Dietary Supplementation with Bacillus licheniformis and Bacillus subtilis Modulates Immunity, Serum Metabolome, and Intestinal Homeostasis in Cats
Meiting Zhang, Haocheng Xu, Tianfeng Zhang, Jia Kang, Zhihao Xu, Peng Wu, Yu Niu, Yonghao Shi, Yifan Zhong, Caimei Yang

TL;DR
This study shows that adding two types of probiotics to cat diets improves gut health, immunity, and reduces inflammation.
Contribution
The novel finding is that Bacillus licheniformis and Bacillus subtilis each uniquely modulate different metabolic pathways in cats.
Findings
Bacillus licheniformis improves fecal quality, modulates amino acid metabolism, and alters gut microbiota composition.
Bacillus subtilis enhances immune markers, reduces inflammation, and regulates lipid metabolism pathways.
Both probiotics increase IgA, IgM, and antioxidant levels while decreasing TNF-α in serum.
Abstract
The health of domestic cats, particularly gastrointestinal well-being, is garnering growing scientific interest, with probiotics recognized for their crucial role in sustaining gut homeostasis. This study investigates the impact of probiotics (Bacillus licheniformis, Bacillus subtilis) on feline immunity, serum metabolomics, and intestinal homeostasis. The results suggest that Bacillus licheniformis and Bacillus subtilis improve fecal quality, mitigate inflammatory responses, enhance immune function, and strengthen the intestinal barrier. Furthermore, Bacillus licheniformis modulates amino acid metabolic pathways, while Bacillus subtilis regulates lipid metabolic pathways. Together, these findings suggest that both strains hold promfecalas feed additives for improving feline gut health. In recent years, as the number of cats has increased, the intestinal health of cats has receied…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1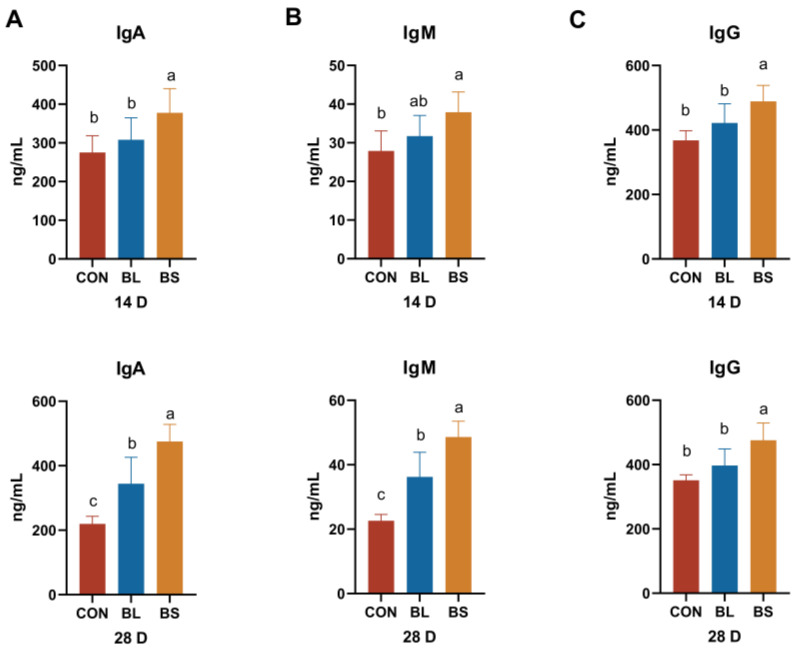 Figure 2
Figure 2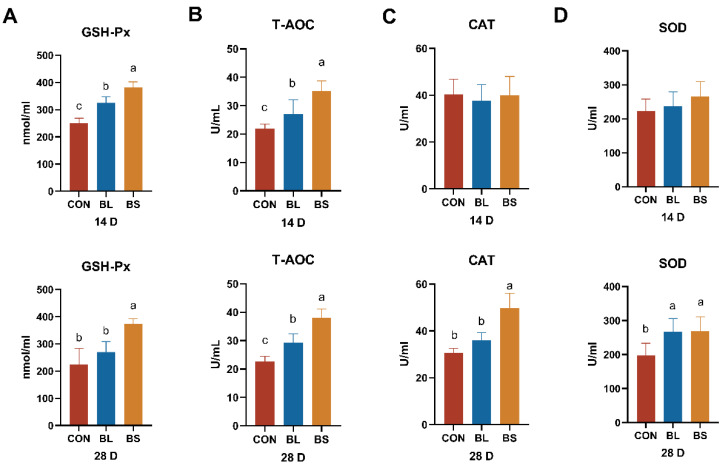 Figure 3
Figure 3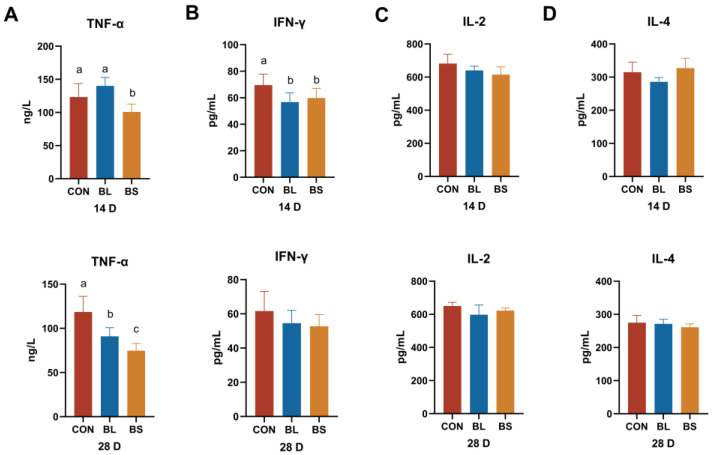 Figure 4
Figure 4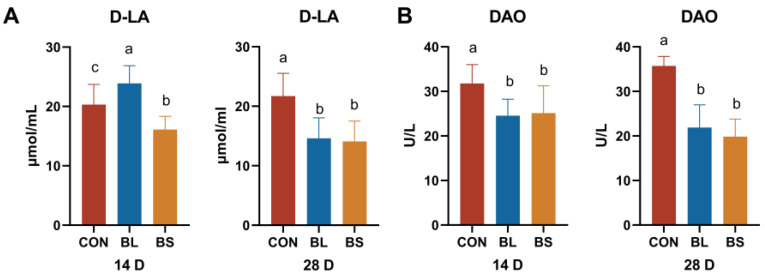 Figure 5
Figure 5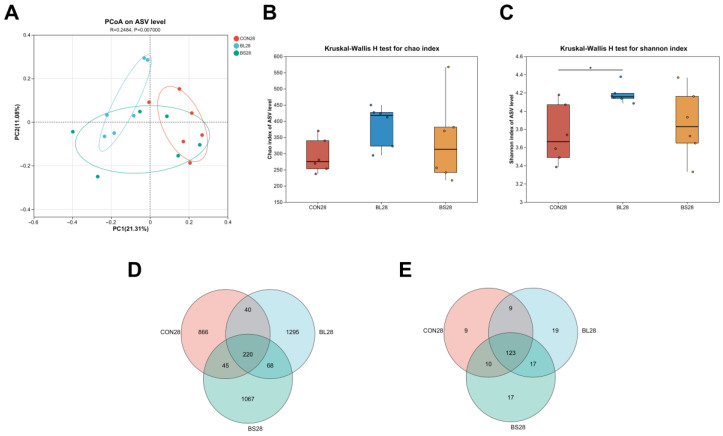 Figure 6
Figure 6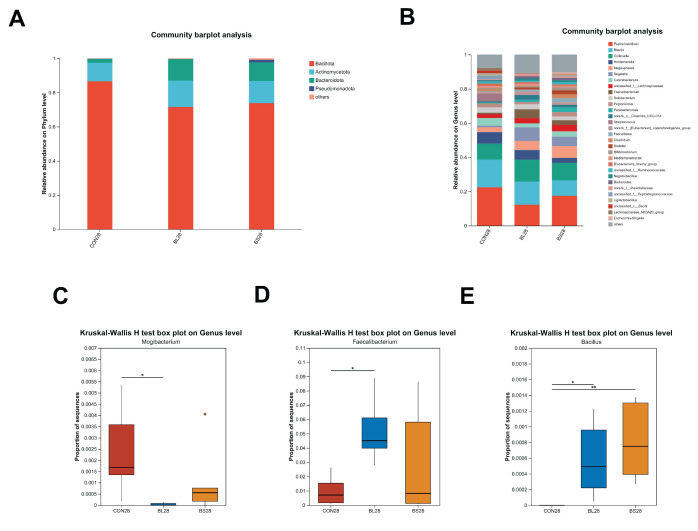 Figure 7
Figure 7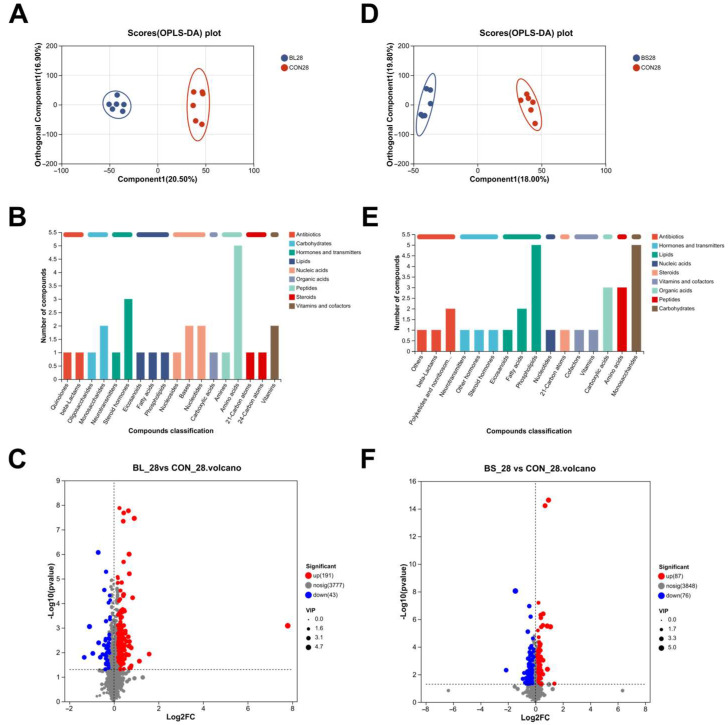 Figure 8
Figure 8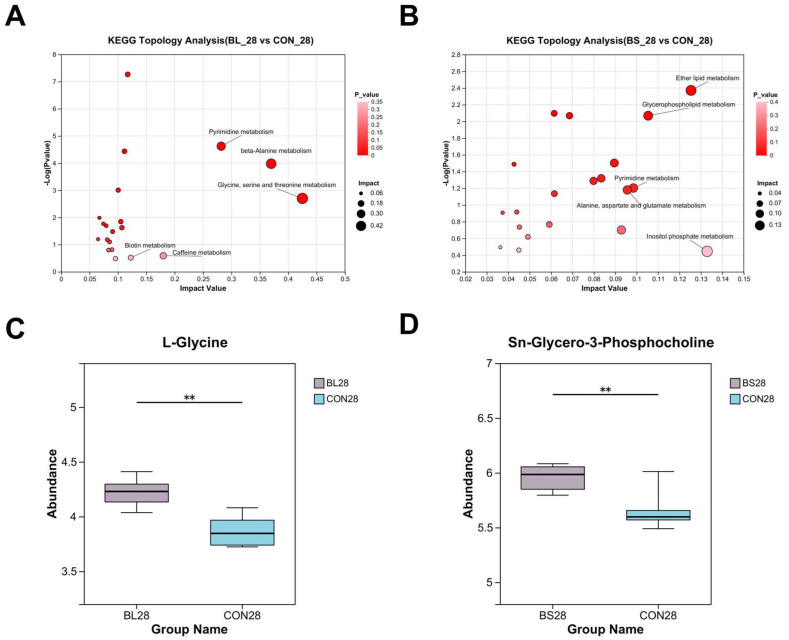 Figure 9
Figure 9Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGut microbiota and health · Probiotics and Fermented Foods · Biochemical effects in animals
1. Introduction
As dogs and cats occupy an increasingly important position in households, their health has garnered significant attention. Due to the rising ownership rate of cats, feline health has received increasing attention in recent years, driven by the rising number of cat owners. The gut microbiota, as an important part of the intestine, takes part in the digestion, absorption, and metabolism of nutrients [1]. A healthy and stable intestinal microbiota can resist inflammation and maintain the health of the host [2]. As a primary strategy for modulating gut microbiota, probiotics maintain the microecological homeostasis of the intestine by altering the structure of gut microbiota, thereby preserving the physical health of felines [3,4]. A study has found that chronic intestinal inflammation associated with feline inflammatory bowel disease (IBD) involves a sophisticated interplay between the mucosal immune system and gut microbiota in a host with genetic susceptibility [5]. A robust gut microbiome exhibits dual pro- and anti-inflammatory properties, maintaining homeostatic equilibrium to mitigate pathological hyperinflammation, while preserving immune responsiveness to infectious challenges [6]. The tripartite interplay governing probiotic–host cell–microbiota dynamics critically modulates host physiological homeostasis [7].
Probiotics, recognized as bioactive microorganisms conferring host health benefits, have been used as a functional food and dietary supplements, with emerging applications in canine and feline health management. Probiotics play a significant role in improving intestinal health primarily by regulating the balance of the gut microbiota, strengthening the intestinal barrier, and modulating immune responses [8]. Probiotics limit the colonization of pathogenic bacteria by competing for intestinal binding sites and nutritional resources, while concurrently enhancing the intestinal barrier function and reducing epithelial permeability [9]. B. licheniformis can form spores; its multiple benefits mainly focus on the gastrointestinal and immune systems. B. licheniformis alleviates the subhealth state through recovering the gut microbiota balance and reducing inflammation [10]. B. licheniformis BCG facilitated the digestion and absorption of nutrients in broilers, strengthened the intestinal physical barrier, and alleviated intestinal inflammation by altering microbial diversity and optimizing the microbiota structure. B. subtilis is environmentally stress-resistant due to its ability to form spores. It also synthesizes bioactive compounds—such as vitamins, enzymes, and short-chain fatty acids—that confer benefits to the host [11]. B. subtilis DE-CA9^TM^ intervention significantly ameliorated key serum oxidative stress markers, including advanced oxidation protein products (AOPP), diacron-reactive oxygen metabolites (d-ROMS), and thiobarbituric acid reactive substances (TBARS) [12]. Dietary supplementation with 0.01% B. subtilis C3102 enhanced canine intestinal health and fecal quality, evidenced by reduced ammonia concentration and improved fecal consistency [13]. Previous research conducted in our laboratory has demonstrated that B. licheniformis HJ0135 enhances immune responses, improves growth performance, and mitigates lipopolysaccharide (LPS)-induced inflammatory damage in weaned piglets [14]. However, research into B. Licheniformis in cats is still in its early stages.
Although B. licheniformis and B. subtilis have demonstrated beneficial effects on intestinal health, their applications and beneficial effects in feline species remain limited. Moreover, studies investigating the metabolomic effects in felines and the microbiota-metabolite interactions remain scarce. This study sought to elucidate how B. licheniformis and B. subtilis affect the feline gut microbiota and metabolome via multi-omics techniques, thereby demonstrating their potential functions in regulating intestinal health.
2. Materials and Methods
2.1. Animals and Experimental Treatments
Thirty-six healthy adult cats (Ragdoll cats) aged one year or older, (3.48 ± 0.71 kg) were randomly divided into three groups (n = 12 per group), and all cats were housed individually throughout the study. The feeding experiments were conducted for 35 days, comprising a 7-day adaptation period followed by a 28-day formal. The groups were assigned as follows: control group (CON), fed with the basal diet; B. licheniformis group (BL), provided with the basal diet supplemented with 1 × 10^10^ CFU/g of B. licheniformis; B. subtilis group (BS), served the basal diet supplemented with 1 × 10^10^ CFU/g of B. subtilis. Figure 1 illustrates the experimental design. The probiotics utilized in this study were supplied by Zhejiang Huijia Bio-technology Co., Ltd. (Huzhou, China). Before the trial, all cats received comprehensive immunization and deworming. All cats were free from medications known to affect gut microbiota for at least one month preceding the trial. Feces were collected twice daily. Every cat was supplied with 70 g of food once daily and had free access to clean drinking water at all times. The 28-day research phase was started after one week of acclimation. During the trial, body weight was assessed on days 1, 7, 14, 21, 28, and feed intake per day was noted to calculate the average daily feed intake (ADFI).
2.2. Sample Collection
A volume of 2 mL of blood was taken from the cephalic vein located in the forelimb of six randomly selected cats from each group on the 14th and 28th day. The serum was gathered by centrifugation at 3000 rpm for 15 min at 4 °C and preserved at −20 °C until further testing. Simultaneously, fresh feces were collected from six randomly selected cats from each group on day 28 of the experiment using sterilized forceps and transferred into a cryopreservation tube, then kept at −80 °C.
The fecal scoring was evaluated according to Table 1 on days 1–7, 14, and 28.
2.3. Serum Biochemical Analysis
Serum antioxidant indices, including superoxide dismutase (SOD), total antioxidant capacity (T-AOC), glutathione peroxidase (GSH-Px), catalase (CAT), D-lactic acid (DLA), and diamine oxidase (DAO) were detected using kits commercially available from the Ao Qing Biotechnology Co., Ltd. (Nanjing, Jiangsu, China).
2.4. Immune Cytokine Analysis
Serum concentrations of tumor necrosis factor-α (TNF-α), interferon-γ (IFN-γ), interleukin-2 (IL-2), interleukin-4 (IL-4), immunoglobulin (Ig)G, IgA, and IgM were quantified using enzyme-linked immunosorbent assay (ELISA). All assay kits were acquired from Ao Qing Biotechnology Co., Ltd. (Nanjing, Jiangsu, China).
2.5. Fecal Microbiome Analysis
Genomic DNA was extracted from fecal samples using the E.Z.N.A.^®^ Stool DNA Kit (Omega Bio-tek, Norcross, GA, USA). The V3–V4 hypervariable regions of the bacterial 16S rRNA gene were subsequently amplified by polymerase chain reaction (PCR) with the universal primers 338F (5′–ACTCCTACGGGAGGCAGCA–3′) and 806R (5′–GGACTACHVGGGTWTCTAAT–3′). PCR products were purified using the AxyPrep DNA Gel Extraction Kit and subjected to paired-end sequencing on an Illumina MiSeq PE300 platform (Illumina, San Diego, CA, USA). Quantification was performed with a Quantus Fluorometer (Promega, Madison, WI, USA). Raw sequencing data were demultiplexed and quality-filtered with Fastp (v0.20.0). Raw sequencing data were processed in QIIME 2 (version 2020.8; (https://qiime2.org, accessed on 6 March 2024)).
To perform taxonomic annotation of ASVs, the Naive Bayes was applied, and the abundance of each classified ASV was quantified across all samples. Alpha diversity was assessed via the Shannon and Chao indices to quantify both species richness and evenness within each sample. Principal Coordinate Analysis (PCoA) based on Bray–Curtis illustrated the similarity of communities. Bacterial abundance was calculated as a percentage of abundance at the phylum and genus taxonomic levels. The Kruskal–Wallis rank sum test was used to evaluate the significant differences in species abundance, and the species information with significant differences among multiple groups was obtained.
2.6. Analysis of Serum Metabolomics
Metabolite extraction was conducted by combining 100 μL of serum with acetonitrile/methanol (1:1, v/v). The samples were vortexed for 30 s and sonicated at low temperature for 30 min, then incubated at −20 °C for 30 min to induce protein precipitation. After centrifugation at 13,000 rpm and 4 °C for 15 min, the supernatant was removed and blown dry using nitrogen gas. The residue was redissolved in 100 μL of a 1:1 acetonitrile-water solution, sonicated at 4 °C for 5 min, and centrifuged once more at 13,000 rpm and 4 °C for 10 min. The resulting supernatant was transferred to injection vials for LC-MS/MS analysis, which was performed by Majorbio Bio-Pharm Technology Co., Ltd. (Shanghai, China) using a UHPLC-Orbitrap Exploris 240 system fitted with an ACQUITY UPLC HSS T3 column (100 mm × 2.1 mm, 1.8 μm; Waters, Milford, MA, USA).
Orthogonal projections to latent structures-discriminant analysis (OPLS-DA) and permutation tests for model validation were implemented using the R package “ropls” (version 1.6.2). Differential metabolites were chosen according to the thresholds of a variable importance in projection (VIP) score exceeding 2.0 and a p-value less than 0.05 from Student’s t-test. MetaboAnalyst 5.0 was utilized for pathway enrichment analysis, with significantly changed Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways detected via the hypergeometric test and adjusted for multiple comparisons (FDR < 0.05).
2.7. Statistical Analysis
SPSS software (version 27.0; SPSS Inc., Chicago, IL, USA) was utilized for the analysis of experimental data, with statistical significance set at p < 0.05. GraphPad Prism software (version 9; Dotmatics, Boston, MA, USA) was employed to create the graphics. Values of p < 0.05 (*) and p < 0.01 (**) were considered statistically significant and extremely significant, respectively. When comparing groups under the same indicator, significant differences are indicated by entries labeled with different letters (a, b, c) (p < 0.05).
3. Results
3.1. Growth Performance
Table 2 illustrates the effects of probiotics on feline body weights and ADFI throughout the course of the experiment. No significant difference was observed in the body weight and ADFI.
3.2. Fecal Scores
Fecal scoring is presented in Table 3. On days 3, 5, 7, 14, and 21, the cats in the BS group had higher fecal scores than those in the CON group (p < 0.05); meanwhile, the fecal scores of the BL group on days 4, 5 had a higher rating than the CON group.
3.3. Immunoglobulin Parameters
BS treatment significantly increased the level of the IgA, IgM, and IgG in the blood, as depicted on days 14 and 28 in Figure 2 (p < 0.05). The IgA and IgM concentrations in the BL group differed significantly from those in the CON group by day 28 (p < 0.05). However, the IgG of the BL group had no significant difference compared with the CON group.
3.4. Antioxidant Parameters
The effects of probiotics on antioxidant markers in feline serum were evaluated by quantifying the concentrations of CAT, SOD, GSH-Px, and T-AOC, and the relevant results are presented in Figure 3. Relative to the CON group, the BL and BS groups raised the level of GSH-Px and T-AOC on day 14 (p < 0.05). The BS group exhibited higher GSH-Px, T-AOC, CAT, and SOD on day 28 (p < 0.05). Additionally, the BL group had significantly elevated serum T-AOC and SOD levels on day 28 relative to the CON group. However, there were no significant differences in serum GSH-Px and CAT values in cats on day 28.
3.5. Inflammatory Factors
The impacts of the BL group and BS group on the serum cytokines are illustrated in Figure 4. Relative to the control group, the BS group exhibited a significant reduction in TNF-α and IFN-γ levels on day 14 (p < 0.05). In addition, IFN-γ levels in the BL group were significantly lower than those in the CON group on day 14 (p < 0.05). On day 28, TNF-α content in both the BL and BS groups was significantly lower than that in the CON group (p < 0.05).
3.6. Gut Barrier Function Parameters
Figure 5 depicts the serum levels of D-LA and DAO. In contrast to the CON group, the BL and BS group markedly reduced the serum concentrations of D-LA and DAO on day 28 (p < 0.05). Moreover, DAO levels had decreased in both the BS and BL groups, but DLA levels had decreased only in the BS group on day 14 (p < 0.05).
3.7. Fecal Microbiota Composition
Figure 6 presents the results of cat fecal microbiota diversity analysed using 16S rRNA sequencing technology. PCoA revealed distinct clustering of microbial communities according to group assignment, with clear separation observed among all three groups. (Figure 6A, p = 0.007). According to the Shannon index, the diversity of the CON group differed significantly from that of the BL group, but the Chao index revealed that there were statistically insignificant differences among the groups. (Figure 6B,C; p > 0.05). In order to characterize the taxonomic overlap across groups, an analysis was conducted of shared and unique taxa at both the amplicon sequence variants (ASVs) and genus levels. Across fecal samples from the three experimental groups, 3601 ASVs were identified in total at the ASV level. Among these, 220 ASVs were shared by all groups, while 1295 and 1067 ASVs were uniquely identified in the BL and BS groups, respectively. (Figure 6D). At the genus level, the common microbial genera of the CON, BL and BS groups were 123, and the unique genera of the three groups were 9, 19, and 17, respectively (Figure 6E).
Figure 7A,B illustrate the relative abundance of gut microbiota in cats, categorized by phylum and genus, respectively. At the phylum level, Bacillota represented the most abundant bacterial taxon in all groups, with initial relative abundances of 86.54% (CON group), 71.56% (BL group), and 73,78% (BS group) (Figure 7A). At the genus level, Peptoclostridium, Blautia, Collinsella, Holdemanella, and Megasphaera are the main dominant bacterial communities (Figure 7B). In addition, at the genus level, the relative abundance of Faecalibacterium and Bacillus was significantly increased in the BL group (Figure 7D; p < 0.05), while the relative abundance of Bacillus was significantly increased in the BS group (Figure 7E; p < 0.01). The relative abundance of Mogibacterium decreased significantly compared to the CON group after 28 days of feeding with BL (Figure 7C; p < 0.05).
3.8. Serum Metabolomics
Figure 8A illustrates the results of the cat serum metabolomics sequencing. Supervised mode identification of OPLS-DA showed that the experimental groups clustered distinctly. As shown in Figure 8A,D, the addition of BL and BS led to a clear separation between these two groups and the CON group. This result indicates that the metabolic characteristics of the two groups differ significantly from those of the control group. The KEGG total compound classification statistics chart shows that the BL group consists primarily of peptides, hormones, and transmitters (Figure 8B), whereas the BS group consists primarily of carbohydrates and lipids (Figure 8E). As shown in the volcano plot, it presents the expression levels of metabolites that were differentially expressed after the addition of BL. In total, 191 metabolites were significantly upregulated and 43 were downregulated in the BL group relative to the control group (Figure 8C). Following the addition of BL, 87 metabolites exhibited significant upregulation and 76 showed significant downregulation (Figure 8F).
The KEGG Topology Analysis of BL and BS Group Serum is displayed in Figure 9A,B. The results showed that the abundance of the synthesis pathways for Pyrimidine metabolism, β-Alanine metabolism, Glycine, serine and threonine metabolism, and Biotin metabolism increased in the BL group (Figure 9A). Furthermore, the BS group exhibited increased activity in the synthesis pathways of Ether Lipid metabolism, Pyrimidine metabolism, Alanine, aspartate and glutamate metabolism, Glycerophospholipid metabolism, and Inositol phosphoric acid metabolism (Figure 9B). Among the identified differential metabolites, the level of L-Glycine in the BL group was found to be significantly higher compared with that in the control group (Figure 9C; p < 0.05), while sn-Glycero-3-Phosphocholine was detected at a significantly higher level in the BS group (Figure 9D; p < 0.05).
4. Discussion
As cats become more popular as pets, greater attention is being paid to their health issues, such as diarrhea, obesity, and urinary problems. The gut is crucial for animal health, especially the microbes in it, which affect how nutrients are absorbed and metabolized [4]. Probiotics, which are beneficial live microorganisms, offer several advantages, including a high safety profile, non-toxicity, absence of residual accumulation, and minimal environmental impact [3]. The concentration of probiotics required for a clinical effect is often quoted as being at least 10^6^ CFU/mL in the small bowel and at least 10^8^ CFU/g in the colon [16]. Guo confirmed that a high-dose probiotic (Bacillus spp., Bifidobacterium spp., Clostridium butyricum, Lactobacilli spp., Lactococcus spp., Leuconostoc cremoris, Saccharomyces spp., or Streptococcus spp.) regimen demonstrates superior therapeutic efficacy over low-dose administration in managing antibiotic-associated diarrhea [17]. This finding aligns with the dose–effect threshold reported by Ouwehand [18]. Consequently, the probiotic dosage administered in this study was established at 1 × 10^10^ CFU/day, selected based on previous dose-response studies. Probiotic supplementation demonstrates therapeutic potential in enhancing intestinal homeostasis and reducing diarrheal incidence [19]. Torres-Henderson and colleagues found that the Enterococcus faecium strain SF68 reduced fecal scores and ameliorated diarrhea in their study population [20]. The supplementation of BL and BS in the present study significantly improved fecal scores in cats, which aligns with previous findings.
IgA represents the predominant immunoglobulin isotype across mucosal surfaces, serving as the primary immunological barrier against pathogenic microbial colonization and invasion. The present study has demonstrated that dietary supplementation with BL significantly enhances serum IgA levels in juvenile pigs [21]. IgM is a primary antibody generated during the initial immune response to antigen exposure. The substance exhibits potent bactericidal activity, robust complement system activation, and immunomodulatory functions [22]. IgG is the predominant antibody found in serum, being particularly prevalent in blood and other extracellular fluids [23]. The results showed that adding BL and BS to the diet of broilers significantly increased their serum IgA and IgM levels, which suggests that the broilers’ humoral immune capacity was enhanced [24]. Dong demonstrated that dietary supplementation with Bacillus subtilis BYS2 significantly increased serum IgG levels in broilers, with a 31.6% elevation compared to control-group counterparts [25]. Consistent with established literature, the current study demonstrated that BL and BS supplementation significantly elevated serum IgA and IgM concentrations in broilers. In contrast, BL administration failed to induce statistically significant increases in IgG levels, potentially attributable to the limited duration of the experimental intervention.
Serum antioxidant enzymes, including GSH-Px, T-AOC, SOD, and CAT, constitute critical elements of the endogenous antioxidant defense system. Acting in coordination, these enzymes mitigate oxidative harm through the neutralization of excess reactive oxygen species (ROS), a process that helps sustain cellular redox homeostasis [26]. SOD is an important endogenous antioxidant enzyme that acts as part of the body’s first line of defense against ROS [27]. Moreover, the glutathione system plays a pleiotropic role in defending cells against metabolic, oxidative, and metal stresses [28]. Lei observed that BS supplementation enhanced the antioxidant capacity of laying hens [29], and Bai revealed that Bacillus subtilis fmbJ (CGMCCN 0943) could increase the antioxidant capacity of broiler chickens by increasing GSH-Px and SOD activities in serum [30]. Consistent with previous research, we observed that BS increased GSH-Px, SOD, CAT, and T-AOC activities, and BL increased GSH-Px and T-AOC.
TNF-α is a pro-inflammatory cytokine that plays a central role in modulating innate immunity [31,32]. IFN-γ is crucial for maintaining tissue homeostasis, immune responses, and inflammatory reactions [33]. IL-2 is a crucial cytokine that regulates immune homeostasis and promotes T-cell activation, proliferation, and differentiation [34]. Dietary probiotic supplementation significantly reduced serum TNF-α concentrations in weaned piglets compared to the CON group, demonstrating an anti-inflammatory effect [35]. Research has revealed that the supplementation of Bacillus licheniformis MCC 2514 and Bifidobacterium breve NCIM 5671 downregulates the expression of pro-inflammatory cytokines while upregulating anti-inflammatory cytokines, including IL-4 [36]. Previous research conducted in our laboratory indicates that BL supplementation upregulates anti-inflammatory cytokine IL-4 and downregulates pro-inflammatory TNF-α levels [37]. In alignment with prior research, our study observed a reduction in pro-inflammatory cytokines, such as TNF-α and IFN-γ. Nevertheless, there was no substantial alteration in the concentrations of IL-4 and IL-2. This could be due to the inadequate dosage and duration of the experiment.
The gastrointestinal tract represents a major immunological organ, playing an essential role in host defense against pathogens. Probiotic colonization in the intestinal lumen modulates gut microbiota composition, thereby enhancing intestinal homeostasis and immune function [38]. D-LA and DAO serve as complementary biomarkers of intestinal barrier integrity, with their combined assessment providing a more robust evaluation of intestinal health status [39]. Yang demonstrated that Bacillus subtilis HH2 supplementation significantly reduced serum DAO activity following enterotoxigenic Escherichia coli (ETEC) challenge in beagles, concomitant with the amelioration of diarrheal symptoms [40]. Adding Bifidobacterium lactis (B. lactis) and Lactobacillus plantarum significantly improved intestinal barrier function. Plasma D-LA concentration decreased by 30.38%, as did DAO concentration (by 22.68%) [41]. The current study demonstrated that both BL and BS supplementation significantly reduced serum D-LA concentrations and DAO activity compared to control groups, consistent with previous reports of their intestinal barrier-protective effects.
Specifically, Fusobacteria, Bacteroidetes, and Firmicutes were identified as the dominant and most prevalent bacterial phyla in the fecal microbiota of both dogs and cats [42,43]. The present study identified that the dominant phyla within the intestinal microbiota are Bacillota, Actinomycetota, Bacteroidota, and Pseudomonadota, which differ from previous reports. This discrepancy may be due to host-specific factors, such as breed (predominantly Ragdoll cats) and age (>1 year). The commensal bacterium Faecalibacterium prausnitzii plays a key role in IBD pathogenesis, and Butyrate mediates the anti-inflammatory effects of Faecalibacterium prausnitzii in intestinal epithelial cells through Dact3 [44]. Previous studies have suggested that Butyrate exerts potent effects on a variety of colonic mucosal functions, such as inhibition of inflammation and carcinogenesis, reinforcing various components of the colonic defense barrier and decreasing oxidative stress [45]. Given that Faecalibacterium is a major butyrate producer, it could increase intestinal butyrate levels by modulating beneficial microbiota. This would contribute to improved intestinal barrier function and overall gut homeostasis. Microbial compositional analysis revealed that the BL increased the relative abundance of Faecalibacterium, and the BL and BS increased the relative abundance of Bacillus, while the Mogibacterium relative abundance was decreased. Prior research has indicated that Bacillus hs a protective effect and performs functions that are important for sustaining gut health [46]. Previous studies have established that the relative abundance of Mogibacterium within the microbiome of patients with colorectal cancer (CRC) increases progressively from mucosal-invasive carcinoma to advanced stages [47]. M Oba demonstrated that B. subtilis ATCC PTA-122264 significantly reduced fecal abundance of potential pathogens, including Streptococcus spp. Escherichia coli, and Cyanobacteria, suggesting its antimicrobial efficacy in modulating gut microbiota composition [48]. Consistent with previous findings, BL and BS improve the health of the intestinal tract by increasing the levels of beneficial bacterial taxa such as Faecalibacterium and Bacillus, and decreasing the levels of detrimental bacteria, including Mogibacterium. Within this study, the BL group exhibited a total of 3777 differential metabolites relative to the control group. The primary annotated metabolic pathways included Pyrimidine metabolism, beta-Alanine metabolism, and Glycine, serine, and threonine metabolism. Notably, the level of the metabolite L-Glycine in the BL group was significantly upregulated. Similarly, the BS group showed 3848 differential metabolites, with major pathways enriched in Lipid metabolism, Pyrimidine metabolism, Glycerophospholipid metabolism, and Alanine, aspartate, and glutamate metabolism. Likewise, the level of sn-Glycero-3-Phosphocholine in the ether lipid metabolism pathway of the BS group showed an upregulated trend. The distinct metabolic reprogramming induced by BL (amino acid metabolism emphasis) versus BS (Lipid metabolism focus) suggests strain-specific bioactivity. Sn-Glycero-3-Phosphocholine serves as a critical biosynthetic precursor for phosphatidylcholine (PC). Studies have underscored the importance of PC metabolism, in which the balance between its anabolic and catabolic processes is essential for preserving the structural integrity of cellular membranes, supporting hepatic function, and ensuring proper neurological activity [49]. Through 16S rRNA gene sequencing analysis, both BL and BS supplementation increased the abundance of beneficial bacteria, including Faecalibacterium. As a butyrate producer, Faecalibacterium contributes to the suppression of inflammation and reinforcement of the intestinal barrier. Concurrently, sn-Glycero-3-Phosphocholine, a precursor for phospholipid membrane synthesis, provides essential substrates for the regeneration of intestinal cells. Together, these effects create a low-inflammatory microenvironment with enhanced barrier integrity, which in turn facilitates the colonization and expansion of beneficial bacteria. These findings are consistent with the observed attenuation of inflammatory markers and the promotion of a beneficial microbiota composition. Research has demonstrated that L-Glycine exhibits significant potential in mitigating inflammatory responses, modulating immune functions, and preserving cellular viability [50,51]. These results align with those of prior studies, confirming that BL supplementation effectively lowers serum TNF-α levels and mitigates inflammatory responses. Glycine is a constituent of significant proportions in the proteins of both gram-positive and gram-negative gut bacteria [52]. This result suggests that glycine has a crucial function in fostering the optimal proliferation of the intestinal microbiota [53]. Our research revealed that elevated serum L-glycine levels, alongside an increased abundance of Faecalibacterium and Bacillus, may be the result of L-Glycine’s ability to maintain intestinal homeostasis, encourage the growth of beneficial bacteria, and strengthen the intestinal functional barrier.
5. Conclusions
In conclusion, dietary supplementation of BL and BS can improve the immune ability of cats by increasing the content of IgA and IgM in serum, and it has significant antioxidant capacity and potent inflammatory protection. Additionally, the supplementation of BL and BS can elevate the abundance of beneficial bacterial taxa while lowering the abundance of detrimental bacteria. This strategy improves the health of the intestinal tract by adjusting the structure of the gut microbiota. Analytical findings indicated that the differentially abundant metabolites in the BL group were primarily tied to amino acid metabolism pathways, whereas those in the BS group were mainly related to lipid metabolism. Despite these findings, this study has limitations. The sample was restricted to Ragdoll cats, warranting future investigations across diverse breeds to comprehensively assess the health impacts of BL and BS. Additionally, the 35-day experimental period captured only short-term physiological responses; extended studies are needed to evaluate long-term effects. Collectively, these results suggest that BL and BS possess considerable potential for applications in intestinal health maintenance.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Markowiak P. Śliżewska K. Effects of probiotics, prebiotics, and synbiotics on human health Nutrients 20179102110.3390/nu 909102128914794 PMC 5622781 · doi ↗ · pubmed ↗
- 2Alessandri G. Argentini C. Milani C. Turroni F. Ossiprandi M.C. Sinderen D.V. Ventura M. Catching a glimpse of the bacterial gut community of companion animals: A canine and feline perspective Microb. Biotechnol.2020131708173210.1111/1751-7915.1365632864871 PMC 7533323 · doi ↗ · pubmed ↗
- 3Zha M. Zhu S. Chen Y. Probiotics and cat health: A review of progress and prospects Microorganisms 202412108010.3390/microorganisms 1206108038930462 PMC 11205573 · doi ↗ · pubmed ↗
- 4Lee D. Goh T.W. Kang M.G. Choi H.J. Yeo S.Y. Yang J. Huh C.S. Kim Y.Y. Kim Y. Perspectives and advances in probiotics and the gut microbiome in companion animals J. Anim. Sci. Technol.20226419721710.5187/jast.2022.e 835530406 PMC 9039956 · doi ↗ · pubmed ↗
- 5Jergens A.E. Feline idiopathic inflammatory bowel disease J. Feline Med. Surg.20121444545810.1177/1098612 X 1245154822736679 PMC 10822384 · doi ↗ · pubmed ↗
- 6Tizard I.R. Jones S.W. The microbiota regulates immunity and immunologic diseases in dogs and cats Vet. Clin. N. Am. Small Anim. Pract.20184830732210.1016/j.cvsm.2017.10.00829198905 · doi ↗ · pubmed ↗
- 7Li Y. Ali I. Lei Z. Li Y. Yang M. Yang C. Li L. Effect of a multistrain probiotic on feline gut health through the fecal microbiota and its metabolite SCF As Metabolites 20231322810.3390/metabo 1302022836837847 PMC 9962843 · doi ↗ · pubmed ↗
- 8Mohammed E.A.H. Ahmed A.E.M. Kovács B. Pál K. The Significance of Probiotics in Aquaculture: A Review of Research Trend and Latest Scientific Findings Antibiotics 20251424210.3390/antibiotics 1403024240149052 PMC 11939380 · doi ↗ · pubmed ↗