In Vitro Activity of Delafloxacin Against Corynebacterium spp
Montserrat Muñoz-Rosa, Cristina Elías-López, Rosa Pedraza, Cristina Riazzo, Cristina Arjona-Torres, Isabel Machuca, Rocio Tejero-García, Julian Torre-Cisneros, Luis Martínez-Martínez

TL;DR
This study evaluates how well delafloxacin works against different Corynebacterium species in the lab and finds it is more effective than other fluoroquinolones.
Contribution
The study introduces delafloxacin as a potential alternative treatment for Corynebacterium infections and evaluates its efficacy compared to other drugs.
Findings
Delafloxacin showed the lowest MIC50/MIC90 values of 0.5/8 mg/L against Corynebacterium isolates.
Mutations in the QRDR of the gyrA gene were linked to increased resistance to delafloxacin.
Delafloxacin gradient strips demonstrated high agreement with microdilution methods.
Abstract
Background/Objectives: The susceptibility of Corynebacterium spp. to antimicrobial agents is species-related, with increasing levels of resistance to fluoroquinolones in several species related to their continued use in clinical practice. The objectives were to determine the in vitro activity of delafloxacin in comparison with other fluoroquinolones against clinical isolates of Corynebacterium spp., to compare MICs of delafloxacin obtained with gradient strips and with reference microdilution, and to investigate the mechanisms related to fluoroquinolone resistance in the tested strains. Methods: Fifty-three clinical isolates, assigned to five species of Corynebacterium spp., were evaluated using reference microdilution for delafloxacin, ciprofloxacin, levofloxacin, and moxifloxacin (with and without reserpine or phenylalanine-arginine β-naphthylamide), and gradient strips for…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
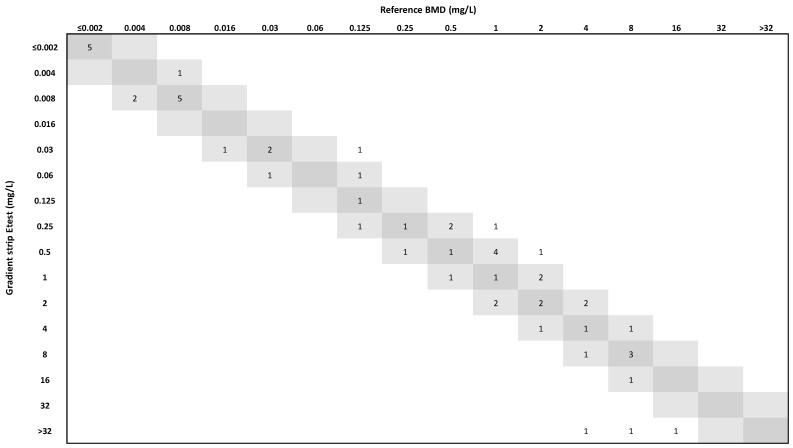 Figure 1
Figure 1- —Laboratorios Menarini S.A.
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDiphtheria, Corynebacterium, and Tetanus · Clostridium difficile and Clostridium perfringens research · Bacterial Identification and Susceptibility Testing
1. Introduction
Corynebacterium species other than C. diphteriae are members of the normal microbiota of skin and mucosa, but have also been described as etiologic agents of a variety of human infections [1,2,3]. Susceptibility of Corynebacterium spp. to antimicrobial agents is species-related, and several species are frequently resistant to multiple antimicrobial agents [4,5,6]. Glycopeptides and linezolid present excellent in vitro activity against most clinically relevant species [1,4,5,6,7]. β-lactams, fluoroquinolones, aminoglycosides, macrolides, and tetracyclines are also of therapeutic interest for Corynebacterium infections [4,5,6].
Previous studies have evaluated the in vitro activity of fluoroquinolones, including ciprofloxacin [6,8,9,10], levofloxacin [6,11], and moxifloxacin [6,10], against Corynebacterium spp. As for other organisms, higher rates of resistance to fluoroquinolones in Corynebacterium spp. have been observed after the continuous use of fluoroquinolones in clinical practice [8,9,10,11,12]. Corynebacterium spp. lack topoisomerase IV [9], and their resistance to fluoroquinolones is primarily linked to point mutations in the quinolone resistance determining region (QRDR) of the gyrA gene, encoding the subunit A of DNA gyrase [9,12,13]. The improved activity of delafloxacin against multiple Gram-positive bacteria compared to older quinolones suggests it may also be more active against Corynebacterium spp.
Delafloxacin is a new non-zwitterionic dual quinolone, with affinity for both DNA gyrase and topoisomerase IV [14,15]. It presents better in vitro activity against Staphylococcus aureus and Streptococcus pneumoniae than older quinolones [16]. There is limited information on the activity of delafloxacin against Corynebacterium spp. In one study on bacteria cultured from patients with cancer [17], minimal inhibitory concentration (MIC) range, MIC_50_, and MIC_90_ of delafloxacin against 30 Corynebacterium spp. were ≤0.03–4, 0.06, and 2 mg/L, much lower than those of ciprofloxacin and levofloxacin. In another recent study [18] on clinical isolates from a single centre (n = 116, 69% C. striatum and 16% C. amycolatum), MIC range, MIC_50_, and MIC_90_ of delafloxacin were 0.002–>1, 1, and >1 mg/L.
Delafloxacin is not yet included in the antimicrobial panels for automatic susceptibility testing systems commonly used in clinical laboratories, and in vitro testing of this compound is usually performed using discs or gradient strips, although evidence of the reliability of these approaches when considering Corynebacterium spp. is lacking.
The objectives of this study were as follows: (i) to determine the in vitro activity of delafloxacin in comparison with ciprofloxacin, levofloxacin, and moxifloxacin against a collection of clinical isolates of Corynebacterium spp. using standardized broth microdilution (BMD), (ii) to compare MIC values of delafloxacin obtained with gradient strips with those obtained with BMD, and (iii) to investigate the mechanisms related to fluoroquinolone resistance in the tested strains.
2. Results
The MIC range, MIC_50_, and MIC_90_ values obtained with BMD and gradient strips are presented in Table 1. Detailed fluoroquinolone MICs and changes in QRDR of gyrA for every isolate are presented in Table 2.
MICs of moxifloxacin were at least two times lower than those of ciprofloxacin for 45/53 (84.9%) isolates, although for C. glucuronolyticum MICs of moxifloxacin were the same (4/10 isolates) or two times higher than MICs of ciprofloxacin (3/10 isolates). Similarly, MICs of moxifloxacin were at least four times lower than those of levofloxacin for 42/53 (79.2%). MICs of ciprofloxacin and levofloxacin were within one 2-log dilution step for C. jeikeium (10/10 isolates), C. striatum (10/10 isolates), and C. urealyticum (9/10 isolates), but were lower for ciprofloxacin in the case of C. glucuronolyticum (8/10 isolates) and C. amycolatum with wild-type gyrA (4/4).
Delafloxacin was at least two times more active than the three other tested fluoroquinolones for 50/53 (94.3%) isolates. The remaining three isolates correspond to one C. jeikeium with a double mutation in QRDR, and two C. glucuronolyticum with wild-type QRDR. For the 53 Corynebacterium spp. evaluated (when using reference BMD), 43.4% and 26.4% were inhibited at ≤0.25 mg/L and ≤0.016 mg/L. At ≤0.25 mg/L, the highest percentage of inhibition was observed for C. amycolatum (84.6%) and the lowest for C. striatum (0%) and C. urealyticum (10.0%) (Table 1).
MICs of delafloxacin were just 4, 8, 16, and ≥32 times lower than those of moxifloxacin for 21, 13, 3, and 4 isolates, and for 3 and 1 additional isolates they were ≥4 and ≥8 times lower (Table 2). From another point of view and for the complete set of tested isolates, the MIC_50_ of delafloxacin by the reference method was 8, 16, and 32 times lower than the MIC_50_ of moxifloxacin, ciprofloxacin, and levofloxacin, respectively. Similarly, the MIC_90_ of delafloxacin was 4, >4, and >4 times lower than the MIC_90_ of moxifloxacin, ciprofloxacin, and levofloxacin, respectively (Table 1).
All isolates (except one C. amycolatum) for which MICs of delafloxacin were ≤0.016 mg/L had a wild type gyrA QRDR. Most gyrA QRDR mutations occurred in position 87 (Ser in the five evaluated species) alone or combined with a second mutation in position 91 (Asp in the wild type sequences). In several species, a particular mutation pattern was observed in most isolates (i.e., Ser87Ile in C. glucuronolyticum, Ser87Phe plus Asp91Ala in C. striatum, or Ser87Val plus Asp91Tyr in C. urealyticum). As far as we know, some novel mutations in a concrete species have also been observed, as detailed in Table 2, expanding our information on the importance on this region on resistance of Corynebacterium spp. to fluoroquinolones. A point mutation in the QRDR of gyrA was related to a moderate increase in MIC values, while double mutations were observed among isolates with MICs of delafloxacin ranging from 0.125 to 16 mg/L (Table 2). This confirms previous observations in Corynebacterium spp. [9,12,13].
As shown in Table 2, MICs of delafloxacin for isolates of the same species with just the same gyrA QRDR mutation(s) are not the same: i.e., for C. glucuronolyticum isolates with the Ser87Ile change, MICs range from 0.25 to 2 mg/L, and for C. amycolatum with Ser87Arg, MICs range from 0.016 to 0.125 mg/L, suggesting the presence of additional mechanism(s) of resistance. However, for all tested isolates, MIC values in the presence of the efflux pump inhibitors reserpine or phenylalanine-arginine β-naphthylamide (PAβN) were the same or within one two-fold dilution of those obtained in the absence of the corresponding inhibitor.
The delafloxacin gradient strips showed an essential agreement (EA) of 88.7%, slightly below the threshold of the acceptance criteria (≥90%). Bias was −5% compared to the reference method, which is inside the acceptable range of ±30%. However, the categorical agreement (CA) was 92.5% and the Kappa index was 0.848, indicating an almost perfect concordance. MIC_50_ and MIC_90_ values of delafloxacin by BMD and gradients strips were the same or within one dilution step, except for the MIC_50_ for C. jeikeium and the MIC_90_ for C. urealyticum (Table 1, Figure 1).
3. Discussion
Because of the increasing clinical relevance of multiple species of Corynebacterium spp. other than C. diphtheriae and their frequent resistance to available antimicrobial agents, it is necessary to evaluate the activity of new compounds against these organisms. Delafloxacin is a new fluoroquinolone of great therapeutic interest because of its dual activity against both DNA gyrase and topoisomerase IV [6,7]. This translates into an improved activity against different Gram-positive bacteria (i.e., S. aureus, Streptococcus pneumoniae), for which ciprofloxacin and most other fluoroquinolones preferentially target topoisomerase IV [19]. Although Corynebacterium spp. lacks topoisomerase IV, delafloxacin is still more active against these microorganisms than the three tested fluoroquinolones. This can be related to the previous observation that, beyond its dual action on DNA gyrase and topoisomerase IV, the main target of delafloxacin for S. aureus is, in fact, GyrA, for which delafloxacin is about six times more active than ciprofloxacin [20]. It is likely that targeting DNA gyrase is more efficient than interacting with topoisomerase IV, since the former works ahead of the replication fork to remove DNA supercoils and, consequently, inhibits faster DNA replication. Additionally, delafloxacin differs structurally from other quinolones in presenting a 3-hydroxyazetidine-1-yl substituent at position 7 of the 6-fluoroquinolone core [20], which determines that under acidic conditions the compound is predominantly in a non-ionized form, which improves the activity in acid environments and, in addition, may favor the intrabacterial accumulation of the compound in comparison with other fluoroquinolones.
A large proportion (40/53; 75.5%) of the isolates evaluated in this study were resistant to both ciprofloxacin and moxifloxacin, according to European Committee on Antimicrobial Susceptibility Testing (EUCAST) breakpoints (breakpoints have not been defined for levofloxacin). Delafloxacin is much more active than the tested comparators, in particular against C. amycolatum, C. glucuronolyticum, and, to a lesser extent, C. jeikeium, although MICs of delafloxacin increase in parallel to those of comparative quinolones, as also indicated in a recent report [18]. The worse results were obtained with C. striatum and C. urealyticum; both species often show high resistance rates to all fluoroquinolones, limiting their use as possible treatment options.
In the absence of clinical breakpoints of delafloxacin for Corynebacterium spp. to interpret susceptibility testing data, it is difficult to evaluate the actual impact of its improved in vitro activity from a therapeutic point of view. Considering EUCAST breakpoints for S. aureus (susceptible: ≤0.25 mg/L for skin and skin structure infections and ≤0.016 mg/L for community-acquired pneumonia, respectively), only 45.3% and 21.4% of the tested isolates were below these reference breakpoints. As a comparison, 75.5% and 71.7% of the isolates were resistant to ciprofloxacin and moxifloxacin, respectively.
Despite the limited number of studies in which mutations in the QRDR of the gyrA gene have been determined in Corynebacterium spp., most mutations detected in this study have already been reported. As an example, the Ser87Phe plus Asp91Ala double change frequently observed in C. striatum in this study was also the most common pattern in other studies [9,12].
Beyond the clear importance of gyrA mutations in compromising the in vitro activity of delafloxacin, the observation that isolates with the same mutations present different MICs of the compound suggests that other mechanisms of resistance modulate delafloxacin resistance in Corynebacterium spp. Resistance to fluoroquinolones caused by efflux pumps has been described in Staphylococcus spp. and other Gram-positive organisms [21,22], which can be easily demonstrated using active efflux inhibitors [23,24]. The relevance of active efflux in the resistance of Corynebacterium spp. to tetracycline and other compounds has also been previously reported [25,26,27]. We hypothesized that efflux pumps could also contribute to fluoroquinolone resistance in our isolates, but we have not observed any relevant difference in the MICs of fluoroquinolones determined in the presence of reserpine or PaβN. This may indicate that active efflux is not really an important mechanism of resistance of the tested isolates against delafloxacin and other fluoroquinolones, or that the inhibitors are not adequate to inhibit such a mechanism, perhaps because their penetration into corynebacteria is limited due to the different surface structure of this microorganism compared to Staphylococcus or other Gram-positive bacteria. New studies are warranted to evaluate the importance of active efflux and other mechanisms as a cause of delafloxacin resistance in Corynebacterium spp.
Although the reference method for determining the MIC of delafloxacin is broth microdilution, this method is difficult to implement in daily work, and, as this antibiotic is not yet included in most commercial panels of semi-automated microdilution systems, we have explored the reliability of using gradient strips. The obtained results suggest that this approach may represent an acceptable option for testing delafloxacin against Corynebacterium spp., although further studies are needed to define categorical errors with the strips when, eventually, specific clinical breakpoints are defined.
The major limitation of this study is that we have focused just on the five species commonly found in our centre, but have not evaluated other species of Corynebacterium or related organisms, and that the number of studied isolates was also limited. Further research on these groups of Gram-positive bacteria is also warranted.
4. Materials and Methods
4.1. Bacterial Strains
Fifty-three clinical isolates (one per patient) of Corynebacterium spp., including C. amycolatum (n = 13), C. glucuronolyticum (n = 10), C. jeikeium (n = 10), C. striatum (n = 10), and C. urealyticum (n = 10), recovered at the clinical microbiology laboratory of the Reina Sofía University Hospital of Cordoba, Spain, and representing the most frequently identified species at our centre during 2018–2022, were evaluated. Species were identified using MALDI-TOF/MS (MALDI Biotyper; Bruker Daltonics, Bremen, Germany).
4.2. Susceptibility Testing
MICs of delafloxacin, ciprofloxacin, levofloxacin, and moxifloxacin (all from MedChemExpress, Monmouth Junction, NJ, USA), with and without reserpine (20 mg/L) (MedChemExpress) and PAβN (25 mg/L) (Sigma-Aldrich, Taufkirchen, Germany), were determined by reference BMD (ISO guidelines) [28] using cation-adjusted Mueller–Hinton broth (Sigma-Aldrich) supplemented with 5% lysed horse blood (ThermoFisher, Waltham, MA, USA) and 20 mg/L β-NAD (Sigma-Aldrich) according to the EUCAST recommendations for fastidious organisms. Streptococcus pneumoniae ATCC 49619 and Staphylococcus aureus ATCC 29213 were used as control strains.
Additionally, susceptibility to delafloxacin was also determined with gradient strips (Etest, BioMèrieux, Rhone, France) using Mueller–Hinton agar with 5% lysed horse blood and 20 mg/L β-NAD (BioMérieux, France). Although EUCAST has not defined clinical breakpoints of delafloxacin for Corynebacterium, for analyzing the impact of gyrA mutations on the MICs of delafloxacin, the EUCAST breakpoints (V_15.0) [29] for Staphylococcus aureus (susceptible: ≤0.25 and ≤0.016 mg/L, for skin and skin structure infections and for community-acquired pneumonia, respectively) were considered.
To evaluate the performance of the gradient strips, EA and bias were calculated according to the recommendations in ISO-20776-2:2021 [30]. Rates of CA were also calculated following the definitions by ISO 20776-2:2007 [31]. The Kappa index was used to evaluate concordance based on Landis and Koch guidelines [32], and was calculated using the Statistical Package for the Social Sciences (IBM SPSS Statistics 19.0, Armonk, NY, USA) software.
4.3. Sequencing and Gene Analysis
To amplify the QRDR of the gyrA gene of C. amycolatum, C. jeikeium, C. striatum, and C. urealyticum, primers CorynA1: GCGGCTACGTAAAGTCC and CorynA2: CCGCCGGAGCCGTTCAT [9] were used. A novel pair of primers for QRDR of C. glucuronolyticum were designed (GluQRDR-F: TACGCATCCTTAATGCCCTG and GluQRDR-R: CCAATCGACCTGAATGAGGA). The amplicons (both strands) were sequenced using Sanger technology (Sistemas Genómicos, Valencia, Spain). Quality control of the sequences was assessed with CLC Genomics Workbench 24.0.3 (Qiagen, Germantown, MD, USA) by manual check and correction of basecalls in the electropherogram and end-trimming. Mutations were defined after BLAST alignment [https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 1 January 2025)] against the following corresponding reference sequences: C. amycolatum quinolone-susceptible clinical isolate [9] (Accession number NCBI: AY559039), C. glucuronolyticum strain FDAARGOS_1053 (Accession number NCBI: NZ_CP066007.1), C. jeikeium ATCC 43734 (Accession number NCBI: NZ_GG700813.1), C. striatum ATCC 6940 (Accession number NCBI: AY559038), and C. urealyticum DSM 7109 [33] (Accession number NCBI: NC_010545.1).
5. Conclusions
In summary, compared to the other fluoroquinolones analyzed, delafloxacin was the most in vitro active compound against Corynebacterium spp. It may represent a therapeutic alternative for infections caused by these organisms, and could help preserve the efficacy of fluoroquinolones through targeted therapy. The clinical implications of these findings warrant further study, including clinical efficacy and risk of emergence of new resistance.
The increase in the MICs of fluoroquinolones against Corynebacterium spp. is related to non-conservative mutations in the QRDR of the gyrA gene of the studied isolates. The level of increase depends on whether there are one or two mutations, their positions, and the considered species. Indirect evidence suggests that additional mechanisms contribute to decreased activity of delafloxacin against Corynebacterium spp.
Gradient strips may represent a reasonable strategy for determining the MIC of delafloxacin against Corynebacterium spp. in routine work.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Funke G. von Graevenitz A. Clarridge J.E.3rd Bernard K.A. Clinical microbiology of coryneform bacteria Clin. Microbiol. Rev.19971012515910.1128/CMR.10.1.1258993861 PMC 172946 · doi ↗ · pubmed ↗
- 2Bernard K. The genus Corynebacterium and other medically relevant coryneform-like bacteria J. Clin. Microbiol.2012503152315810.1128/JCM.00796-1222837327 PMC 3457441 · doi ↗ · pubmed ↗
- 3Zasada A.A. Mosiej E. Contemporary microbiology and identification of Corynebacteria spp. causing infections in human Lett. Appl. Microbiol.20186647248310.1111/lam.1288329573441 · doi ↗ · pubmed ↗
- 4Olender A. Antibiotic resistance and detection of the most common mechanism of resistance (MLSB) of opportunistic Corynebacterium Chemotherapy 20135929430610.1159/00035746724480817 · doi ↗ · pubmed ↗
- 5Khodadadi R.B. El Zein S. Rivera O’Connor C.G. Stevens R.W. Schuetz A.N. Abu Saleh O.M. Fida M. Retrospective analysis of antimicrobial susceptibility profiles of non-diphtheriae Corynebacterium species from a tertiary hospital and reference laboratory, 2012–2023 J. Clin. Microbiol.202462 e 011992410.1128/jcm.01199-2439555924 PMC 11633113 · doi ↗ · pubmed ↗
- 6Neemuchwala A. Soares D. Ravirajan V. Marchand-Austin A. Kus J.V. Patel S.N. In Vitro Antibiotic Susceptibility Pattern of Non-diphtheriae Corynebacterium Isolates in Ontario, Canada, from 2011 to 2016 Antimicrob. Agents Chemother.201862 e 01776-1710.1128/AAC.01776-1729339389 PMC 5914010 · doi ↗ · pubmed ↗
- 7Gómez-Garcés J.L. Alos J.I. Tamayo J. In vitro activity of linezolid and 12 other antimicrobials against coryneform bacteria Int. J. Antimicrob. Agents 20072968869210.1016/j.ijantimicag.2006.11.03217475450 · doi ↗ · pubmed ↗
- 8Martínez-Martínez L. Suárez A.I. Ortega M.C. Perea E.J. Comparative in vitro activities of new quinolones against coryneform bacteria Antimicrob. Agents Chemother.1994381439144110.1128/AAC.38.6.14398092851 PMC 188228 · doi ↗ · pubmed ↗