —National Key Research and Development Program of Hainan
- —Leizhou Goat (Hainan Black Goat) Conservation Farm of Hainan
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCurcumin's Biomedical Applications · Selenium in Biological Systems · Sperm and Testicular Function
1. Introduction
There is increasing interest in understanding the role of natural antioxidants in preserving sperm quality under oxidative stress. Spermatozoa, rich in polyunsaturated fatty acids (PUFAs) and inherently deficient in antioxidant defenses, are highly susceptible to oxidative damage [1,2,3]. Oxidative stress, defined as an imbalance between reactive oxygen species (ROS) production and antioxidant capacity, has been linked to male infertility in both humans and livestock [4,5]. In livestock, this phenomenon compromises fertility, impedes breeding programs, and threatens agricultural sustainability. Environmental pollutants, nutritional deficiencies, and physiological stressors further exacerbate oxidative stress in breeding males, leading to reduced sperm viability, motility, and fertilization capacity [6].
ROS-mediated oxidative stress induces DNA damage and apoptosis in spermatozoa [7]. Apoptosis, characterized by caspase activation, mitochondrial membrane potential disruption, phosphatidylserine externalization, and DNA fragmentation, has a direct negative impact on sperm function [8,9,10]. Apoptosis directly affects the quality of sperm and exhibits typical markers of apoptosis [11]. However, the apoptotic response of goat sperm under oxidative stress remains underexplored, limiting species-specific antioxidant interventions.
Curcumin, a polyphenolic compound derived from Curcuma longa, possesses strong antioxidant properties by scavenging ROS, inhibiting lipid peroxidation, and modulating antioxidant enzyme systems [12,13,14,15]. Its high bioavailability and favorable safety profile make it an attractive candidate for reproductive health applications [16]. Curcumin’s ability to regulate cellular pathways involved in oxidative defense, inflammation, and apoptosis further supports its potential as a therapeutic agent [17]. In addition, curcumin has been shown to enhance mitochondrial activity and preserve membrane integrity, thus protecting spermatozoa from oxidative damage such as lipid peroxidation and DNA fragmentation [18,19].
Despite these promising attributes, the specific biochemical pathways through which curcumin modulates sperm metabolism and lipid profiles remain unclear, particularly in genetically distinct breeds such as the Hainan Black goat. Native to the subtropical Hainan region of China, these goats exhibit unique physiological adaptations that may influence their response to oxidative stress and antioxidant treatment [20]. Investigating the effect of curcumin in this breed may yield valuable insights into its utility for improving sperm quality and reproductive performance in livestock.
Metabolomics, lipidomics, and proteomics represent comprehensive approaches for identifying biomarkers associated with reproductive dysfunction, offering detailed insights into molecular alterations in sperm cells [21,22]. Among these, metabolomics and lipidomics provide high-resolution phenotypic snapshots by capturing biochemical activity, making them particularly valuable for studying cellular responses to environmental stressors and pathological conditions [23,24]. These methods may offer a thorough understanding of curcumin’s impact on cellular metabolism, energy dynamics, and membrane structure.
This study aimed to evaluate the effects of curcumin supplementation on semen quality, apoptosis, ultrastructure, and oxidative stress markers in sperm exposed to oxidative stress. By applying metabolomic and lipidomic profiling, we sought to elucidate the underlying biochemical pathways affected by curcumin and to identify metabolites and lipid species that may serve as biomarkers of sperm resilience. Through this integrated approach, we provide a detailed investigation into the molecular mechanisms by which curcumin enhances sperm function and counteracts oxidative damage. Our findings support curcumin’s potential as a natural antioxidant for improving sperm quality and offer critical insights into breed-specific antioxidant strategies for sustainable livestock breeding and genetic improvement.
2. Materials and Methods
2.1. Animals
Hainan Black male goats were sourced from the Genetic Resource Preservation Farm of Hainan Province (Danzhou, Hainan, China). All procedures complied with the Standards for the Protection and Utilization of Laboratory Animals of the People’s Republic of China and were approved by the Animal Care and Use Committee of Hainan University (protocol HNUAUCC-2022-000140). Bucks were clinically healthy and exhibited normal libido.
2.2. Collection and Dilution of Seminal Fluid
Twenty sexually mature Hainan Black goats (Capra hircus), aged 1.5–2 years and weighing 35.5 ± 3.2 kg, in optimal reproductive condition were selected from a standardized breeding facility. All bucks exhibited normal libido. Semen was collected using a standardized electroejaculation protocol, starting at 3 V with 5 s pulses, followed by 0.5 V increments every 30 s up to a maximum of 6 V. Collections were performed three times per week for three consecutive weeks to generate biological replicates (n = 5 per group). Fresh ejaculates were retained for further analysis if they met the following criteria: volume ≥ 0.75 mL, opalescent white coloration, absence of contamination, progressive motility ≥ 80%, sperm concentration ≥ 1.5 × 10^9^ cells/mL, and normal morphology ≥ 85%.
2.3. Preparation of Diluent and Experimental Design
All chemicals, including D-glucose (CAS 50-99-7), D-fructose (CAS 57-48-7), Tris base (CAS 77-86-1), streptomycin sulfate (CAS 3810-74-0), dimethyl sulfoxide (DMSO, CAS 67-68-5), and curcumin (≥94% purity, CAS 458-37-7), were obtained from Sorbolol (Beijing, China). Menadione (crystalline, ≥99%, CAS 58-27-5) was purchased from Sigma-Aldrich (St. Louis, MO, USA).
The base extender was formulated as a citrate-Tris buffer containing Tris-hydroxymethyl aminomethane (27 g/L), sodium citrate (13.75 g/L), fructose (10 g/L), and streptomycin sulfate (1 × 10^6^ IU/L). Curcumin stock solutions (10 mmol/L) were prepared in DMSO and diluted to final concentrations of 5, 25, and 50 μmol/L. Menadione working solution (10 mM) was freshly prepared in DMSO and diluted to 0.1 mM [25,26].
Pooled semen samples (n = 5 biological replicates) were randomly assigned to five groups through stratified randomization: (1) Control: citrate-Tris buffer + 0.1% DMSO; (2) Menadione (Men): 0.1 mM menadione; (3) L-Men-Cur: 0.1 mM menadione + 5 μmol/L curcumin; (4) M-Men-Cur: 0.1 mM menadione + 25 μmol/L curcumin; (5) H-Men-Cur: 0.1 mM menadione + 50 μmol/L curcumin. All treatments maintained a final DMSO concentration of 0.1% (v/v). After additive administration, samples were equilibrated in a 37 °C water bath (Memmert WB14, Schwabach, Germany) for 10 min, followed by a 60 min incubation under atmospheric conditions. All samples were immediately stored at −80 °C following treatment preparation for subsequent omics analyses.
2.4. Detection of Antioxidant Properties
Total antioxidant capacity (T-AOC), superoxide dismutase (SOD), glutathione peroxidase (GSH-Px), catalase (CAT), and malondialdehyde (MDA) levels were assessed using commercial kits (A015-2-1, A001-2-2, A005-1-1, A007-1-1; Nanjing Jiancheng Bioengineering Institute, Nanjing, China), according to the manufacturer’s instructions. Absorbance was measured at 593 nm, 550 nm, 412 nm, 405 nm, and 532 nm, respectively. Reactive oxygen species (ROS) were detected using the E004-1 kit (Nanjing Jiancheng). Sperm smears were fixed in 4% paraformaldehyde in PBS for 30 min and permeabilized with 1% Triton X-100/PBS for 2 h. Samples were stained with 0.1 mM DCFH-DA for 60 min and counterstained with DAPI (Roche Diagnostics, Basel, Switzerland). Fluorescence images were captured using a laser confocal microscope (LSM 900 META, Zeiss, Oberkochen, Germany).
2.5. Properties Mitochondrial Function Assays
Intracellular ATP levels were determined using an ATP assay kit (A095-1-1, Nanjing Jiancheng Bioengineering Institute, Nanjing, China), following a modified protocol. Sperm suspensions (1 × 10^7^ cells/mL) were centrifuged at 600× g for 10 min at 4 °C. Pellets were lysed with 100 μL of extraction buffer (50 mM Tris-HCl, 1 mM EDTA, 0.1% Triton X-100, pH 7.8) and reacted with a luciferase-based reagent. Luminescence was read at 636 nm using a SpectraMax i3x multimode plate reader.
Mitochondrial membrane potential (ΔΨm) was assessed using a JC-1 staining kit (C2006, Nanjing Jiancheng Bioengineering Institute, Nanjing, China). After staining and washing, fluorescence was measured at excitation/emission wavelengths of 525/590 nm (aggregates) and 488/525 nm (monomers).
2.6. Metabolite Extraction and Analysis
Samples were divided into three groups: Control, Men, and Men-Cur (25 μmol/L curcumin). After thawing on ice, 100 μL aliquots were mixed with 400 μL of methanol:acetonitrile (1:1 v/v) containing 1% isotope-labeled internal standards. The mixture was vortexed and ultrasonicated for 10 min in an ice bath, followed by protein precipitation at –40 °C for 1 h. Supernatants were collected after centrifugation (12,000× g, 15 min, 4 °C) and subjected to UPLC-MS/MS analysis (ACQUITY UPLC, Waters Corporation (Milford, MA, USA); Xevo TQ-S, Waters).
Data were processed using ProteoWizard and XCMS for peak detection and alignment. Multivariate analysis (OPLS-DA) and univariate t-tests were conducted. Metabolites with VIP > 1, p < 0.05, and FC > 1.5 or <0.67 were considered significant. Pathway enrichment was performed using KEGG.
2.7. Lipid Extraction and LC-MS Analysis
Lipid extraction involved sequential addition of water (200 μL), MTBE (800 μL), and methanol (240 μL), followed by vortexing, ultrasonication (20 min), and centrifugation (14,000× g, 10 °C, 15 min). The organic phase was dried under nitrogen, reconstituted in isopropanol/acetonitrile (90:10), centrifuged, and analyzed.
UHPLC separation was performed on a Nexera LC-30A system (Shimadzu, Kyoto, Japan) with a C18 column at 45 °C and 300 μL/min flow rate. MS detection was conducted on a QExactive™ (Thermo Scientific, Waltham, MA, USA) in positive ion mode. Raw data were analyzed using LipidSearch. Differential lipids were screened by VIP > 1, FC > 1.5 or <0.67, and p < 0.05. Pearson correlation analysis was applied to evaluate lipid associations.
2.8. Sperm Ultrastructure
Sperm pellets were fixed in 2.5% glutaraldehyde, followed by 1% osmium tetroxide. Samples were dehydrated through graded ethanol, transitioned with isoamyl acetate, and dried using a critical point dryer (Quorum K850, East Sussex, UK). After gold sputtering, samples were observed under a scanning electron microscope (Hitachi SU8100, Tokyo, Japan).
For TEM, sperm were fixed in 2% glutaraldehyde, post-fixed in 1% osmium tetroxide, dehydrated, embedded in epoxy resin, and sectioned. Ultrathin sections were stained and visualized using a JEM-1400 PLUS (JEOL Ltd., Tokyo, Japan) at 80 kV.
2.9. Evaluation of Apoptosis
Apoptotic sperm were identified using the TUNEL assay (Click-iT™ Plus, Thermo Fisher). Samples were fixed in 4% paraformaldehyde, permeabilized with 0.1% Triton X-100, and incubated with TUNEL reaction mixture at 37 °C for 60 min. Nuclei were counterstained with DAPI and visualized under a Zeiss LSM 880 confocal microscope.
2.10. Quantitative Real-Time PCR
Total RNA was extracted using TRIzol (Beyotime, Shanghai, China), and cDNA was synthesized with M-MLV reverse transcriptase. Gene expression was quantified using SYBR Green-based qPCR (CFX96, Bio-Rad, Hercules, CA, USA). Relative expression levels were calculated via the 2^−ΔΔCt^ method, normalized to β-actin. Primer sequences are listed in Table 1.
2.11. Statistical Analyses
All data are presented as mean ± SEM. Statistical analyses were performed using SPSS 20.0 (IBM, USA). One-way ANOVA followed by LSD post hoc tests was used to assess differences among groups. Significance was defined at p < 0.05. Graphs were generated using GraphPad Prism 10 (GraphPad Software, San Diego, CA, USA).
3. Results
3.1. The Specific Effects of Curcumin on the Antioxidant and Metabolic Capacity of Sperm Under Oxidative Stress
As illustrated in Figure 1, oxidative stress significantly reduced sperm T-AOC (Figure 1A), which was effectively restored by curcumin treatment. Similarly, oxidative stress markedly suppressed the activity of key antioxidant enzymes—including CAT (Figure 1B), SOD (Figure 1C) and GSH-Px (Figure 1D)—while curcumin significantly enhanced their activities. Lipid peroxidation, assessed by MDA levels (Figure 1E), increased under oxidative conditions but was significantly attenuated by curcumin. ROS accumulation, a hallmark of oxidative stress, was also significantly inhibited following curcumin treatment (Figure 1F). Furthermore, curcumin reversed oxidative stress-induced mitochondrial dysfunction by restoring mitochondrial membrane potential and ATP production, thereby supporting cellular energy homeostasis (Figure 1G,H). These findings suggest that curcumin effectively mitigates oxidative damage by enhancing both antioxidant defenses and mitochondrial function. Among the tested concentrations, the M-Men-Cur group (25 μmol/L) exhibited the most pronounced protective effect.
3.2. Multivariate Statistical Analysis and Clustering Analysis of the Metabolome
Multivariate analysis revealed distinct metabolic profiles in sperm subjected to oxidative stress and curcumin treatment. Principal component analysis (PCA) indicated high intra-group consistency, while orthogonal partial least squares discriminant analysis (OPLS-DA) confirmed significant separation between groups (Figure 2A), with robust model performance (Q^2^ > 0.5, R^2^Y > 0.9). Oxidative stress induced notable metabolic alterations, which were partially reversed by curcumin. Differential abundance analysis identified 171 altered metabolites in the oxidative stress group (117 upregulated, 54 downregulated) and 109 in the curcumin-treated group (52 upregulated, 57 downregulated) (Figure 2B). Hierarchical clustering demonstrated clear metabolic divergence between groups (Figure 2C). Cross-comparison revealed 33 overlapping differentially abundant metabolites (DAMs) exhibiting opposing expression patterns: those elevated under oxidative stress were downregulated following curcumin treatment, and those suppressed by stress were upregulated by curcumin (Figure 2D). These findings suggest that curcumin exerts targeted regulatory effects on metabolic alterations induced by oxidative stress.
3.3. Identification of Distinct Metabolites
Metabolite categorization revealed that lipids and lipid-like molecules, along with organic acids and derivatives, were the predominant classes of differentially abundant metabolites in both the oxidative stress and curcumin-treated groups (Figure 3A,B). The top 20 metabolites, ranked by VIP scores, demonstrated strong discriminatory power between groups (Figure 3C,D). These metabolites were further prioritized based on their log_2_(fold change) (log_2_FC) values to characterize their expression patterns (Figure 3E,F). Notably, chlorpromazine—a compound associated with pro-oxidative effects—was elevated under oxidative stress conditions. In contrast, metabolites such as nobiletin, glabridin, docosapentaenoic acid, and ketotifen, known for their antioxidant properties, were significantly upregulated following curcumin treatment.
3.4. Analysis of Differential Correlations Among Metabolites
Spearman correlation analysis was conducted on the top 50 metabolites with the highest VIP scores. Additionally, direct correlation analysis was performed to compare metabolite relationships between the oxidative stress and curcumin-treated groups. The correlation coefficient ranged from 1.0 (indicating a strong positive correlation) to −1.0 (indicating a strong negative correlation), with 0 representing no correlation. Gentisic acid exhibited a strong negative association with Cys-Ala (R = −0.95, p < 0.01), and strong positive correlations with Chamanetin (R = 0.92, p < 0.01) and Glyoxal (R = 0.80, p < 0.01) (Figure 4A). A strong positive correlation was also observed between 3-hydroxybutyric acid and Oxodecanoylcarnitine (R = 0.94, p < 0.01), as well as between L-2-amino-3-oxybutyric acid and L-homoserine lactone (R = 0.83, p < 0.01). Glucosyl acid showed a strong negative correlation with L-homoserine lactone (R = −0.95, p < 0.01), as well as strong positive correlations with Oxodecanoylcarnitine (R = 0.81, p < 0.01) and 3-hydroxybutyric acid (R = 0.85, p < 0.01) (Figure 4B).
3.5. The Significance of Distinct Metabolites Within Metabolic Pathways
Metabolic pathway analysis was conducted using the KEGG database to assess the involvement of differential metabolites in specific biochemical pathways. Several differential metabolites were enriched in pathways such as the citric acid cycle (TCA cycle), pentose and gluconate interconversion, arachidonic acid metabolism, vitamin B6 metabolism, the HIF-1 signaling pathway, and the cAMP signaling pathway, which are potentially important in the context of oxidative stress (Figure 5A). In the curcumin-treated group, pathways such as histidine metabolism, the pentose phosphate pathway, glycine/serine/threonine metabolism, sphingolipid metabolism, glyoxylate and dicarboxylate metabolism, and the cAMP signaling pathway were implicated in curcumin-mediated metabolic regulation (Figure 5B). Distinct metabolites were identified within these pathways, with notable examples including gluconic acid, 3-hydroxybutyric acid, and argininosuccinic acid. Relative content analysis and ROC curve evaluation were performed for these three metabolites. All exhibited AUC values greater than 0.8, suggesting strong predictive value for distinguishing semen status following curcumin treatment under oxidative stress (Figure 5C).
3.6. Multivariate Analysis of Lipid Profiles
To distinguish lipidomic differences between groups, an OPLS-DA model was applied. The results showed that the oxidative stress and curcumin-treated groups were distributed on opposite sides of the central axis, indicating clear separation between lipid profiles (Figure 6A). The model’s reliability and predictive power were supported by high R^2^Y and Q^2^ values. Permutation testing confirmed the absence of overfitting, as indicated by the negative intercept of the Q^2^ regression line on the Y-axis (Figure 6B). Major lipid classes were identified and quantified, with phosphatidylcholine (PC) and phosphatidylethanolamine (PE) showing higher relative abundance in both the oxidative stress and curcumin-treated groups, followed by acylcarnitine (AcCa) (Figure 6C). Additionally, the overall lipid composition was quantified in each group to assess curcumin’s influence on lipid regulation (Figure 6D,E).
3.7. Screening of Differential Lipid Molecules
Differential lipid analysis was conducted based on criteria of fold change (FC > 1.5 or FC < 0.67), VIP > 1, and p < 0.05. The screened lipids are listed in Table 2 and Table 3. Volcano plots (Figure 7A,B) highlighted lipid molecules that met the selection criteria. Under oxidative stress, key differential lipids included triglycerides (TG), lysophosphatidylcholine (LPC), and lysophosphatidylethanolamine (LPE) (Figure 7C). In contrast, curcumin treatment primarily altered fatty acids (FA) and LPCs (Figure 7D). Oxidative stress led to increased lipid accumulation (Figure 7E), whereas curcumin treatment significantly reduced lipid content (Figure 7F).
3.8. Correlation Between Different Lipids
To evaluate coordinated lipid responses, Pearson correlation analysis was performed. In the oxidative stress group, TG levels were positively correlated with diacylglycerol (DG), LPE, and LPC (Figure 8A), suggesting a shared lipid regulatory pattern under stress conditions. In the curcumin-treated group, LPC was positively correlated with LPE, FA, and TG, indicating lipid co-regulation during curcumin-mediated recovery (Figure 8B).
3.9. Effect of Curcumin on Sperm Ultrastructure Under Oxidative Stress
Transmission electron microscopy (TEM) revealed that curcumin preserved the ultrastructure of spermatozoa. Treated sperm exhibited intact plasma membranes closely apposed to the nuclear membrane, homogeneous chromatin distribution, and complete acrosomes with continuous membranes (Figure 9A). Mitochondria appeared structurally intact, with distinct inner and outer membranes and well-defined intermembrane spaces (Figure 9B,C). In contrast, oxidative stress induced apoptotic features including plasma membrane blebbing (Figure 9D,E), mitochondrial swelling, vacuolation, deformation, and loss of cristae (Figure 9F). Scanning electron microscopy (SEM) showed oxidative stress-related head deformities such as acrosomal disruption, flattening, or rupture (Figure 10A), which were significantly ameliorated by curcumin (Figure 10B).
3.10. Curcumin Alleviates Sperm Apoptosis Under Oxidative Stress
TUNEL staining demonstrated that curcumin significantly reduced sperm apoptosis rates compared to the oxidative stress group (Figure 11A). qPCR analysis showed that expression levels of pro-apoptotic genes (Fas, Bax, Caspase3) were significantly decreased, while anti-apoptotic Bcl2 expression was upregulated following curcumin treatment (Figure 11B–E). These findings indicate that curcumin mitigates oxidative stress-induced apoptosis in Hainan Black goat sperm, with the most pronounced effect observed at 25 μmol/L (M-Men-Cur).
4. Discussion
This study demonstrates that curcumin exerts significant protective effects against oxidative stress-induced damage in Hainan Black goat sperm. Through enhancement of antioxidant defense systems, curcumin improved semen quality, reduced apoptosis, and preserved sperm ultrastructure. Metabolomic analyses revealed that curcumin modulates key metabolic pathways related to energy production and oxidative defense, with gluconic acid, 3-hydroxybutyric acid, and argininosuccinic acid identified as central metabolites.
4.1. Effects of Curcumin on Sperm Metabolism of Hainan Black Sheep Under Oxidative Stress
Spermatozoa primarily rely on glucose metabolism via the pentose phosphate pathway (PPP) to generate NADPH and maintain redox homeostasis [27]. Elevated gluconic acid levels, a byproduct of glucose oxidation, have been implicated in promoting oxidative stress [28]. The elevation of gluconic acid following curcumin treatment suggests activation of the PPP to sustain ATP levels and sperm function [29]. Concurrently, NADPH derived from the PPP serves as a cofactor for regenerating reduced glutathione (GSH) from its oxidized form (GSSG), bolstering antioxidant defenses against oxidative damage [30,31]. Furthermore, 3-hydroxybutyric acid, elevated in curcumin-treated groups, enhances mitochondrial membrane potential, reduces DNA fragmentation, and improves mitochondrial resilience against oxidative stress [32,33]. Curcumin also restored argininosuccinic acid levels—a metabolite depleted in malformed sperm—surpassing concentrations observed in the oxidative stress group [34]. Collectively, these metabolic shifts underscore curcumin’s role in rebalancing energy metabolism, antioxidant activity, and structural integrity in stressed sperm.
4.2. Effects of Curcumin on Sperm Lipids of Hainan Black Sheep Under Oxidative Stress
Lipidomic analyses revealed pronounced remodeling of sperm lipid profiles following curcumin treatment. Oxidative stress induced a marked accumulation of lysophosphatidylcholines (e.g., LPC(16:0e), LPC(16:1e), LPC(18:3e)), a signature of membrane disruption linked to impaired sperm integrity [35]. Curcumin attenuated this LPC enrichment, suggesting preservation of membrane architecture. Triglycerides (TGs), important energy stores for spermatogenesis [36], were elevated under oxidative stress. Although physiological TG levels can correlate positively with motility in certain species (e.g., white salmon) [37], pathological TG accumulation—reported here and in diabetic rodent models—perturbs membrane dynamics and compromises motility [38]. Consistently, Slc22a14-deficient mice exhibit TG-driven metabolic dysfunction with reduced β-oxidation and depleted tricarboxylic-acid (TCA) intermediates [39]. Curcumin normalized TG levels, likely mitigating these adverse effects.
Sperm membranes have an unusual lipid composition with a high contribution of sphingolipids [40]. In rat sperm heads, sphingomyelin (SM) can be converted to ceramide during the acrosome reaction, facilitating membrane reorganization [41]. However, higher SM content has been observed in stallion sperm with poor freezing resistance and correlates negatively with motility and membrane integrity [42]. Curcumin reduced SM abundance, a change potentially stabilizing the membrane and limiting stress-induced remodeling. Overall, curcumin counters oxidative injury by restoring lipid homeostasis: lowering peroxidation-prone LPCs, normalizing energy-critical TGs, and modulating SM-dependent membrane dynamics.
4.3. Effect of Curcumin on Oxidative Stress-Induced Sperm Apoptosis of Hainan Black Goat
Apoptosis is a major determinant of sperm quality, affecting motility, membrane integrity, and acrosomal function [43,44]. Notably, cryo-induced apoptosis negatively correlates with motility and plasma membrane integrity during sperm preservation [45,46]. In this study, apoptosis in goat sperm exposed to oxidative stress—with or without curcumin—was evaluated by TUNEL in combination with DAPI staining, a well-established approach in andrology research [47]. Curcumin markedly reduced the proportion of apoptotic sperm, underscoring apoptosis as a sensitive indicator of oxidative damage in goat semen.
Ultrastructural analyses revealed canonical features of apoptosis, including membrane blebbing, apoptotic body formation, nuclear envelope defects, and nuclear fragmentation, mirroring observations in human sperm [48]. Particularly prominent were mitochondrial alterations, consistent with mitochondria-dependent apoptotic signaling [49,50]. Oxidative stress elevated endogenous ROS and malondialdehyde (MDA) while lowering the activities of SOD, CAT, and GSH-Px, indicating both excessive ROS generation and diminished antioxidant performance. Elevated ROS drives lipid peroxidation, compromises membrane quality, and precipitates loss of mitochondrial membrane potential (MMP), cytochrome-c release, and caspase activation—hallmarks of mitochondria-mediated apoptosis [51,52,53,54,55,56]. In contrast, curcumin, acting as an effective ROS scavenger, curtailed ROS accumulation, improved antioxidant enzyme status, restored MMP, and ultimately restrained apoptosis. Collectively, these data support a model in which oxidative stress triggers ROS-driven mitochondrial damage and mitochondria-dependent apoptosis in goat spermatozoa, while curcumin interrupts this cascade by re-establishing redox balance and mitochondrial function.
5. Conclusions
Curcumin (25 μmol/L) exhibits significant protective effects against oxidative stress-induced damage in Hainan Black goat sperm. It enhances antioxidant capacity, preserves mitochondrial function, restores membrane integrity, and reduces apoptosis. Integrated metabolomic and lipidomic analyses revealed that curcumin modulates critical pathways involved in energy metabolism, redox balance, and lipid homeostasis, with gluconic acid, 3-hydroxybutyric acid, and argininosuccinic acid identified as key biomarkers. These findings provide mechanistic insights into curcumin’s role in mitigating oxidative damage and highlight its potential as a natural antioxidant for improving semen quality and fertility in livestock breeding programs.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aitken R.J. Smith T.B. Jobling M.S. Baker M.A. De Iuliis G.N. Oxidative stress and male reproductive health Asian J. Androl.201416313810.4103/1008-682X.12220324369131 PMC 3901879 · doi ↗ · pubmed ↗
- 2Sanocka D. Kurpisz M. Reactive oxygen species and sperm cells Reprod. Biol. Endocrinol. RB&E 200421210.1186/1477-7827-2-1215038829 PMC 400757 · doi ↗ · pubmed ↗
- 3Aitken R.J. Roman S.D. Antioxidant systems and oxidative stress in the testes Oxidative Med. Cell. Longev.20081152410.4161/oxim.1.1.684319794904 PMC 2715191 · doi ↗ · pubmed ↗
- 4Darbandi M. Darbandi S. Agarwal A. Sengupta P. Durairajanayagam D. Henkel R. Sadeghi M.R. Reactive oxygen species and male reproductive hormones Reprod. Biol. Endocrinol. RB&E 2018168710.1186/s 12958-018-0406-230205828 PMC 6134507 · doi ↗ · pubmed ↗
- 5Marín R. Abad C. Rojas D. Chiarello D.I. Alejandro T.G. Biomarkers of oxidative stress and reproductive complications Adv. Clin. Chem.202311315723310.1016/bs.acc.2022.11.00436858646 · doi ↗ · pubmed ↗
- 6Bansal A.K. Bilaspuri G.S. Impacts of oxidative stress and antioxidants on semen functions Vet. Med. Int.2010201068613710.4061/2011/68613720871827 PMC 2943128 · doi ↗ · pubmed ↗
- 7Sharma P. Kaushal N. Saleth L.R. Ghavami S. Dhingra S. Kaur P. Oxidative stress-induced apoptosis and autophagy: Balancing the contrary forces in spermatogenesis Biochim. Et Biophys. Acta Mol. Basis Dis.2023186916674210.1016/j.bbadis.2023.16674237146914 · doi ↗ · pubmed ↗
- 8Barroso G. Morshedi M. Oehninger S. Analysis of DNA fragmentation, plasma membrane translocation of phosphatidylserine and oxidative stress in human spermatozoa Hum. Reprod.2000151338134410.1093/humrep/15.6.133810831565 · doi ↗ · pubmed ↗