Looking for ESKAPE Bacteria: Occurrence and Phenotypic Antimicrobial Resistance Profiles in Wild Birds from Northern and Central Italy Sites
Guido Grilli, Maria Cristina Rapi, Laura Musa, Giacomo Di Giacinto, Fabrizio Passamonti, Stefano Raimondi, Oriana Cianca, Maria Pia Franciosini

TL;DR
This study found that wild birds in Italy carry bacteria known for antimicrobial resistance, including some that are dangerous to humans.
Contribution
The study provides new insights into the occurrence and resistance profiles of ESKAPE bacteria in wild birds in Italy.
Findings
Thirty-seven ESKAPE bacteria isolates were identified from 141 wild birds.
Multidrug-resistant isolates were found among Klebsiella pneumoniae and Enterobacter hormaechei.
Escherichia coli was frequently detected and often co-isolated with clinically relevant bacteria.
Abstract
Background/Objectives: Antimicrobial resistance is a critical global health challenge. Among resistant pathogens, the group of bacteria collectively referred to as ESKAPE (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp.) is of particular concern due to their ability to evade multiple classes of antimicrobials. This study aimed to investigate the occurrence and resistance patterns of ESKAPE bacteria in wild birds from Northern and Central Italy sites, and to assess the presence of other bacteria of public health relevance. Methods: Cloacal swabs were collected from 141 wild birds. Samples were processed on selective and differential media, and bacterial identification was performed using Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry. Antimicrobial susceptibility was…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
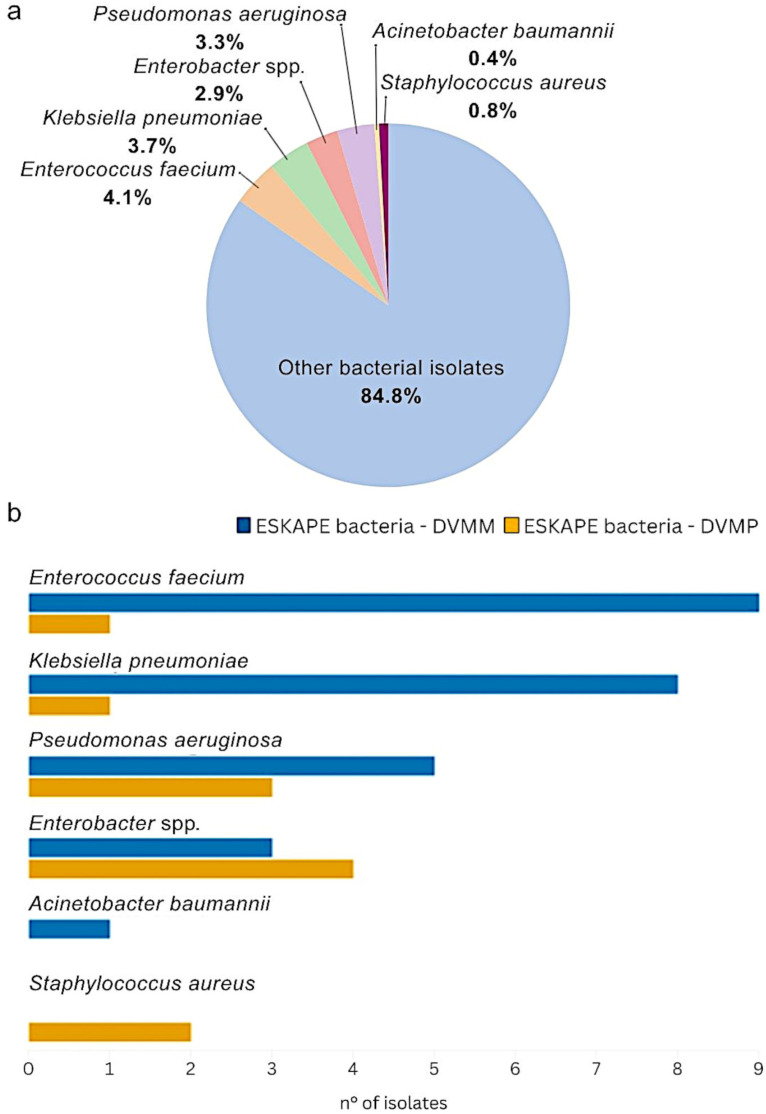 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Pharmaceutical and Antibiotic Environmental Impacts · Bacterial Identification and Susceptibility Testing
1. Introduction
Antimicrobial resistance (AMR) is currently one of the most significant public health challenges, with substantial economic and social implications [1,2,3]. The selective pressure resulting from the prolonged use of antimicrobials in both human and veterinary medicine has driven the emergence of multidrug-resistant (MDR) pathogens capable of evading multiple classes of antimicrobial agents [4,5,6]. Consequently, continued research into innovative therapeutic strategies to combat AMR is essential. In this scenario, the ESKAPE bacteria (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp.) are a source of concern [7,8]. These bacteria are a leading cause of mortality among immunocompromised subjects, especially in hospital and healthcare settings, due to their resistance not only to first-line antibiotics, but also to critically important antimicrobials (CIAs) in human medicine [7,8,9]. ESKAPE are not confined to clinical environments; they have also been detected in soil, surface water, and wastewater [10]. Their persistence in different habitats and ability to engage in horizontal gene transfer (HGT) allow them to acquire resistance through interactions with other bacteria [11]. In this respect, wildlife may serve as a link in the transmission cycle of AMR bacteria among humans, domestic animals, and the environment [12,13].
Although wild animals are not typically exposed to antimicrobials directly, their extensive use in human and veterinary medicine indirectly has driven the emergence of antimicrobial-resistant bacteria (ARB) in free-ranging species [14,15,16]. As ARB are generally present in natural ecosystems due to anthropogenic contamination, wild species inhabiting urban, agricultural, or landfill areas, or frequently interacting with humans and domestic animals, are more likely to carry ARB than those from less disturbed habitats [14,15,17,18,19]. Based on this, it is essential to identify wildlife species whose behavioral, ecological, and feeding characteristics make them suitable “sentinels” for assessing the extent of AMR in the environment [14,20]. Given their broad geographic distribution and high mobility, birds are acknowledged as effective bioindicators [17,21,22,23], capable of reflecting the presence and environmental dissemination of ARB potentially pathogenic to humans, ESKAPE group included [14,24,25]. Nevertheless, wild avian species are rarely included in AMR monitoring programs. Numerous studies have reported a high prevalence of AMR bacteria in bird populations inhabiting areas close to human settlements and with intensive livestock production in different geographical contexts [14]; however, considering the ESKAPE group, most of the available epidemiological investigations on wild birds have focused on single ESKAPE species or genera, while comprehensive assessments of the entire group remain limited [25]. Recent research has examined S. aureus in wild birds, characterizing resistance and virulence determinants [26], and Enterococcus spp. (including E. faecium) through combined phenotypic and genomic approaches [27]. Similarly, different studies have investigated the occurrence and MDR resistance profile, as well as Extended Spectrum Beta-Lactamases (ESBLs) production in K. pneumoniae isolates from different bird species and categories [28,29]. More recently, Silva et al. [25] reviewed extensively the diffusion of ESKAPE in wildlife, including avifauna, in several countries, highlighting the presence of resistant strains circulating among animals, men and the environment. In Italy, only one study conducted in the South by Russo et al. [12] specifically investigated the occurrence and resistance profiles of all bacterial species belonging to the ESKAPE group in avifauna, reporting that 24.5% of 163 birds sampled carried ESKAPE bacteria with variable AMR patterns, including resistance towards CIAs. To the best of our knowledge, no comprehensive study has yet investigated the simultaneous occurrence of all ESKAPE members in wild birds in Northern and Central Italy.
Considering this background, this study, carried out through collaboration between the Department of Veterinary Medicine, University of Perugia (DVMP, Umbria (Central Italy), and the Department of Veterinary Medicine and Animal Science, University of Milan (DVMM), Lombardy (Northern Italy), aimed to investigate, although in preliminary manner, the occurrence and phenotypic AMR profiles of ESKAPE bacteria isolated from wild birds. In addition, the presence of other bacteria of emerging relevance for public health [30,31,32] was documented. The detection of ESKAPE bacteria in wildlife carries important implications for zoonotic transmission and potential risks to human health. Although most research has focused on livestock and domestic animals as sources of clinically relevant resistant bacteria, wildlife constitutes a less monitored but equally relevant reservoir [25]. By providing novel data on the presence and resistance patterns of ESKAPE bacteria in wild avian species from Northern and Central Italy sites, this study provides additional data to help fill existing knowledge gaps and offers further insights into the potential role of wild birds in the clinically relevant ARB and AMR ecology. Furthermore, it highlights the importance of including wildlife in One Health-based surveillance frameworks to enable early detection of emerging resistant and pathogenic bacteria and to identify environmental hotspots before direct human exposure occurs [24].
2. Results
2.1. Sampled Bird Population
A total of 141 wild birds were sampled, with 66 (46.8%) from DVMM, and 75 (53.2%) from DVMP.
The investigated birds represented a broad taxonomic spectrum, including 15 orders, 22 families, and 42 species, comprising both resident and migratory species, as reported in Table 1. Overall, the most represented orders were Passeriformes (n = 45/141; 31.9%) and Strigiformes (n = 27/141; 19.1%), which together accounted for over 51% of all sampled birds. Other orders included Columbiformes (n = 16/141; 11.3%), Apodiformes (n = 12/141; 8.5%), Falconiformes (n = 10/141; 7.1%), Accipitriformes (n = 8/141; 5.7%), and Anseriformes (n = 8/141; 5.7%).
When comparing the two sites, differences emerged in species composition, as also illustrated in Table 1 and in Supplementary Materials (Figure S1). DVMM had a higher representation of some Anseriformes species, particularly Mallard (Anas platyrhynchos), and Apodiformes species, represented by Common Swift (Apus apus) and Alpine Swift (Tachymarptis melba). On the other side, DVMP showed a greater number of Strigiformes, in particular Tawny Owl (Strix aluco). Similarly, diurnal raptors, Accipitriformes and Falconiformes, were more frequently recorded in Central Italy area.
2.2. Bacterial Isolation and Identification
2.2.1. ESKAPE Bacteria Isolation in Both DVMM and DVMP
Out of 141 total cloacal swabs, 243 isolates were obtained and identified. Among these, 37 out of 243 (15.2%) were classified as ESKAPE bacteria, comprising 10 (4.1%) E. faecium, 9 (3.7%) K. pneumoniae, 8 (3.3%) P. aeruginosa, 7 (2.9%) Enterobacter spp., 2 (0.8%) S. aureus, and only 1 (0.4%) A. baumannii, as reported in Figure 1a. Of these, a total of 11 (29.7%) ESKAPE isolates were identified at the DVMP, while 26 (70%) were identified at the DVMM.
2.2.2. Distribution of ESKAPE Bacteria and Their Co-Occurrence with Other Bacterial Isolates at DVMM
Among the ESKAPE isolates collected at DVMM, the most frequently identified were E. faecium (n = 9/26; 34.6%) and K. pneumoniae (n = 8/26; 30.8%), followed by P. aeruginosa (n = 5/26; 19.2%) and Enterobacter spp. (n = 3/26; 11.5%), as represented in Figure 1b. A single A. baumannii (3.8%) was isolated from a Eurasian Magpie (Pica pica). In several cases, ESKAPE bacteria isolated from DVMM samples were found in association with other relevant bacteria, as shown in Supplementary Materials (Table S1). Most K. pneumoniae isolates identified at DVMM were co-isolated with Escherichia coli and Proteus mirabilis, particularly in Common Swifts (Apus apus) and Common Blackbirds (Turdus merula). Two additional K. pneumoniae isolates were recovered alongside E. faecium: 1 from an Alpine Swift (Tachymarptis melba) and the other from a Common Swift (Apus apus). Pseudomonas aeruginosa was detected in association with P. mirabilis and Klebsiella aerogenes in a Common Blackbird (Turdus merula). Considering Enterobacter spp., notable co-isolations were observed, including Enterobacter hormaechei isolated with Enterococcus faecalis and Staphylococcus saprophyticus in a Little Egret (Egretta garzetta), and Enterobacter roggenkampii with E. coli in a European Greenfinch (Chloris chloris). Finally, the single A. baumannii isolate from DVMM was recovered together with Klebsiella oxytoca and E. coli in a Eurasian Magpie (Pica Pica).
2.2.3. Distribution of ESKAPE Bacteria and Their Co-Occurrence with Other Bacterial Isolates at DVMP
Concerning DVMP samplings, among the 11 ESKAPE isolates obtained, the most frequently identified were Enterobacter spp. (n = 4/11; 36.4%), followed by P. aeruginosa (n = 3/11; 27.3%), S. aureus (n = 2/11; 18.2%), K. pneumoniae (n = 1/11; 9.1%), and E. faecium (n = 1/11; 9.1%), as reported in Figure 1b. Specifically, as shown in Supplementary Materials (Table S2), 4 E. hormaechei isolates were obtained: from a Mallard (Anas platyrhynchos) co-isolated with P. aeruginosa; from a Cormorant (Phalacrocorax carbo) with E. coli; from a European Starling (Sturnus vulgaris) with Staphylococcus xylosus; and from a Eurasian Hobby (Falco subbuteo) as a single isolate.
Pseudomonas aeruginosa was detected in a Eurasian Collared-Dove (Streptopelia decaocto), in a Short-toed Snake Eagle (Circaetus gallicus) alongside Mammaliicoccus sciuri and E. faecium, and in a Mallard (Anas platyrhynchos) in association with E. hormaechei. Of the 2 S. aureus isolates, 1 was recovered from a Common Blackbird (Turdus merula), while the other was identified in a Tawny Owl (Strix aluco) together with S. xylosus and E. coli. Additionally, the single isolates of E. faecium and K. pneumoniae were obtained from a Common Swift (Apus apus) and a Eurasian Goshawk (Accipiter gentilis), respectively (Table S2).
2.2.4. Non-ESKAPE Bacteria Isolation in Both DVMM and DVMP
As reported in Supplementary Materials (Tables S1 and S2) among non-ESKAPE bacteria, E. coli was the most prevalent species at both sampling sites, detected in 38 of 66 samples (57.6%) at DVMM and in 46 of 75 samples (61.3%) at DVMP. Proteus mirabilis was also frequently isolated in DVMM (n = 24/66; 36.4%), while E. faecalis was the most common Gram-positive bacterium (n = 33/66; 50%). Additional species of public health concern included Citrobacter freundii, Citrobacter braakii, and Hafnia alvei, the latter isolated from a Little Owl (Athene noctua) and from a Barn Swallow (Hirundo rustica). In DVMP Escherichia marmotae was also detected in several bird species. The 2 isolates of Salmonella Typhimurium were obtained from a Common Moorhen (Gallinula chloropus), and from a Blue Tit (Cyanistes caeruleus) associated with S. saprophyticus. Additionally, K. oxytoca and K. aerogenes were detected in both DVMP and DVMM (Supplementary Materials, Tables S1 and S2).
2.3. Salmonella Serotyping
At the DVMP site, two Salmonella isolates were recovered: one from a Common Moorhen (Gallinula chloropus) and one from a Blue Tit (Cyanistes caeruleus). Both isolates were identified as Salmonella enterica subsp. enterica serovar Typhimurium, according to the Kaufmann–White–Le Minor scheme. No Salmonella isolates were obtained from DVMM samples.
2.4. AMR Profiles of ESKAPE Bacteria by Minimun Inhibitory Concentration (MIC) Determination
The AMR profiles of Gram-negative and Gram-positive ESKAPE bacteria isolated from DVMM and DVMP are summarized in Table 2 and Table 3, respectively.
2.4.1. MIC Results for Gram-Negative ESKAPE Bacteria Isolated at DVMM
The AMR profile of Gram-negative ESKAPE isolates from DVMM are shown in Table 2. In details all E. hormaechei isolates were susceptible to the tested agents, whereas the single E. roggenkampii isolate, recovered from a European Greenfinch (Chloris chloris), was resistant to enrofloxacin, florfenicol, flumequine, sulfisoxazole, trimethoprim/sulfamethoxazole, and tetracycline. Among the K. pneumoniae isolates, resistance patterns varied: 4 out of 8 (50%) were fully susceptible to the tested antimicrobials, while the remaining isolates showed resistance to flumequine (n = 4/8; 50%), enrofloxacin (n = 4/8; 50%), cefazolin (n = 3/8; 37.5%), cefotaxime (n = 3/8; 37.5%), sulfisoxazole (n = 3/8; 37.5%), tetracycline (n = 3/8; 37.5%), amoxicillin/clavulanic acid (n = 2/8; 25%), and trimethoprim/sulfamethoxazole (n = 2/8; 25%). Resistance to aminosidine and gentamicin was also observed in some isolates. The single A. baumannii isolate, recovered from a Eurasian Magpie (Pica pica), was resistant to gentamicin but remained susceptible to trimethoprim/sulfamethoxazole, enrofloxacin and colistin. For several other antimicrobials, no validated clinical breakpoints are currently available for this species, and therefore MIC values could not be interpreted. Pseudomonas aeruginosa isolates were susceptible to colistin (n = 5/5; 100%), florfenicol (n = 5/5; 100%), and gentamicin (n = 5/5; 100%), while only 1 isolate (n = 1/5; 20%) resulted susceptible to enrofloxacin. Nonetheless, elevated MIC values for aminosidine and sulfisoxazole were observed in several isolates.
2.4.2. MIC Results for Gram-Negative ESKAPE Bacteria Isolated at DVMP
With regard to the DVMP Gram-negative ESKAPE bacteria (Table 2), the single K. pneumoniae isolate, obtained from a Common Swift (Apus apus), was susceptible to all tested antimicrobials except cefazolin, for which intermediate susceptibility was observed. The E. hormaechei isolates exhibited high levels of resistance to enrofloxacin (n = 3/4; 75%), florfenicol (n = 3/4; 75%), sulfisoxazole (n = 3/4; 75%), flumequine (n = 3/4; 75%), trimethoprim/sulfamethoxazole (n = 3/4; 75%), and to third-generation cephalosporins (n = 2/4; 50%).
Similarly, to the DVMM site, P. aeruginosa isolates from DVMP exhibited elevated MIC values for sulfisoxazole (>512 µg/mL) and aminosidine (>32 µg/mL), yet remained susceptible to colistin, gentamicin, and florfenicol. Only 1 of the 3 (33.3%) isolated strain resulted resistant to enrofloxacin, while the other 2 presented reduced susceptibility to this molecule.
2.4.3. MIC Results for Gram-Positive ESKAPE Bacteria Isolated at DVMM
Regarding Gram-positive ESKAPE bacteria from DVMM (Table 3), E. faecium isolates were uniformly susceptible to florfenicol (n = 9/9; 100%). Resistance was observed to enrofloxacin (n = 4/9; 44.4%), tetracycline (n = 3/9; 33.3%), erythromycin (n = 2/9; 22.2%), ampicillin (n = 1/9; 11.1%), amoxicillin/clavulanic acid (n = 1/9; 11.1%), and penicillin (n = 1/9; 11.1%).
2.4.4. MIC Results for Gram-Positive ESKAPE Bacteria Isolated at DVMP
Among Gram-positive ESKAPE isolates from DVMP, both S. aureus isolates were resistant to sulfisoxazole and clindamycin, with 1 isolate also resistant to tetracycline. The single E. faecium isolate was resistant only to tetracycline, but susceptible to all other tested antimicrobials for which interpretation was feasible, as reported in Table 3.
2.5. MDR Profile Among ESKAPE Bacteria
Of the 37 ESKAPE bacteria tested, 10 (27%) exhibited a MDR profile. Among these, 80% (n = 8/10) were Gram-negative, with 5 isolates from DVMM (62.5%) and 3 from DVMP (37.5%), while the remaining 20% (n = 2/10) were Gram-positive, one from each site.
2.5.1. MDR in Gram-Negative ESKAPE Bacteria Isolated at Both Sites
Among the Gram-negative isolates from DVMM, MDR was observed in 5 cases: the single E. roggenkampii isolate and 4 out of 8 (50%) Klebsiella pneumoniae isolates recovered from a Common Redstart (Phoenicurus phoenicurus), European Pied Flycatcher (Ficedula hypoleuca), Alpine Swift (Tachymarptis melba), and Common Blackbird (Turdus merula). The MDR K. pneumoniae isolates were resistant to different combinations of beta-lactams, fluoroquinolones, tetracycline, and sulphonamides.
At DVMP, 3 of the 4 (75%) E. hormaechei isolates, obtained from a Mallard (Anas platyrhynchos), a Cormorant (Phalacrocorax carbo), and a Eurasian Hobby (Falco subbuteo), displayed MDR phenotypes, with resistance to third-generation cephalosporins, fluoroquinolones, phenicols, sulphonamides, and tetracyclines.
2.5.2. MDR in Gram-Positive ESKAPE Bacteria Isolated at Both Sites
Regarding Gram-positive ESKAPE bacteria, MDR was less frequent. At DVMM, a single E. faecium isolate from a Common Woodpigeon (Columba palumbus) exhibited resistance to penicillins, macrolides, and tetracycline.
At DVMP, one Staphylococcus aureus isolate from a Common Blackbird (Turdus merula) showed resistance to lincosamides, sulphonamides, and tetracycline, thus fulfilling the MDR definition.
3. Discussion
Antimicrobial resistance is recognized as a serious public health threat and has been described as “an ecological problem” [35], involving complex interactions among different microbial populations that influence the health of animals, humans, and the environment [10]. The World Health Organization (WHO) has compiled a list of critical bacteria, identifying the ESKAPE bacteria as those for which urgent actions are required considering that, alongside E. coli, they are increasingly associated with MDR profile [36,37]. In this context, the present study aimed to provide preliminary insights into the potential role of wild birds in the dissemination of ESKAPE bacteria and to characterize their phenotypic AMR profiles.
In our study differences were observed among wild bird species sampled in the two sites: DVMM had a higher number of Anseriformes and Apodiformes, particularly Mallard (Anas platyrhynchos), Common Swift (Apus apus), and Alpine Swift (Tachymarptis melba) respectively, while DVMP showed a greater number of Strigiformes, such as Tawny Owl (Strix aluco) and Little Owl (Athene noctua), as well as diurnal raptors (Accipitriformes). This regional variation likely reflects differences in local habitat structure, landscape composition, and anthropogenic pressures between Lombardy’s peri-urban environment and Umbria’s more internal rural context, emphasizing the complementary ecological contributions of both sites to the study.
Among the ESKAPE bacteria, a total of 7 Enterobacter spp. isolates were recovered across both sampling sites. Six were identified as E. hormaechei, of which 4 were collected at DVMP (n = 4/75; 5.3%) and 2 at DVMM (n = 2/66; 3.0%), and 1 E. roggenkampii at DVMM (n = 1/66; 1.5%) from a European Greenfinch (Chloris chloris).
Enterobacter spp. are generally considered part of the intestinal microbiota of both humans and animals, where they are typically non-pathogenic [38,39]. Nonetheless, E. hormaechei and E. roggenkampii are members of the Enterobacter cloacae complex (ECC), a cluster of closely related species characterized by their potential to cause disease and their capacity to acquire and spread antimicrobial resistance genes (ARGs) [40,41]. Enterobacter hormaechei, in particular, is regarded as one of the most clinically significant species within the ECC, owing genomic islands associated with high pathogenicity [39]. Its public health importance has increased in recent years due to its capacity to disseminate ESBLs and carbapenemases [42], and the WHO has listed E. hormaechei among critically important pathogens [43,44,45,46,47]. In our study the co-occurrence of E. hormaechei with other relevant Gram-negative bacteria, such as E. coli, P. aeruginosa, and K. pneumoniae noticed in both DVMP and DVMM, is noteworthy. It is known that the plasmid-mediated AmpC beta-lactamase (AmpC) genes (e.g., act-1) in E. hormaechei can be transferred to other bacterial species, underscoring its potential to disseminate ARGs to bacteria of relevance in human medicine [48]. Regarding the AMR profiles of the isolates from DVMP, beyond their intrinsic resistance (IR) to ampicillin, amoxicillin/clavulanic acid, and cefazolin [33,34], Enterobacter spp. displayed acquired resistance to fluoroquinolones, quinolones, phenicols, and sulphonamides, with 75% (n = 3/4) showing a MDR profile. Similarly, MDR profile was observed in the single E. roggenkampii isolated at DVMM. By contrast, E. hormaechei isolates from DVMM were susceptible to all tested antimicrobials, possibly reflecting differences in environmental pressures or antimicrobial exposure [49]. Notably, 1 E. hormaechei isolated at DVMP isolated from a Common Starling (Sturnus vulgaris) was resistant to colistin, a last-resort antimicrobial for the treatment of MDR Gram-negative human infections [50,51,52]. These findings align with those of Russo et al. [12], who examined 163 wild birds in Southern Italy and detected Enterobacteriaceae in 53.9% of samples, including Enterobacter spp. These isolates frequently exhibited resistance to tetracycline, ciprofloxacin, and trimethoprim/sulfamethoxazole, closely mirroring the resistance profiles of the DVMP isolates in our study. Similarly, Foti et al. [53] reported the isolation of several Enterobacter species from wild birds, many of which were resistant to multiple antimicrobial classes, with high resistance rates to aminoglycosides and, to a lesser extent, fluoroquinolones. In addition, Gargano et al. [54] described E. cloacae carrying beta-lactam resistance determinants in migratory Woodcocks (Scolopax rusticola) from Italy. Although reports of E. hormaechei and E. roggenkampii isolated from wild birds are scarce, limiting the possibility of direct comparisons with existing studies, numerous investigations have described a high occurrence of MDR Enterobacter species in wild bird populations [12,53,55,56]. Together with our findings, this evidence reinforces the notion that wild birds may serve as reservoirs for MDR Enterobacter spp. across diverse geographic contexts.
Pseudomonas aeruginosa, widely distributed in the environment and frequently associated with both animals and humans [57] is recognized as a major nosocomial pathogen due to its intrinsic and acquired resistance mechanisms. For this reason, it has also been designated by the WHO as a priority target for the development of new antimicrobial therapies [36]. Resistance in P. aeruginosa is often linked to biofilm formation and the presence of mobile genetic elements encoding carbapenemases or ESBLs [58,59]. Pseudomonas aeruginosa was isolated from wild birds at both DVMP and DVMM sites. Specifically, this bacterium was recovered from 3 birds at DVMP and from 5 wild birds at DVMM, with a prevalence of 4% (n = 3/75) and 7.6% (n = 5/66), respectively. Aside from the IR of this species to beta-lactams and cephalosporins [33,34], the DVMM isolates were susceptible to most of the tested antimicrobials. Our findings are consistent with those of Russo et al. [12] who reported 18 (n = 18/163; 11%) P. aeruginosa isolates from various wild bird species, all of which were susceptible to the majority of the tested antimicrobials.
Conversely, other studies have reported markedly different resistance profiles among P. aeruginosa isolates from birds [60,61,62]. That said, it should be noted that for this species, the interpretation of susceptibility to several antimicrobials was limited by the absence of standardized clinical breakpoints in current reference guidelines. Nevertheless, certain considerations can be made regarding the MIC values obtained. In our isolates, MICs for flumequine, sulfisoxazole, and aminosidine could be considered high (>16 µg/mL, >512 µg/mL, and >32 µg/mL, respectively), with the exception of 1 DVMM isolate, recovered from a barn swallow (Hirundo rustica), which exhibited a MIC of 8 µg/mL for aminosidine.
Klebsiella pneumoniae is a Gram-negative bacillus included among the ESKAPE group due to its high propensity for acquiring and disseminating ARGs [36,63]. It is a well-recognized cause of serious healthcare-associated infections, including pneumonia, urinary tract infections, septicemia, and surgical site infections [64]. The increasing global prevalence of MDR and carbapenemase-producing K. pneumoniae isolates poses a significant public health threat, severely limiting therapeutic options and contributing to elevated mortality rates [64]. In the present study, a total of 9 K. pneumoniae isolates were recovered: 8 from wild birds sampled at DVMM and 1 from DVMP, corresponding to prevalence of 12.1% (n = 8/66) and 1.3% (n = 1/75), respectively. Excluding the IR to ampicillin reported by EUCAST [34] and CLSI [33] for this species, the DVMP isolate was intermediate to cefazolin but remained susceptible to all other tested antimicrobials. In contrast, the DVMM isolates demonstrated a concerning resistance pattern, with 50% (n = 4/8) classified as MDR. Resistance was most frequently observed to flumequine (n = 4/8; 50%), enrofloxacin (n = 4/8; 50%), tetracycline (n = 3/8; 37.5%), cefazolin (n = 3/8; 37.5%), cefotaxime (n = 3/8; 37.5%), and sulfisoxazole (n = 3/8; 37.5%). Additionally, 2 isolates (n = 2/8; 25%) were resistant to amoxicillin/clavulanic acid and to trimethoprim/sulfamethoxazole. These findings partially align with previous studies reporting a general trend of resistance to tetracyclines, sulfonamides, aminoglycosides, phenicols, and fluoroquinolones [65,66,67], and are also consistent with recent evidence indicating an increasing detection of K. pneumoniae resistant isolates in wildlife species, including birds [68]. Remarkably, all antimicrobial agents to which the K. pneumoniae isolates in this study exhibited resistance are classified as therapeutically important for human medicine [69].
Acinetobacter baumannii is a major opportunistic pathogen in healthcare settings and is also listed by the WHO as a “critical priority” for the development of new antimicrobials due to its frequent MDR phenotype [36,70]. In the present study, a single isolate was isolated from a Eurasian Magpie (Pica pica) at DVMM. These findings are consistent with previous studies on wild bird populations in different countries, where A. baumannii isolation rates have generally been low. For instance, Russo et al. [12] found no A. baumannii among 163 cloacal swabs from wild birds in Southern Italy, and similarly low detection rates have been reported in other geographical contexts [71]. On the opposite, the study by Dahiru et al. [72] in Nigeria reported A. baumannii in 31% (n = 15/48) of fecal samples from wild birds in agricultural settings. In view of its AMR profile, the single A. baumannii isolate, apart from IR displayed toward the most antimicrobial tested, was susceptible to colistin and trimethoprim/sulfamethoxazole, but resistant to gentamicin. Although only a single A. baumannii isolate was recovered in the present study, when considered alongside the existing literature this finding contributes to the evidence that wild avifauna may play a role, albeit limited, in the dissemination of A. baumannii, eventually including isolates of potential public health relevance [25,71,72].
Considering Gram-positive ESKAPE bacteria, S. aureus is a human pathogen associated with both community and hospital-acquired infections, ranging from mild skin inflammation to severe diseases such as pneumonia, sepsis, and endocarditis [73]. Its clinical relevance is further amplified by methicillin-resistant S. aureus (MRSA), which complicates treatment due to resistance to beta-lactams and other antimicrobial classes [73]. In our study, S. aureus isolates were detected only in DVMM, from a Common Blackbird (Turdus merula) and a Tawny Owl (Strix aluco), in association with E. marmotae and E. coli, respectively. Both isolates were resistant to sulfisoxazole and clindamycin but remained susceptible to most other antimicrobials, including fluoroquinolones, cephalosporins, and aminoglycosides. Sánchez-Ortiz et al. [74] reported that 82.6% (n = 19/23) of Staphylococcus spp. isolates from wounds in rehabilitated wild birds were resistant to at least one antimicrobial, with clindamycin resistance being common, while Carrel et al. [75] documented a progressive increase in resistance to sulphonamides (trimethoprim/sulfamethoxazole) in S aureus human isolates in USA. Comparatively, Russo et al. [12] isolated 6 S. aureus (n = 6/163; 3.7%) from wild birds, reporting more frequent resistance to CIAs than that observed in our study. However, the isolate from the Common Blackbird (Turdus merula) in DVMP also exhibited resistance to tetracycline, displaying an MDR phenotype. This finding is noteworthy in light of other studies reporting higher AMR rates in Gram-positive isolates from birds in rural and urban habitats [76,77,78,79]. Although the number of S. aureus isolates in the present study was limited, it is important to note that both isolates were phenotypically susceptible to oxacillin. While the CLSI now recommends cefoxitin as it provides more accurate detection of methicillin resistance phenotypes [33], oxacillin resistance testing has historically been considered a central step in the phenotypic characterization of staphylococci, as it was long regarded as the marker antimicrobial for detecting methicillin-resistant staphylococci (MRS) [80,81]. Based on these results, the S. aureus isolates from DVMP are unlikely to be classified as MRSA, although definitive conclusions cannot be drawn in the absence of further molecular investigations.
Enterococcus faecium is a gut commensal in humans and animals but is also can be a nosocomial pathogen [82]. Its clinical importance stems from its IR to several antimicrobial classes, including cephalosporins and aminoglycosides, and its increasing resistance to ampicillin and vancomycin [33,34,82,83]. For these reasons, E. faecium is among the WHO prioritized ESKAPE [36]. In our study, 10 E. faecium isolates were recovered, 9 (n = 9/66; 13.6%) from DVMM and 1 (n = 1/75; 1.3%) from DVMP. Across both sites, most isolates were susceptible to florfenicol and penicillin, whereas higher resistance rates were observed for tetracyclines, with the single isolate from DVMP and 3 of 9 isolates (33.3%) from DVMM being resistant. Resistance to enrofloxacin was also notable, detected in 44.4% (n = 4/9) of DVMM isolates, which were recovered from 3 specimens of Common Woodpigeon (Columba palumbus) and 1 from a Eurasian Hoopoe (Upupa epops), while the single DVMP isolate was intermediate to this molecule. Additionally, most of DVMM isolates exhibited reduced susceptibility to macrolides and rifampicin. This condition may suggest the presence of ARGs, such as erm(B), although molecular characterization was beyond the scope of this study and warrants further investigation [82,84,85]. The findings of the present study are consistent with previous reports confirming the occurrence of resistance to fluoroquinolones, macrolides, and tetracyclines in E. faecium from wildlife [12,27,76,86]. Although resistance to these agents is so widespread that they are rarely used for the treatment of enterococcal infections [82], the results observed in this and similar studies are concerning, as they reveal substantial resistance to antimicrobials of public health relevance [58]. Notably, 1 E. faecium isolate from a Common Woodpigeon (Columba palumbus) at DVMM exhibited a MDR phenotype, showing resistance to penicillin, ampicillin, amoxicillin/clavulanic acid, enrofloxacin, erythromycin, and tetracycline. These results may underscore the influence of urban environments on the AMR, as the antimicrobials tested belong to the classes mostly used in humans as well as in companion animals, exerting considerable selective pressure at the environmental level [87,88]. Lastly, in the present study most E. faecium isolates exhibited uniformly elevated MIC values for kanamycin, with 2 isolates exceeding 250 µg/mL. This finding aligns with the well-documented IR of E. faecium to low-level concentration of several aminoglycosides, including kanamycin, mediated by a chromosomally encoded aminoglycoside-modifying enzyme [82,89]. Wild-type E. faecium typically shows kanamycin MICs in the range of 128–256 µg/mL; however, the markedly high MIC values (≥500 µg/mL) observed in the 2 DVM isolates may indicate the presence of additional acquired aminoglycoside resistance determinants [89], underscoring the need for further molecular characterization.
A special attention should be given to the high number of non-ESKAPE bacterial isolates (n = 206/243; 84.8%) recovered from both DVMM and DVMP, as several species are considered of relevance for public health.
Escherichia coli was the most frequently isolated bacterium, both at DVMM (n = 38/66; 57.6%) and DVMP (n = 46/75; 61.3%). Recent literature increasingly includes E. coli within the ESKAPE group, hence the term “ESKAPEE”, due to its frequent MDR phenotype and widespread environmental distribution [90,91]. In our samples, E. coli was often recovered in association with other bacteria, particularly Gram-positive bacteria such as E. faecalis and S. aureus, especially in passerines and Columbiformes. Such interspecies co-occurrence may enhance HGT, particularly in complex environmental microbiomes, thereby accelerating the spread of AMR among bacterial populations [92]. Additionally, 6 (n = 6/75; 8%) E. marmotae isolates were collected at DVMP, isolated with other relevant bacterial species, such as S. aureus and E. faecalis. This species is recognized as an emerging pathogen capable of acquiring ESBL and AmpC resistance genes [93]. Other non-ESKAPE but clinically significant bacteria detected included Salmonella enterica serovar Typhimurium and Bacillus cereus, both known not only as zoonotic agents, but also as bacteria with established resistance mechanisms [94]. These bacteria were recovered exclusively from DVMP with a prevalence of 2.7% (n = 2/75) and 1.3% (n = 1/75), respectively. Citrobacter spp., in particular C. freundii, C. koseri, and C. braakii, were isolated in both study sites. These opportunistic pathogens are known to harbor and exchange ARGs [32,95,96,97]. Klebsiella oxytoca and K. aerogenes were also detected, at DVMP from a Little Owl (Athene noctua) and a Barn Owl (Tyto alba), and at DVMM from a Eurasian Magpie (Pica pica) and a Common Blackbird (Turdus merula). These sedentary species share habitats with humans, potentially facilitating cross-species transmission of ARB. Although these bacteria are generally opportunistic pathogens in immunocompromised individuals, they can act as important vectors for the transfer of ARGs, including ESBLs and carbapenemases [98,99]. Finally, Mammaliicoccus sciuri (formerly Staphylococcus sciuri) and S. xylosus, common skin commensals with potential zoonotic relevance, were predominantly isolated in DVMP, particularly from raptors. Mammalicoccus sciuri is a recognized reservoir of S. aureus mecA homologues within Staphylococcal Cassette Chromosome (SCCmec) elements, owing to their high sequence homology and phylogenetic relatedness to mecA [100].
This study presents several strengths, including the simultaneous investigation of ESKAPE pathogens in wild birds from two geographically and ecologically distinct areas of Italy, Umbria and Lombardy. These regions were deliberately selected as they could represent markedly different ecological contexts, both in terms of natural environments and degrees of anthropogenic influence, thus providing a comprehensive overview of the occurrence of ESKAPE bacteria and their AMR profiles in wild birds. This dual-site approach can contribute a broader assessment of the potential role of avifauna in the dissemination of ARB across distinct Italian landscapes. Overall, the detection of AMR ESKAPE bacteria in both regional sites, particularly in Central Italy, where anthropogenic impact is lower compared to Lombardy, highlights the importance of wild birds as reservoirs and potential vectors of AMR bacteria and determinants, reinforcing the need for ongoing surveillance within a One Health framework. Furthermore, the inclusion of multiple avian species with diverse ecological niches increases the representativeness and ecological relevance of the sampled population. Nonetheless, some limitations should be acknowledged. First of all, the unequal number of samples collected between the two study sites, together with the biodiversity of the birds investigated may limit the robustness of direct statistical comparisons. Moreover, it should be emphasized that the primary aim of this work was descriptive in nature, focusing on the detection of ESKAPE bacteria in wild birds and the characterization of their AMR profiles. Accordingly, the study was not deliberately designed for inferential statistical analyses, which would have required larger and more homogeneous sample sizes. Rather, the value of the present investigation lies in providing baseline data that can contribute to addressing existing knowledge gaps and serve as a reference for future, more extensive studies. Another important limitation refers to the fact that, for several antimicrobial agents, the absence of established clinical breakpoints limited the interpretation of MIC values resulting in either under- or overestimation of resistance levels. The absence of molecular analyses, as this was not the objective of our study, restricted the ability to identify the genetic basis of the observed resistance phenotypes. To address this gap, targeted molecular analyses are planned as a follow-up to this work, thereby clarifying the genetic basis of the observed resistance phenotypes.
4. Materials and Methods
4.1. Sampling
A total of 141 cloacal swabs were collected from various species of wild birds between February and September 2024. In particular 75 samples were obtained from birds coming from the Wild Umbria Rescue Centre (Umbria region) or occasionally brought by private citizens and received at the DVMP hospital. The remaining 66 samples were obtained from birds housed at the WWF Wildlife Rescue Centre in Vanzago (Lombardy region), affiliated with the DVMM.
At the time of admission to the rehabilitation centers, as well as to the hospital at the DVMP site, an identification form was completed for each animal, recording the species, reason for admission, medical history, the date and location of finding.
Sampled birds were admitted due to various traumatic injuries, including those caused by collisions with vehicles, hunting or attacks by domestic animals. All cloacal swabs were collected, in both the rescue centers and/or, in the case, of DVMP, at the veterinary hospital, by the attending veterinarian during the clinical examination under standard, non-invasive conditions, before any antimicrobial treatment was given. After collection, cloacal swabs were placed in sterile tubes containing semisolid Amies transport medium (Copan Italia S.p.A., Brescia, Italy) and immediately stored at 4 °C and transported to the respective microbiology laboratories, either the Laboratory of Infectious Diseases in Perugia or the Avian Pathology Unit in Lodi (Milan), for further analyses.
4.2. Bacterial Isolation and Identification
To isolate and identify bacterial species from the sampled wild birds, an initial enrichment step was performed by inoculating each cloacal swab into sterile tubes containing 10 mL of Buffered Peptone Water (BPW) (ThermoFisher, Oxoid, UK), followed by incubation at 37 °C for 18–24 h. After enrichment, aliquots of the broth cultures were plated onto different growth media simultaneously, represented by Blood agar (ThermoFisher, Oxoid, UK), MacConkey agar (ThermoFisher, Oxoid, UK), and Brilliance UTI Clarity agar (ThermoFisher, Oxoid, UK), and incubated under aerobic conditions at 37 °C for an additional 18–24 h. Bacterial growth was subsequently examined, and representative colonies were selected based on morphological and phenotypic characteristics for species-level identification. This was achieved through Matrix-Assisted Laser Desorption/Ionization Time-of-Flight (MALDI-TOF) mass spectrometry (MS) using the MBT Microflex LT/SH system (Bruker Daltonik GmbH, Bremen, Germany), following the manufacturer’s instructions and protocol adapted from Rosa et al. [101], with the direct colony transfer method. Briefly, a small portion of each isolated colony was applied onto duplicate wells of disposable target plates using a sterile toothpick. The spots were then overlaid with 1 µL of α-cyano-4-hydroxycinnamic acid (HCCA) matrix solution (50% acetonitrile, 47.5% water, 2.5% trifluoroacetic acid; Bruker Daltonik GmbH, Bremen, Germany) and allowed to dry at room temperature. Mass spectra were then acquired in positive ion mode. Each run included duplicate spots of the Bruker Bacterial Test Standard. Spectra were analyzed against the MALDI Biotyper (MBT) Compass^®^ Library (HT) software (2023, Bruker Daltonik GmbH, Bremen, Germany), that encompasses 4320 species across 712 microorganism genera; bacterial identification was assigned based on the degree of overlap between the mass spectral profiles of the analyzed isolates and the reference spectra contained in the library. No subsequent serotyping analyses, nor phylogenetic or molecular confirmation methods, were applied in this study.
For the isolation of Salmonella spp., samples were processed according to the International Standard ISO 6579-1:2017 [102]. Colonies that growing on selective and differential media were confirmed and identified at the genus level using a MALDI-TOF mass spectrometer (MBT Microflex LT/SH, Bruker Daltonik GmbH, Bremen, Germany) applying the protocol described before. Serotyping was performed according to the Kaufmann–White–Le Minor scheme [103] using direct slide agglutination with the specific antisera (Statens Serum Institute, Copenhagen, Denmark).
4.3. Antimicrobial Susceptibility Testing (AST)
The antimicrobial susceptibility of ESKAPE isolates was assessed by broth microdilution method to determine the MIC value of each antimicrobial compound. All tests were performed at the Experimental Zooprophylactic Institute of Lombardy and Emilia-Romagna, “Bruno Ubertini”, Territorial Laboratory in Lodi. This laboratory provided full diagnostic support for both the DVMM and DVMP cohorts. Analyses were performed using the Sensititre System for MIC plate testing (Thermo Fisher Scientific, Oxoid, UK). To ensure the quality and accuracy of AST procedures, American Type Culture Collection (ATCC) standard reference (E. coli ATCC 25922, S. aureus ATCC 29213) were used. The testing procedures followed standardized protocols [104,105].
For Gram-negative ESKAPE isolates, the following antimicrobials were tested across defined concentration ranges: aminosidine (AN; 1–32 µg/mL), amoxicillin/clavulanic acid (AMC; 0.25–32 µg/mL), ampicillin (AMP; 0.25–32 µg/mL), cefazolin (CFZ; 0.5–8 µg/mL), cefotaxime (CTX; 0.5–4 µg/mL), colistin (CL; 0.03125–8 µg/mL), enrofloxacin (ENR; 0.015625–32 µg/mL), florfenicol (FFC; 1–64 µg/mL), flumequine (FLU; 1–16 µg/mL), gentamicin (GEN; 0.25–32 µg/mL), kanamycin (KAN; 2–32 µg/mL), sulfisoxazole (SFX; 128–512 µg/mL), tetracycline (TET; 0.5–16 µg/mL), and trimethoprim/sulfamethoxazole (SXT; 0.0625–16 µg/mL). For Gram-positive isolates, the antimicrobials tested included: amoxicillin/clavulanic acid (AMC; 0.25–16 µg/mL), ampicillin (AMP; 0.03125–16 µg/mL), cefazolin (CFZ; 0.25–8 µg/mL), ceftiofur (CTF; 0.25–8 µg/mL), clindamycin (DA; 0.5–2 µg/mL), enrofloxacin (ENR; 0.25–4 µg/mL), erythromycin (ERY; 0.03125–8 µg/mL), florfenicol (FFC; 2–8 µg/mL), kanamycin (KAN; 8–32 µg/mL and 250–500 µg/mL), oxacillin (OX; 0.25–4 µg/mL), penicillin (P; 0.03125–16 µg/mL), rifampicin (RD; 0.625–2 µg/mL), sulfisoxazole (SFX; 128–512 µg/mL), tetracycline (TET; 0.25–16 µg/mL), tilmicosin (TIL; 8–32 µg/mL), and trimethoprim/sulfamethoxazole (SXT; 0.125–8 µg/mL).
Bacterial susceptibility to each antimicrobial agent was first classified as susceptible (S), intermediate (I), and resistant (R) based on the breakpoints provided by the European Committee on Antimicrobial Susceptibility Testing (EUCAST), the Clinical and Laboratory Standards Institute (CLSI), or the Comité de l’Antibiogramme de la Société Française de Microbiologie (CA-SFM). Isolates showing intermediate susceptibility were then considered susceptible for the evaluation of the results [106,107]. As a general rule, EUCAST breakpoints [106,108] were preferentially applied. When not available, CLSI [33,109] and CA-SFM recommendations [110,111] breakpoints were used as an alternative reference. The latter were specifically used for the interpretation of MIC values for aminosidine (also known as paromomycin), as reported in other studies [112], and for kanamycin, particularly in Enterobacteriaceae and S. aureus, respectively. In general, preference was given to the most recent editions of the aforementioned guidelines; when breakpoints were not available in those versions, older editions were consulted. In the absence of established clinical breakpoints, epidemiological cut-off values (ECOFFs) defined for the target bacterial species, or, when unavailable, for closely related species, were considered [113]. Where no standardized breakpoints were available in any of the aforementioned sources, raw MIC values were reported. Specifically, given the absence of specific clinical breakpoints, MIC values for enrofloxacin were interpreted using those established for ciprofloxacin as a representative fluoroquinolone, consistently with the approach adopted in previous studies [114,115]. The sources used for the interpretation of AST results are reported in the Supplementary Materials, Table S3.
MDR bacteria were classified as those resistant to at least one antimicrobial agent in three or more distinct classes of antimicrobials [116].
5. Conclusions
This study investigated preliminarily the role of wild birds as potential reservoirs and sentinels of ARB, including clinically relevant ESKAPE bacteria. Among these, Enterobacter spp. were the most frequently isolated in DVMP, while E. faecium and K. pneumoniae were more commonly recovered in DVMM. Although E. coli is not universally included in the ESKAPE group, its frequent detection at both study sites, often in association with other Gram-positive and Gram-negative bacteria, highlights its ecological importance and potential role in HGT. Such interactions may facilitate the dissemination of AMR traits across bacterial species, enhancing microbial adaptability and persistence in different habitats.
Despite certain limitations related to our investigation, our results support the hypothesis that wild birds may harbor bacteria with both intrinsic and acquired resistance determinants. These include ESKAPE bacteria as well as other zoonotic or emerging pathogens of public health concern. The findings of this study reinforce the importance of integrating wildlife into AMR surveillance programs, particularly within a One Health framework, to better understand and mitigate the environmental dissemination of resistance. Continued monitoring in ecologically distinct areas, such as those represented by DVMM and DVMP, will be essential for identifying emerging threats and informing targeted interventions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1WHO Ten Threats to Global Health in 2019 Available online: https://www.who.int/news-room/spotlight/ten-threats-to-global-health-in-2019(accessed on 14 April 2025)
- 2O’Neill J. Antimicrobial resistance: Tackling a crisis for the health and wealth of nations The Review on Antimicrobial Resistance Wellcome Trust London, UK 2014116
- 3Salam M.A. Al-Amin M.Y. Salam M.T. Pawar J.S. Akhter N. Rabaan A.A. Alqumber M.A.A. Antimicrobial resistance: A growing serious threat for global public health Healthcare 202311194610.3390/healthcare 1113194637444780 PMC 10340576 · doi ↗ · pubmed ↗
- 4Holmes A.H. Moore L.S.P. Sundsfjord A. Steinbakk M. Regmi S. Karkey A. Guerin P.J. Piddock L.J.V. Understanding the mechanisms and drivers of antimicrobial resistance Lancet 201638717618710.1016/S 0140-6736(15)00473-026603922 · doi ↗ · pubmed ↗
- 5Laxminarayan R. Duse A. Wattal C. Zaidi A.K.M. Wertheim H.F.L. Sumpradit N. Vlieghe E. Hara G.L. Gould I.M. Goossens H. Antibiotic resistance—The need for global solutions Lancet Infect. Dis.2013131057109810.1016/S 1473-3099(13)70318-924252483 · doi ↗ · pubmed ↗
- 6Caneschi A. Bardhi A. Barbarossa A. Zaghini A. The use of antibiotics and antimicrobial resistance in veterinary medicine, a complex phenomenon: A narrative review Antibiotics 20231248710.3390/antibiotics 1203048736978354 PMC 10044628 · doi ↗ · pubmed ↗
- 7De Oliveira D.M.P. Forde B.M. Kidd T.J. Harris P.N.A. Schembri M.A. Beatson S.A. Paterson D.L. Walker M.J. Antimicrobial resistance in ESKAPE pathogens Clin. Microbiol. Rev.202033 e 00181-1910.1128/CMR.00181-1932404435 PMC 7227449 · doi ↗ · pubmed ↗
- 8Navidinia M. The clinical importance of emerging ESKAPE pathogens in nosocomial infections Arch. Adv. Biosci.20167435710.22037/jps.v 7i 3.12584 · doi ↗