Novel Silent Mutations in the HIRA Gene Associated with Litter Size in Sonid Sheep
Chen Wang, Zhana Naren, He Bu, Ming Cang, Guifang Cao, Buhe Nashun, Bin Tong

TL;DR
This study found new mutations in the HIRA gene linked to litter size in Sonid sheep, offering potential genetic markers for improving sheep breeding.
Contribution
The study identifies 15 novel HIRA gene mutations in Sonid sheep, some of which are associated with litter size and predicted to affect protein structure.
Findings
Three HIRA gene mutations (c.1521C>G, c.1572C>T, c.1578G>A) are significantly associated with litter size in Sonid sheep.
Two mutations (c.1273G>A and c.1735A>G) are predicted to alter the protein structure of HIRA.
The mutations may enhance mRNA stability by lowering the minimum free energy of the secondary structure.
Abstract
This study investigated the association between genetic variants of the HIRA gene and litter size in Sonid sheep. Through analysis, we identified fifteen novel mutations in the HIRA gene. Among these, the c.1521C>G, c.1572C>T, and c.1578G>A variants were associated with litter size, while c.1273G>A and c.1735A>G were predicted to alter protein structure. These variants could serve as valuable genetic markers for improving prolificacy in sheep breeding and highlight new variants for further functional research on the HIRA gene. Improving ovine reproductive efficiency is an important breeding goal that could substantially enhance economic viability in the sheep industry. The histone cell cycle regulator (HIRA) gene has been functionally validated as a reproductive regulator in model organisms such as Drosophila and murine species; however, research on the effects of HIRA on the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
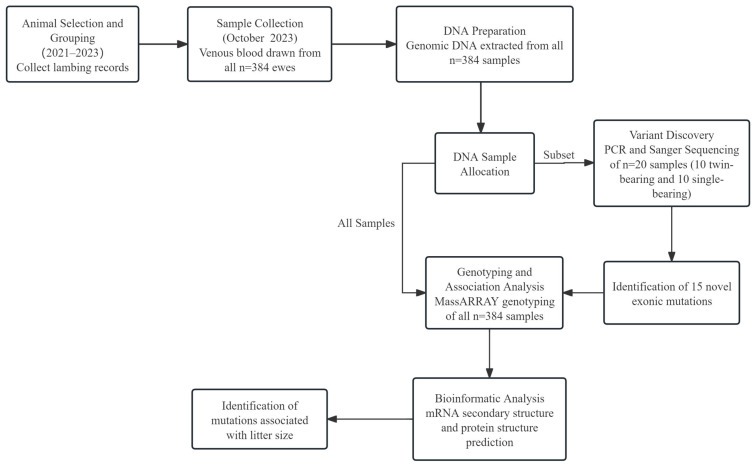 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3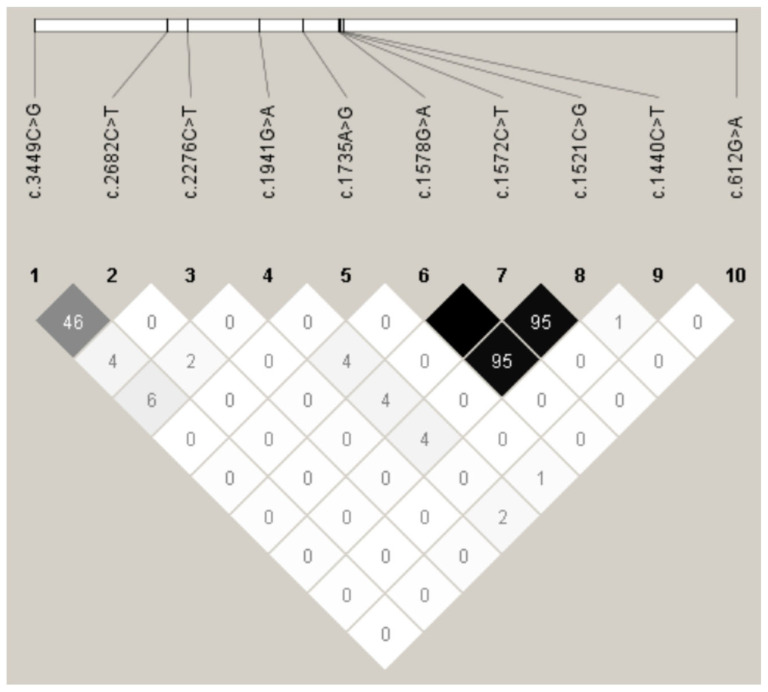 Figure 4
Figure 4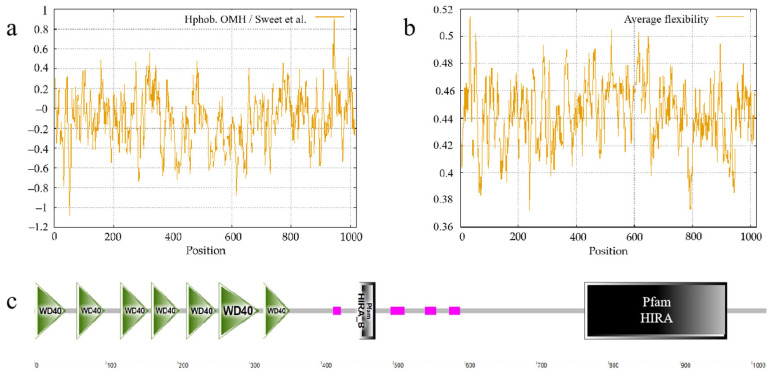 Figure 5
Figure 5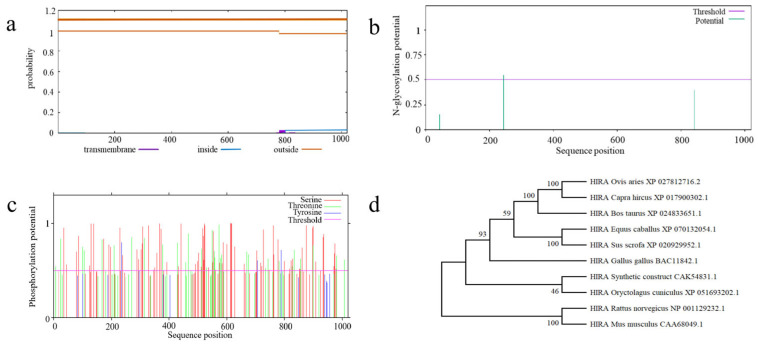 Figure 6
Figure 6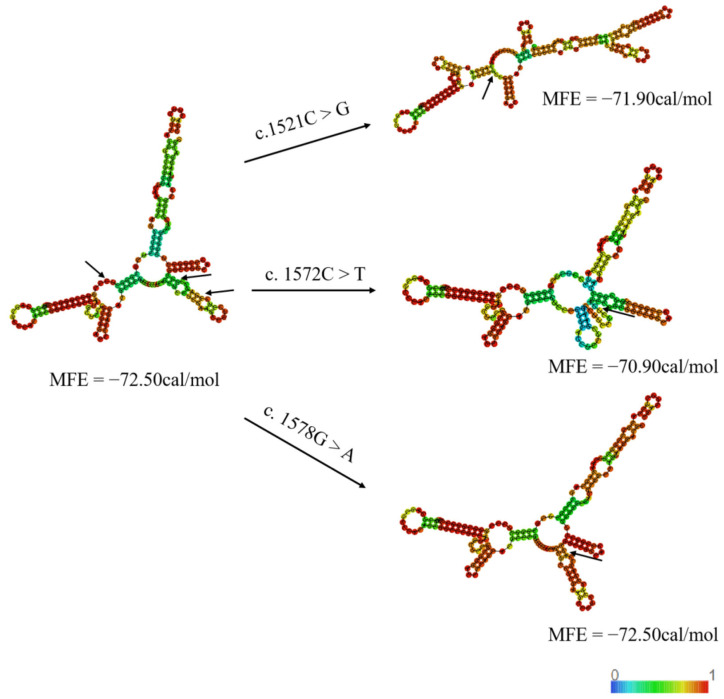 Figure 7
Figure 7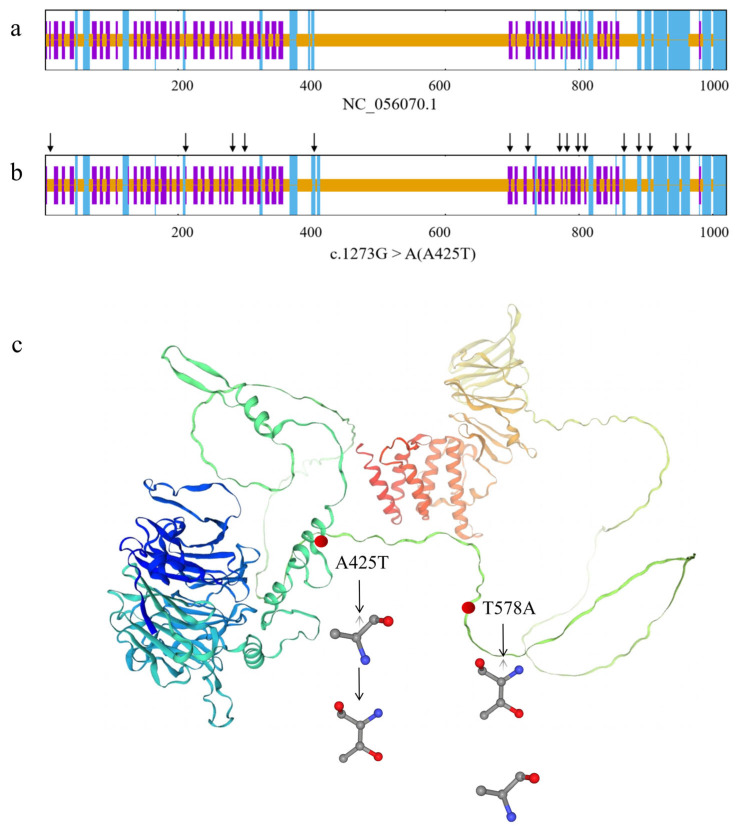 Figure 8
Figure 8- —Central Guidance for Local Scientific and Technological Development Plan
- —Major Science and Technology Projects
- —Major Demonstration Projects for Technological Innovation
- —Xilingol Science & Technology Plan
- —State Key Laboratory for Reproductive Regulation and Breeding of Grassland Livestock
- —Ministry of Human Resources and Social Security of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCancer-related molecular mechanisms research · Genetic and phenotypic traits in livestock · RNA modifications and cancer
1. Introduction
Sheep is one of the most important livestock species globally, with a world inventory of over 1.3 billion head, contributing significantly to meat, wool, and milk production. China has the world’s largest sheep inventory, exceeding 170 million, with Inner Mongolia being a key production region. Sheep husbandry is a cornerstone of the agricultural economy and social fabric of Inner Mongolia [1,2]. Improving the reproductive efficiency of local breeds like Sonid sheep is a high-priority goal for enhancing food security and herders’ livelihoods in this region.
Sonid sheep is a local, indigenous, coarse wool breed found in Inner Mongolia, China, predominantly distributed across the Sonid grassland of the Mongolian Plateau under arid continental climatic conditions [3]. The meat of Sonid sheep is very popular among consumers in Inner Mongolia and even all of China, since it is palatable and recognized as a natural green food [2,3]. However, the production rate of Sonid sheep is constrained by seasonal estrus and low fertility (mean litter size is 1.03~1.13). Currently, the three primary genes recognized as influencing sheep prolificacy are bone morphogenetic protein receptor 1B (BMPR1B), bone morphogenetic protein 15 (BMP15), and growth differentiation factor 9 (GDF9). They all belong to the transforming growth factor β (TGF-β) superfamily, located on the 6, X, and 5 chromosomes of sheep, respectively. Mutations in these genes can influence the ovulation rate and litter size of ewes [3,4,5]. The FecB mutation of BMPR1B, which was first discovered in Booroola Merino sheep, is the main gene responsible for high prolificacy in sheep. These genes that influence the ovulation rate and litter size have been confirmed across various sheep populations worldwide. While mutations in BMPR1B, BMP15, and GDF9 are major well-established genes in terms of prolificacy, their effects are breed-specific, and they do not account for all genetic variation in litter size. Therefore, investigating novel candidate genes, such as HIRA, which has been implicated in reproductive processes in other species, is crucial for improving prolificacy in Sonid sheep. To date, there have been no studies looking into the prolificacy of Sonid sheep. Thus, it is important to investigate breed-specific functional genes and markers associated with prolificacy in Sonid sheep.
Histone cell cycle regulator (HIRA) is a conserved histone chaperone for H3.3 [6], comprising 24 exons encoding 1020 amino acids that participate in chromatin regulatory processes, including transcriptional regulation, spermatid chromatin remodeling, and embryonic development [7,8]. As a member of an evolutionarily conserved protein family, HIRA orthologs have been identified across diverse organisms, including yeast, Drosophila, mice, human, and plants, playing essential roles in maintaining genomic integrity and cellular homeostasis [9,10]. In Drosophila models, HIRA deficiency leads to female sterility, in which maternal histones fail to incorporate into the male pronucleus during fertilization, thereby preventing paternal chromatin reorganization [11]. Vertebrate studies further revealed that HIRA mutations universally result in embryonic lethality [12]. Specifically, murine studies by Nashun et al. established that murine HIRA is indispensable for transcriptional regulation and DNA methylation in oocytes. Its absence disrupts H3.3/H4 histone replacement, thus inducing chromatin abnormalities and subsequent oocyte apoptosis [13]. In sheep, Zhou et al. demonstrated that two single-nucleotide mutations (SNPs), g.71874104G>A and g.71833755T>C in the HIRA gene, significantly influence litter size in Small-tail Han sheep [14], thus highlighting the possibility that the HIRA gene could be considered a candidate gene for reproductive performance in sheep.
Based on the established role of HIRA in reproduction and previous associations in other sheep breeds, we hypothesized that novel genetic variants in the HIRA gene are associated with litter size in Sonid sheep. Therefore, this study aims to achieve several objectives: (1) identify novel mutations in the HIRA gene of Sonid sheep; (2) analyze the association between the novel mutations of the HIRA gene and litter size in Sonid sheep; (3) assess the potential effects of these novel variants on the mRNA and protein characteristics and structure of the HIRA gene in sheep. Our findings may contribute to the development of effective marker-assisted selection strategies to enhance litter size in the Sonid sheep breed and offer new perspectives on the influence of the HIRA gene on reproductive traits in sheep.
2. Materials and Methods
2.1. Samples and Date
A total of 384 healthy, multiparous Sonid sheep ewes (aged 3–4 years, body weight 45–55 kg, with 1–2 previous lambings) were sampled from the Sonid Right Banner Sonid Sheep Breeding Technology Company and the Sonid Left Banner Livestock Germplasm Development Company in Inner Mongolia for association analysis. Sonid sheep is a breed characterized by seasonal estrus in autumn [1]. Based on lambing records from these companies from 2021 to 2023, we sampled 52 Sonid ewes that consistently produced twin litters over three consecutive years for genotyping while sampling 332 ewes that stably produced a single lamb. Blood samples of 384 sheep were collected in October 2023, then immediately placed on ice after collection, transported to the laboratory within 6 h, and stored at −80 °C until DNA extraction. To discover novel variants in the HIRA gene, a subset of 20 ewes (10 from the twin-lamb group and 10 from the single-lamb group) was randomly selected for sequencing. It is important to emphasize that these ewes were mated naturally without any selection of rams. All the sheep were maintained in similar environments with unrestricted access to food and water. Given the impact of the FecB mutation on the litter size in sheep, we initially screened the Sonid ewes for the presence of the FecB mutation, and our experimental animals did not exhibit any FecB mutation.
2.2. DNA Extraction and Sequencing
Twenty ewes were selected for the assessment of variants in the HIRA gene among Sonid sheep, consisting of ten ewes that consistently produced twin lambs and ten ewes that regularly had single lambs. DNA was extracted from 384 blood samples using the Tiangen Blood/Cell/Tissue Genomic DNA Extraction Kit (Tiangen, Beijing, China), in accordance with the instructions provided by the manufacturer. To avoid degradation, the quality and concentration of the extracted DNA were evaluated through agarose gel electrophoresis and with a Nanodrop One spectrophotometer (from Thermo, Waltham, MA, USA) after being stored at −20 °C.
For PCR amplification, 21 primer pairs were designed using Primer Premier 5.0 to target the exon regions of the ovine HIRA gene (Oar_rambouillet_v3.0, NCBI reference sequence: NC_056070.1), as outlined in Table 1 [2]. The total volume of each PCR is 50 μL, which includes 2 μL of the extracted DNA as a template, 1 μM of each primer, 25 μL of Ex Taq DNA polymerase (from Takara, Dalian, China), and 21 μL of ddH2O. The cycling conditions for the PCR were set as follows: an initial denaturation at 94 °C for 5 min, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing for 30 s, and elongation at 72 °C for 1 min and 10 s, with a final extension at 72 °C for 10 min. The specific annealing temperatures for each PCR fragment are listed in Table 1. The quality and quantity of the PCR products were analyzed through 3.0% agarose gel electrophoresis. These products were subsequently sequenced by the Beijing Genomics Institute (BGI, Beijing, China).
2.3. SNP Genotyping Using iPLEX MassARRAY
Fifteen new variants were analyzed using the MassARRAY^®^ SNP genotyping system (Agena Bioscience, San Diego, CA, USA) in the experimental samples. The PCR and extension primers of the HIRA gene were designed from sequences encompassing each target mutation along with approximately 100 bases upstream and downstream, employing the Assay Design Suite v3.0 (http://agenabio.com/assay-design-suite-20-software, accessed on 10 October 2024) with default settings (Supplementary Table S1). The genotypes for each allele were assessed using the Sequenom MassARRAY iPLEX platform. The resulting data were processed with MassARRAY Typer 4.0 Analyzer software (Agena Bioscience, San Diego, CA, USA) [15].
2.4. Bioinformatic Analyses
The fundamental characteristics of the predicted HIRA proteins were analyzed using ProtParam (https://web.expasy.org/protparam/, accessed on 2 December 2024). Potential transmembrane domains were identified through TMHMM (https://services.healthtech.dtu.dk/services/TMHMM-2.0/, accessed on 4 December 2024). The Simple Molecular Architecture Research Tool (SMART) (http://smart.embl-heidelberg.de/, accessed on 5 December 2024) was employed to predict the conserved domains of HIRA. N-glycosylation and phosphorylation sites were predicted with NetOGlyc 4.0 (https://services.healthtech.dtu.dk/services/NetOGlyc-4.0/, accessed on 12 December 2024) and NetPhos 3.1 (https://services.healthtech.dtu.dk/services/NetPhos-3.1/, accessed on 13 December 2024), respectively. The hydrophilicity and average flexibility index of the ovine HIRA amino acid sequences were evaluated using ProtScale (https://web.expasy.org/protscale/, accessed on 3 January 2025). RNAfold was utilized to forecast the mRNA secondary structure of the HIRA gene in sheep (http://rna.tbi.univie.ac.at//cgi-bin/RNAWebSuite/RNAfold.cgi, accessed on 4 January 2025). The secondary structure of ovine HIRA was determined with SOPMA (https://npsa.lyon.inserm.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html, accessed on 4 January 2025), while its tertiary structure and template alignment were projected using SWISS-MODEL (https://swissmodel.expasy.org/, accessed on 7 January 2025). Multiple alignments and the construction of a molecular phylogenetic tree were carried out using Uniprot Align (https://www.uniprot.org/align, accessed on 10 January 2025) and MEGA-X (MEGA 11.0.13, https://www.megasoftware.net/docs, accessed on 15 January 2025). A schematic diagram of the experimental protocol is provided in Figure 1.
2.5. Statistical Analyses
Genotypic and allelic frequency calculations, along with Hardy–Weinberg equilibrium assessments, were performed for the Sonid sheep population. Population genetic metrics were derived using Nei’s methods [16], which included observed heterozygosity (H_o_), expected heterozygosity (H_e_), effective allele numbers (n_e_), and polymorphism information content (PIC). A chi-squared test was applied to compare the allelic frequencies of each variant. Linkage disequilibrium (LD), which encompassed D’ and r^2^ values, was evaluated using HAPLOVIEW version 4.2 [17]. The genetic effects of each SNP on the litter size of Sonid sheep were analyzed using a one-way analysis of variance (ANOVA) followed by Tukey’s multiple comparison test [18].
3. Results
3.1. Variant Discovery in HIRA
Through direct sequencing analysis, we identified 15 novel variants exclusively localized within the exonic regions of the HIRA gene, designated as the c.612G>A, c.1179C>T, c.1206G>A, c.1226A>G, c.1273G>A, c.1440C>T, c.1521C>G, c.1572C>T, c.1578G>A, c.1735A>G, c.1941G>A, c.2276C>T, c.2499G>A, c.2682C>T, and c.3449C>G mutations. The genomic distribution of these variants across the HIRA gene is schematically illustrated in Figure 2a,b.
Missense mutations were identified in the c.1273G>A (Ala425Thr) and 1735A>G (Thr579Ala) mutations of the HIRA gene. In contrast, all other variants showed synonymous changes. The HIRA protein aligns with a comparison of wild-type sequences from various species ranging from chicken to mammals, as illustrated in Figure 3.
3.2. Genetic Diversity Analysis
Allele frequencies, genotype distributions, and genetic indices (H_o_, H_e_, n_e_, PIC, and Hardy–Weinberg equilibria) for the identified variants in the Sonid sheep population are summarized in Table S2. Moderate polymorphism was detected at the c.1941G>A mutation, while all other variants displayed low polymorphism.
3.3. Linkage Disequilibrium Analysis of Novel Variants in HIRA
Linkage disequilibrium (LD) analysis was performed for ten mutations that met the Hardy–Weinberg equilibrium (HWE) criteria. The remaining five identified variants were excluded from the LD analysis because they exhibited significant departure from HWE (p < 0.05), which can complicate the accurate estimation of haplotype frequencies and LD patterns. D’ and r^2^ values were also calculated for Sonid sheep. The analysis revealed near complete linkage disequilibrium (D’ = 1.0; r^2^ = 1.00) between c.1572C>T and c.1578G>A. Strong LD was also observed between c.1572C>T and c.1521C>G (D’ = 1.00; r^2^ = 0.953), as well as between c.1578G>A and c.1521C>G (D’ = 1.00; r^2^ = 0.954). Therefore, these mutations are designated as LD-Sonid for combined locus analysis. The remaining LD patterns are graphically represented in Figure 4.
3.4. Associations Between Genetic Variants and Litter Size
Association analyses were conducted in 384 Sonid ewes for seven variants. Among the fifteen identified mutations, five exhibited significant departures from HWE, and three were found in fewer than ten individuals. Consequently, these eight variants were excluded from the statistical computations. As the c.1521C>G, c.1572C>T, and c.1578G>A mutations were in LD (Figure 4), the association results for the combined LD-Sonid are represented by the c.1521C>G genotype in Table 2. The c.1521C>G, c.1572C>T, and c.1578G>A of LD-Sonid in exon 14 of HIRA were associated with litter size in Sonid sheep (p < 0.01). Detailed results are presented in Table 2.
3.5. Bioinformatic Analysis of Ovine HIRA
3.5.1. Structural Characterization of Ovine HIRA Protein
Hydrophobicity profiling identified peak hydrophobicity (0.904) at residue 945 and minimal hydrophobicity (−1.084) at residue 53 (Figure 5a). Flexibility analysis revealed the maximum average flexibility (0.514) at residue 34 and the minimum (0.372) at residue 238 (Figure 5b). Domain prediction via SMART identified seven WD40 repeats (1–44, 59–98, 120–159, 163–202, 212–254, 257–313, 319–356), four low-complexity regions (LCDs) (416–427, 496–516, 544–560, 578–593), a PFAM: HIRA_B domain (453–475), and a PFAM: HIRA domain (767–965) (Figure 5c). The variants c.1273G>A, c.1521C>G, and c.1735A>G are located within the LCD of HIRA. c.2499G>A and c.2682C>T are found within the PFAM: HIRA domain. Therefore, these mutations should be considered more significant than the other variants identified in this study.
3.5.2. Physicochemical Properties of Ovine HIRA
ProtParam analysis predicted a molecular weight of 111,086.10 Da and an isoelectric point of 8.18. Leucine constituted the most abundant residue (10.5%), while tyrosine was the least frequent (1.4%). TMHMM analysis confirmed the absence of transmembrane domains (Figure 6a). Post-translational modification sites included 1 N-glycosylation site (Asn244) and 114 phosphorylation sites (32 threonine, 3 tyrosine, and 79 serine) (Figure 6b,c).
3.5.3. Phylogenetic Relationships
A phylogenetic tree was created with MEGA X using the Neighbor-Joining method (utilizing 1000 bootstrap replicates) based on HIRA sequences from model organisms (Figure 6d). The phylogenetic tree demonstrated that sequences from domesticated mammalian species (sheep and cattle) clustered separately from avian species and were also distinct from those of common rodent model organisms (rats and mice).
3.5.4. RNA Secondary Structure Modulation via Variants
RNAfold-based minimum free energy (MFE) predictions demonstrated secondary structural alterations in the point mutations [20]. Figure 7 specifically illustrates the protein secondary structural alterations at three litter size-associated variants. The mutations in exon 14 exhibited a decreasing trend in the MFE changes. Specifically, the MFE of c.1521C>T decreased from −72.50 to −71.90 kcal/mol, and that of c.1572C>T decreased from −72.50 to −70.90 kcal/mol. Meanwhile, the MFE of c. 1578G>A did not change. The structural alterations and MFE changes associated with other variants are illustrated in Figure S1.
3.5.5. Structural Consequences of Missense Mutations
SOPMA analysis predicted secondary structure proportions for wild-type HIRA as follows: 15.00% α-helices, 0.00% β-sheets, 65.20% random coils, and 19.80% extended strands (Figure 8a). The c.1273G>A (Ala425Thr) mutation induced 16 conformational alterations (indicated by black arrows), modifying proportions to 15.29% α-helices, 0.00% β-sheets, 64.22% random coils, and 20.49% extended strands (Figure 8b). Conversely, c.1735A>G (Thr579Ala) showed no secondary structure changes.
Homology modeling using chain A of growth and differentiation factor 8 (PDB ID: AOA667GSB8.1.A; 91.81% sequence identity) demonstrated that both Ala425Thr and Thr579Ala substitutions induced discernible tertiary structural perturbations (Figure 8c).
4. Discussion
A growing body of evidence indicates a critical link between the HIRA gene and reproduction. Studies conducted by Dilg et al. [8] and Pchelintsev et al. [21] have demonstrated that mutations in this gene significantly increase embryonic mortality rates. In Drosophila models, it has been reported that the overexpression of the dHira protein disrupts the regulation of cell cycle-related genes, thus leading to asynchronous nuclear division and subsequent developmental arrest during early embryogenesis [22]. Interestingly, research has also indicated that while female mice carrying HIRA mutations exhibit normal ovulation, they fail to produce offspring when paired with wild-type males [23]. Moreover, Zhou et al. explored genetic associations between specific HIRA variants, g.71874104G>A and g.71833755T>C, and litter size in Small-tail Han sheep [14]. In particular, our study is the first study of HIRA gene mutations linked to prolificacy in Sonid sheep.
Even though synonymous mutations do not alter the amino acid sequence, they can nonetheless influence the expression, splicing, stability [24,25,26], and secondary structure of mRNA [27,28], in addition to affecting the translation, folding [29], and functional properties of proteins [30]. Synonymous mutations can influence the production of active, properly folded proteins, thereby affecting physiological activity [29]. Research shows that a decrease in MFE may lead to the enhanced stability of the secondary structure of mRNA [30,31]. In our studies, the MFE of the silent LD-Sonid mutation induced a decreasing trend. Rose J et al. reported that even a single base pair alteration can result in changes to the secondary structures of mRNA. [31]. Based on predicted mRNA secondary structural alterations induced by mutations, we postulate that c.1572C>T serves as the primary functional variant due to its significant change in MFE (ΔMFE = −1.6 kcal/mol). Studies have shown that during the maturation of mammalian oocytes, high-stability mRNAs are preferentially retained in post-transcriptional regulation, whereas the accelerated degradation of mRNA during oocyte aging can impair embryonic developmental potential [32]. Increased mRNA stability may prevent the abnormal accumulation of reproduction-related genes, such as maternal mRNAs, thereby supporting normal embryonic development [33]. Enhanced mRNA stability may also promote the spatiotemporal regulation of reproductive genes in germ cells, thus affecting ovarian maturation and oocyte function [34,35]. Furthermore, high-stability mRNAs maintain the stable expression of critical genes under environmental stress, aiding reproductive cells in executing their functions, particularly in response to oxidative stress [36,37]. Recent studies have found that silent SNPs and mRNA secondary structures affect litter size in Awassi and Hamdani sheep [38]. Thus, we propose that c.1572C>T and c.1521C>G could affect the litter size of sheep by enhancing mRNA stability and altering its secondary structure. Although the c.1578G>A mutation did not alter the MFE in our prediction, even single-base changes can induce local structural rearrangements in mRNA that are not fully captured by global MFE calculations [30]. These subtle changes might affect the binding sites for regulatory microRNAs or RNA-binding proteins, thereby influencing HIRA mRNA stability or translation in a tissue-specific manner [39]. Therefore, the combined effect of all three mutations, through a potential interplay of mRNA stability and translation efficiency, might underlie the significant association with litter size. However, further experimental validation is needed to support this hypothesis.
In this study, two missense mutations c.1273G>A (A425T) and c.1735A>G (T425A) were identified. The mutation c.1273G>A exhibited significant departures from HWE, and fewer than ten individuals had the mutation c.1735A>G, which prevented effective association analysis. However, we still conducted analyses of the tertiary structures of the protein to provide references and scientific clues for the study of the HIRA gene. The two missense mutations were modeled within the tertiary structure of the HIRA protein, where they were predicted to cause noticeable structural changes. Notably, both missense mutations are localized within the LCD of the HIRA protein, as predicted by SMART analysis. Primarily, these mutations may disrupt the conservation of the amino acid sequence of the LCD, thereby leading to protein conformational disorders through the reduction in protein stability [40]. As a chaperone protein for histone H3.3, a decrease in the stability of HIRA will impair its ability to mediate histone deposition, thereby affecting chromatin remodeling [41]. Crucially, HIRA also regulates the expression of steroidogenic genes, which are controlled by gonadotropins such as Follicle-stimulating hormone receptor (FSHR) and luteinizing hormone/chorionic gonadotropin receptor (LHCGR), ad these also directly modulate estradiol (E_2_) biosynthesis [42,43]. Missense mutations may reduce the chromatin accessibility of these genes and inhibit hormonal synthesis, thereby disrupting the cooperative interaction between FSH/LH and granulosa cells as well as theca cells [44]. Consequently, we postulate that c.1273G>A (Ala425Thr) and c.1735A>G (Thr579Ala) maybe alter LCD architecture to impair granulosa cells functionality, ultimately perturbing oocyte maturation until ovulation in sheep. However, this hypothesis still needs additional experimentation. Future studies should verify the association between mutations and litter size in a large sheep population.
Sonid sheep is a characteristic breed of Mongolia sheep. Recent studies on Mongolia sheep have identified prolificacy-associated mutations in the following key genes: the g.46544883A>G, c.1040T>C, and g.46547859C>T of GDF9 in Mongolia sheep [18], the c.240C>T and c.279C>T of LEPR in both Mongolia and Ujimqin sheep [45], the g.29346567C>T and c.1470G > T of BMPR1B in Mongolia sheep [46], and the g.50985975 G>A and c.755 T>C of BMP15 in Mongolia sheep, along with the g.50988478C>A and g.50987863G>A of BMP15 in Ujimqin sheep [47]. The identification of breed-specific molecular markers for prolificacy is a key objective in genetic selection programs aimed at improving sheep productivity [48]. Based on the bioinformatic results and association analysis of this study, we identified that the mutations c.1521C>G, c.1572C>T, and c.1578G>A of LD are associated with litter size in Sonid sheep. Hence, the current findings may be utilized in marker-assisted selection (MAS) strategies to enhance prolificacy in low-fertility breeds including Sonid sheep.
5. Conclusions
In summary, the results of this study indicated that the LD-Sonid mutation (including c.1521C>G, c.1572C>T, and c.1578G>A) of HIRA was associated with litter size in Sonid sheep. These markers could be potentially utilized in MAS to enhance litter size in Sonid sheep. The silent mutation LD-Sonid could potentially improve the stability of mRNA by decreasing the MFE and affecting the secondary structure of HIRA mRNA. The missense mutations c.1273G>A (A425T) and c.1735A>G (T425A) induce conformational rearrangements in the tertiary protein structure, potentially altering the chromatin functions of HIRA. The effects of other mutation variants on the litter size of Sonid sheep require further investigation in a larger population of sheep. We should note that the association results require validation in larger, independent cohorts. Furthermore, the functional impact of the identified mutations on mRNA stability and the protein function of HIRA remains to be experimentally validated. In conclusion, these results offer significant genetic markers for sheep breeding and highlight new variants for further functional research on the HIRA gene.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1China National Commission of Animal Genetic Resources (CNCAGR) Sheep and Goats, Animal Genetic Resources in China China Agriculture Press Beijing, China 2011
- 2Xiang J. Li H. Guo Z. Li T. Yamada T. Li X. Bao S. Da L. Borjigin G. Cang M. Effect of FABP 4 Gene Polymorphisms on Fatty Acid Composition, Chemical Composition, and Carcass Traits in Sonid Sheep Animals 20251522610.3390/ani 1502022639858226 PMC 11758647 · doi ↗ · pubmed ↗
- 3Paz E. Quiñones J. Bravo S. Montaldo H.H. Sepúlveda N. Genotyping of BMPR 1B, BMP 15 and GDF 9 genes in Chilean sheep breeds and association with prolificacy Anim. Genet.201546989910.1111/age.1225425530177 · doi ↗ · pubmed ↗
- 4Ahlawat S. Sharma R. Roy M. Mandakmale S. Prakash V. Tantia M.S. Genotyping of Novel SN Ps in BMPR 1B, BMP 15, and GDF 9 Genes for Association with Prolificacy in Seven Indian Goat Breeds Anim. Biotechnol.20162719920710.1080/10495398.2016.116770627135147 · doi ↗ · pubmed ↗
- 5Liu Q.Y. Pan Z.Y. Wang X.Y. Hu W.P. Di R. Yao Y.X. Progress on major genes for high fecundity in ewes Front. Agric. Sci. Eng.2014128210.15302/J-FASE-2014042 · doi ↗
- 6Rai T.S. Puri A. Mc Bryan T. Hoffman J. Tang Y. Pchelintsev N.A. van Tuyn J. Marmorstein R. Schultz D.C. Adams P.D. Human CABIN 1 is a functional member of the human HIRA/UBN 1/ASF 1a histone H 3.3 chaperone complex Mol. Cell Biol.2011194107411810.1128/MCB.05546-11PMC 318736821807893 · doi ↗ · pubmed ↗
- 7Yang J.H. Song T.Y. Jo C. Park J. Lee H.Y. Song I. Differential regulation of the histone chaperone HIRA during muscle cell differentiation by a phosphorylation switch Exp. Mol. Med.201648 e 25210.1038/emm.2016.6827515126 PMC 5007640 · doi ↗ · pubmed ↗
- 8Dilg D. Saleh R.N. Phelps S.E. Rose Y. Dupays L. Murphy C. HIRA Is Required for Heart Development and Directly Regulates Tnni 2 and Tnnt 3P Lo S ONE 201611 e 016109610.1371/journal.pone.016109627518902 PMC 4982693 · doi ↗ · pubmed ↗