DSPicable V: TuMV exploits a dual-specificity phosphatase to suppress MAPK signaling and host resistance
Rory Osborne

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
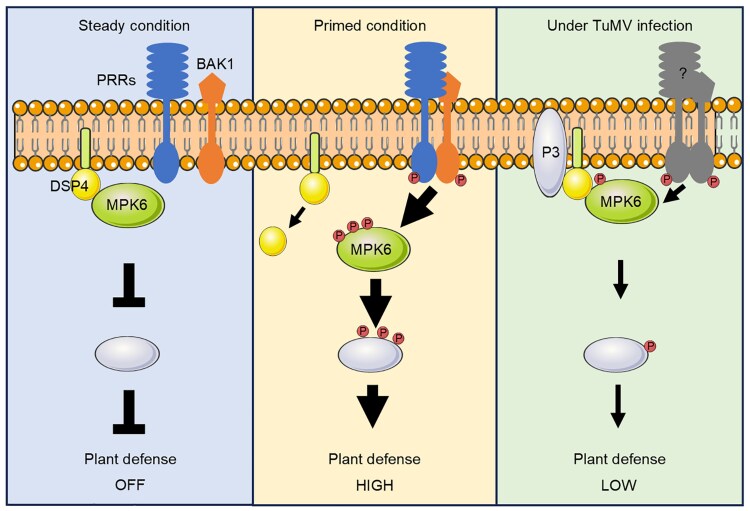 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProtein Tyrosine Phosphatases · Neutrophil, Myeloperoxidase and Oxidative Mechanisms
Posttranslational modifications modulate protein function in all living organisms. The addition or removal of chemical moieties at specific amino acid residues, such as phosphorylation, acetylation, and ubiquitylation, dynamically regulates protein activity and turnover. Phosphorylation is an essential component of cell signaling cascades via MITOGEN-ACTIVATED PROTEIN KINASEs (MPKs). In the context of plant immunity, MPKs directly phosphorylate transcription factors that activate defense responses after microbial perception at the cell surface (Sun and Zhang 2022). To avoid autoimmunity and associated stress, however, it is critical that these signals be appropriately downtuned when they are no longer required. While activation of MPK cascades is well characterized, mechanisms describing their attenuation are less well understood.
Previous studies have suggested that plant and fungal atypical dual-specificity phosphatases (DSPs) deactivate MAPK signaling in rice and Arabidopsis in response to diverse phytopathogens (He et al. 2012). Seeking to understand how the MPK cascade is dampened after viral infection, Yameng Luan, Xue Jiang, and colleagues (Luan et al. 2025) investigated the role of Arabidopsis DSPs in response to turnip mosaic virus (TuMV). The authors first characterized the physiologic resistance of 5 DSP1-5 Arabidopsis thaliana mutants to TuMV, revealing that the dsp4-2 mutation attenuated infection. Given that DSP4 was upregulated in this line, the authors hypothesized that DSP4 negatively regulates viral immunity.
Taking a combined approach of confocal microscopy and quantitative ultracentrifugation, the authors next showed that YFP-DSP4 predominantly localizes to the endomembrane and that this localization likely depends on the prenylation of a cysteine residue near its C-terminus. Furthermore, activation of pattern-triggered immunity (PTI) induced the dissociation of DSP4 from the endomembrane system, suggesting that it might have a role in downstream immunity signaling. Indeed, corroborating previous evidence that DSPs interact with MPKs (Xin et al. 2021), the authors used BiFC and CoIP to show that DSP4 physically interacted with multiple MPKs. Focusing on the DSP4–MPK6 interaction revealed that membrane-bound DSP4 could dephosphorylate MPK6 in vitro. This activity was lost when mutations to the DSP4 prenylation site were introduced, leading the authors to hypothesize that membrane association is required for effective MPK6 interaction/dephosphorylation.
MPK signaling was then shown to be an essential component of viral immunity. TuMV infection induced the expression of 16 MPK genes and promoted the phosphorylation of MPK3 and MPK6. Viral challenge of mpk6 knockouts revealed a lower rate of viral gene expression and infected leaf area as compared with wild type, which was further enhanced when these plants were complemented with pMPK6::FLAG-4×Myc-MPK6. However, concomitant repression of MPK3 via viral-induced gene silencing in a mpk6 knockout vastly decreased host resistance. In summary, the authors concluded that phosphorylated MPK3/6 redundantly suppressed TuMV infection.
Of course, no defense is foolproof, and pathogens regularly co-opt host machinery to facilitate their infection. Encoded in the TuMV genome is protein 3 (P3), a known membrane virulence factor that disrupts Rubisco activity (Lin et al. 2011). P3 was shown to interact with DSP4, which altered DSP4 localization and induced the formation of membrane condensates, suggesting that P3 might reduce the dephosphorylation activity of DSP4. However, an in vitro phosphorylation assay revealed that the opposite was true and that P3 enhanced DSP4 dephosphorylation of MPK6.
This conflicting result suggests either that the relocalization of DSP4 by P3 is an artifact induced by agroinfiltration (which itself induces PTI) or that these membrane condensates remain part of the endomembrane system and concentrate functional DSP4. Some evidence to the latter was demonstrated, as coexpression of both proteins increased the amount of the DSP4 associated with the endomembrane. While clearly showing that DSP4 attenuates an overactive immune system by dampening MPK6 activity, the mechanism of P3-mediated disruption to this process remains unclear. The authors proposed model (Fig.) suggests that P3 reduces the dissociation of DSP4 from the endomembrane system, increasing the dephosphorylation of MPK6 to impair host resistance. Further characterization of the MPK6–DSP4–P3 relationship, especially in vivo, will be required to decode this mechanism.
Recent related articles in The Plant Cell
Ding et al. (2022) show that the barley MPK3 directly phosphorylates the nucleoprotein of Barley yellow striate mosaic virus to inhibit infection, and they propose a cross-kingdom mechanism that regulates the plant–virus–insect interaction. Ma et al. (2025) characterize the activity of OXI1 kinase, which activates MAPKKK5 signaling in response to chitin to activate antifungal defenses. Kim et al. (2023) describe the regulation of the MAPKKK5 INTEGRIN-LINKED KINASE 5 (ILK5) by the extracellular ATP receptor P2K1 in response to elicitors of PTI and during infection to Pseudomonas syringae.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ding ZH, Gao Q, Tong X, Xu WY, Ma L, Zhang ZJ, Wang Y, Wang XB. MAP Ks trigger antiviral immunity by directly phosphorylating a rhabdovirus nucleoprotein in plants and insect vectors. Plant Cell. 2022:34(8):3110–3127. 10.1093/plcell/koac 14335567529 PMC 9338794 · doi ↗ · pubmed ↗
- 2He H, Su J, Shu S, Zhang Y, Ao Y, Liu B, Feng D, Wang J, Wang H. Two homologous putative protein tyrosine phosphatases, Os PFA-DSP 2 and At PFA-DSP 4, negatively regulate the pathogen response in transgenic plants. P Lo S One. 2012:7(4):e 34995. 10.1371/journal.pone.003499522514699 PMC 3325911 · doi ↗ · pubmed ↗
- 3Kim D, Chen D, Ahsan N, Jorge GL, Thelen JJ, Stacey G. The Raf-like MAPKKK INTEGRIN-LINKED KINASE 5 regulates purinergic receptor-mediated innate immunity in Arabidopsis. Plant Cell. 2023:35(5):1572–1592. 10.1093/plcell/koad 02936762404 PMC 10118279 · doi ↗ · pubmed ↗
- 4Lin L, Luo Z, Yan F, Lu Y, Zheng H, Chen J. Interaction between potyvirus P 3 and ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubis CO) of host plants. Virus Genes. 2011:43(1):90–92. 10.1007/s 11262-011-0596-621400205 · doi ↗ · pubmed ↗
- 5Luan Y, Jiang X, Wang Y, Chai M, Li F, Wang A, Wu X, Cheng X. A plant RNA virus hijacks a membrane-anchored dual-specificity phosphatase to attenuate MAPK-mediated immunity for robust infection. Plant Cell 2025:37(10):koaf 232. 10.1093/plcell/koaf 23240990618 · doi ↗ · pubmed ↗
- 6Ma M, Wang P, Chen R, Bai M, He Z, Xiao D, Xu G, Wu H, Zhou JM, Dou D, et al The OXIDATIVE SIGNAL-INDUCIBLE 1 kinase regulates plant immunity by linking microbial pattern–induced reactive oxygen species burst to MAP kinase activation. Plant Cell. 2025:37(1):koae 311. 10.1093/plcell/koae 311PMC 1166359939566103 · doi ↗ · pubmed ↗
- 7Sun T, Zhang Y. MAP kinase cascades in plant development and immune signaling. EMBO Rep. 2022:23(2):e 53817. 10.15252/embr.20215381735041234 PMC 8811656 · doi ↗ · pubmed ↗
- 8Xin J, Li C, Ning K, Qin Y, Shang JX, Sun Y. At PFA-DSP 3, an atypical dual-specificity protein tyrosine phosphatase, affects salt stress response by modulating MPK 3 and MPK 6 activity. Plant Cell Environ. 2021:44(5):1534–1548. 10.1111/pce.1400233464564 · doi ↗ · pubmed ↗