Metabolic effects of trehalose in mice of the C57BL/6 strain with obesity induced by a high carbohydrate-fat diet
A.B. Pupyshev, N.M. Bazhan, A.Yu. Kazantseva, T.V. Yakovleva, V.M. Belichenko, N.V. Goncharova, T.A. Korolenko, M.A. Tikhonova

TL;DR
Trehalose had minimal effects on metabolic parameters in obese C57BL/6 mice fed a high carbohydrate-fat diet, suggesting limited utility for treating diet-induced obesity in humans.
Contribution
The study evaluates trehalose's efficacy in a cafeteria diet-induced obesity model, relevant to human dietary obesity.
Findings
Trehalose increased caloric intake in obese mice but did not reduce body weight or improve metabolic parameters.
No significant changes in liver gene expression related to glucose transport, insulin sensitivity, or autophagy were observed.
The cafeteria diet model showed signs of metabolic syndrome, but trehalose failed to counteract these effects.
Abstract
The ability of trehalose to improve metabolic parameters in mice with experimental obesity has been shown to depend on the type of obesity model. In db/db mice, it reduced body weight, insulin, blood glucose, and cholesterol levels. In mice with obesity induced by high-fat dietary intake, it had no effect on body weight but reduced blood insulin levels with compensatory upregulation of insulin signaling gene expression. We studied the effect of trehalose on overweight and metabolic parameters in C57BL/6 inbred mice with obesity induced by a high carbohydrate-fat diet, the “cafeteria diet”. The cafeteria diet consisted of free access to water, standard chow, fatty foods (lard), and carbohydrates (biscuits) for 18 weeks. All mice were then randomly divided into four groups for four weeks of treatment: (1) water drinking, (2) drinking 3 % trehalose, (3) cafeteria diet and drinking water,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
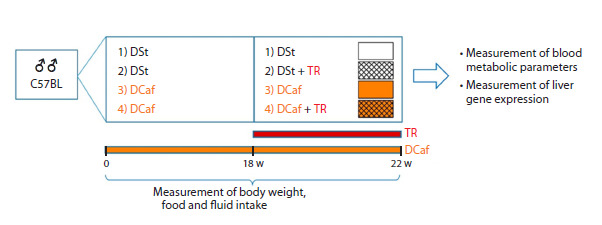 Fig. 1
Fig. 1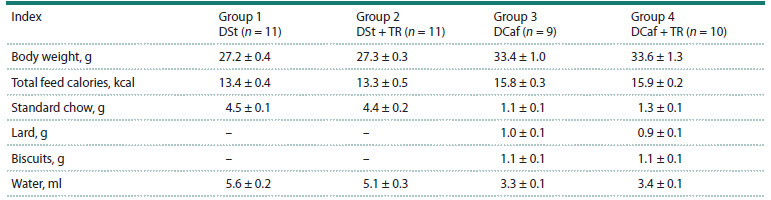 Table 1
Table 1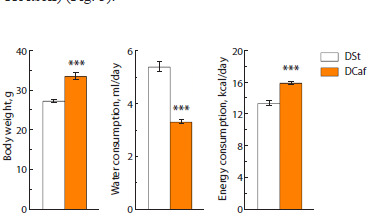 Fig. 2
Fig. 2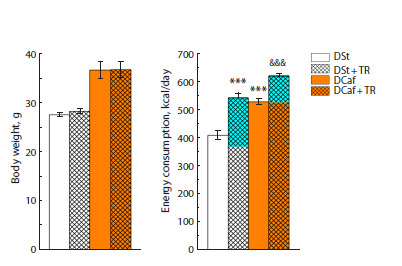 Fig. 3
Fig. 3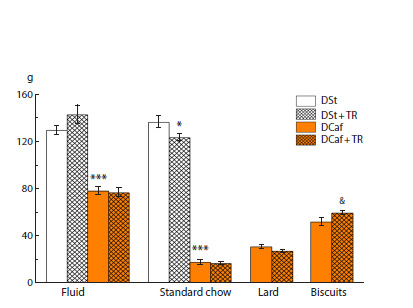 Fig. 4
Fig. 4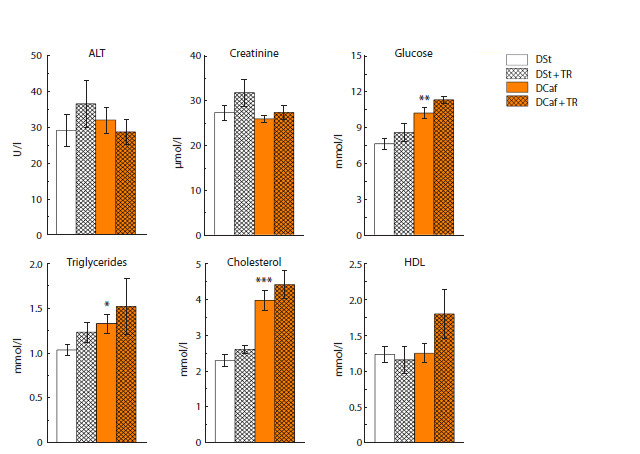 Fig. 5
Fig. 5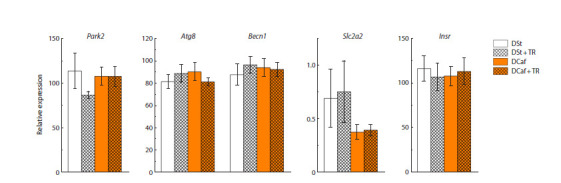 Fig. 6
Fig. 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDiet and metabolism studies · Adipose Tissue and Metabolism · Diet, Metabolism, and Disease
Introduction
Trehalose (TR) has multiple therapeutic properties, the main of which seem to be the chaperone-like activity and activation of autophagy, which is especially important for neuroprotection (Hosseinpour-Moghaddam et al., 2018; Pupyshev et al., 2022b). Other beneficial TR properties include positive effects on cellular metabolism, carbohydrate and lipid metabolism (Arai et al., 2019; Yaribeygi et al., 2019; Kobayashi et al., 2021), while diabetes, obesity, and neurodegeneration are known to be closely related (Pugazhenthi et al., 2017). Neurodegeneration is considered to be inhibited by TR through the activation of mTOR-independent autophagy (Sarkar, 2013; Tamargo-Gómez, Mariño, 2018).
At the same time, TR significantly affects carbohydrate metabolism, as it can reduce blood glucose levels and insulin resistance (Zhang et al., 2018; Zhang, DeBosch, 2019; Korolenko et al., 2021). TR also has a positive effect on lipid metabolism: it reduces the level of triglycerides in the liver and blood (Stachowicz et al., 2019; Zhang, DeBosch, 2019; Korolenko et al., 2021) and ultimately prevents the development of steatosis dependent on autophagy activity (Zhang et al., 2018; Ren et al., 2019; Su et al., 2025). TR regulates lipid metabolism partly through its effect on the release of adiponectin, which promotes fat burning (Arai et al., 2013; Mizote et al., 2016) through the reduction of the secretion of gastric inhibitory polypeptide GIP, which promotes obesity (Yoshizane et al., 2017) through the effects on the expression of lipoxygenase ALOXE3 and arginase 2, which increase energy consumption (Higgins et al., 2018; Zhang et al., 2019).
In a high-fat diet (HFD)-induced obesity model, TR reduces mesenteric and inguinal fat hypertrophy and brown fat gain (Arai et al., 2019), which is accompanied by an increase in thermogenesis both in C57BL/6 mice and in a genetic model of diabetic obesity, ob/ob mice (Zhang et al., 2018), while in the latter case the result depends on the activity of AMPK, TFEB and UCP1, but not on autophagy (Zhang et al., 2018; Rusmini et al., 2019). In general, the results on the effects of TR on excess body weight are quite contradictory (Arai et al., 2010, 2019; Liu et al., 2013; Sahebkar et al., 2019; Korolenko et al., 2021). Some studies reported a tendency to decrease in the mass of total visceral fat (no more than 5 %) and no significant effect on the body weight in mice kept on HFD after 8-week consumption of 2 % TR (Arai et al., 2013; Liu et al., 2013). According to other data (Korolenko et al., 2021), in db/db mice (a monogenic model of diabetic obesity), threeweek treatment with 2 % TR caused a noticeable decrease in the body weight of mice (more than 10 %) and a general therapeutic effect, in particular, a decrease in the levels of cholesterol, triglycerides, and plasma glucose.
It remains unclear to what extent the fat-reducing effect of TR depends on the type of diet or on mutations that cause obesity. In the present study, we considered that the development of alimentary forms of obesity in mice can be induced both by the consumption of HFD food and by the diet with an increased content of both fats and carbohydrates (carbohydrate-fat diet, “cafeteria diet”, DCaf), which is more common in the human population
In this regard, the study aimed to assess the effect of alimentary consumption of TR on the main metabolic parameters (body weight; food, calorie and water consumption) in C57BL/6 mice with obesity induced by keeping the animals on a carbohydrate-fat diet (DCaf). The biochemical parameters and lipid spectrum of the blood plasma and autophagy, assessed by the expression of autophagy genes in the liver, were also studied.
Materials and methods
Modeling obesity. All animal manipulations performed during the study met the ethical standards approved by the legal acts of the Russian Federation, the principles of the Basel Declaration and the recommendations of the independent bioethics committee of the Institute of Cytology and Genetics SB RAS (protocol number No. 76 dated 04/07/2021). The experiments were performed in male C57BL/6 mice of the conventional vivarium of the Institute of Cytology and Genetics SB RAS (Novosibirsk, Russia).
Three-month-old animals were kept individually in a cage under a 12-hour light : 12-hour dark regimen, at a temperature of 22–24 °C, and with free access to water and pelleted chow (Assortiment-Agro, Novosibirsk, Russia). After two weeks, the animals were either left on standard diet (n = 22) or transferred to a high-fat, high-carbohydrate diet (DCaf) (n = 19) consisting of unsalted pork lard, biscuits, and standard pelleted chow. DCaf causes obesity in C57BL/6 mice within 18 weeks (Makarova et al., 2013); DCaf content is the closest to the current daily diet in humans and it also allows to assess the consumption of certain dietary components
18 weeks after the start of DCaf intake, the animals were divided into four groups (Fig. 1): 1) standard diet (the consumption of standard chow) and water (n = 11); 2) standard diet (the consumption of standard chow) and 3 % TR solution (the rationale behind the TR dosage adopted in the current study was based on our recent study (Pupyshev et al., 2024)) (n = 11); 3) DCaf and water (n = 9); 4) DCaf and 3 % TR solution (n = 10). Throughout the experiment, mice body weight was measured weekly while food intake was measured three times a week.
Scheme of the experiment.Abbreviations: DSt – standard laboratory diet, DСaf – cafeteria diet, TR – trehalose
Blood biochemistry. After four weeks of trehalose consumption, the animals were sacrificed, blood and liver samples were collected. Blood serum samples were collected as described previously (Goncharova et al., 2016). Metabolic parameters (ALT, creatinine, glucose, triglycerides, cholesterol, and high-density lipoproteins (HDL)) were determined in the blood serum using an AU 680 Biochemical Analyzer (Beckman Coulter, USA).
Analysis of gene expression. Real-time qPCR was used to evaluate the relative expression levels of liver genes involved in the regulation of carbohydrate metabolism (Insr, encoding the insulin receptor; Slc2a2, encoding the glucose transporter type 2 GLUT2) and autophagy activity (Atg8, encoding the autophagy protein LC3-II; Becn1, encoding the protein Beclin 1; Park2, encoding the protein Parkin), as well as the reference genes Hprt1, B2m, Ppia
Total RNA was isolated from liver samples using the ExtractRNA kit (Evrogen, Moscow, Russia) according to the manufacturer’s instructions. cDNA synthesis was performed using the MMLV RT kit (Evrogen) according to the manufacturer’s protocol (https://evrogen.ru/products/cdna/synthesis/mmlv). The resulting cDNA samples were analyzed by qPCR on a LightCycler-480 II thermal cycler (Roche, Switzerland) using the BioMaster HS-qPCR SYBR Blue (2×) reagent kit (Biolabmiks, Novosibirsk, Russia), with forward (F) and reverse (R) primers (150 nM each) for the studied genes. Primers used for the target genes: Atg8 [F: 5′-AAA GAG TGG AAG ATG TCC GGC-3′ and R: 5′-ACC AGG AAC TTG GTC TTG TCC-3′], Becn1 [F: 5′-GAA CTC ACA GCT CCA TTA CTT A-3′ and R: 5′-ATC TTC GAG AGA CAC CAT CC-3′], Insr [F: 5′-ATC CTC GAA GGT GAG AAG AC-3′ and R: 5′-TGA TAC CAG AGC ATA GGA GC-3′], Park2 [F: 5′-GGT CCA GTT AAA CCC ACC TAC-3′ and R: 5′-TTA AGA CAT CGT CCC AGC AAG-3′], Slc2a2 [F: 5′-GGCTAATTTCAGGACTGGTT-3′ and R: 5′-TTTCTTTGCCCTGACTTCCT-3′]. Primers used for the reference genes: B2m [F: 5′-GTC TTT CTA TAT CCT GGC TCA-3′ and R: 5′-ATG CTT GAT CAC ATG TCT CG- 3′], Hprt1 [F: 5′-TAC CTA ATC ATT ATG CCG AGG A-3′ and R: 5′-GGT CAG CAA AGA ACT TAT AGC C-3′], Ppia [F: 5′- AAA GTT CCA AAG ACA GCA GAA AA-3′ and R: 5′-GCC AGG ACC TGT ATG CTT TAG-3′]. The relative concentration of the tested cDNA was determined using Light- Cycler 480 software (1.5.1 version) and calibration curves.
Statistics. The data were analyzed using STATISTICA 10.0 (StatSoft, TIBCO Software Inc., Palo Alto, CA, USA). The results were expressed as the mean ± standard error of the mean. A two-tailed Student’s t-test was used to compare sample means. A statistically significant level of differences was accepted at p < 0.05 (two-tailed).
Results
Modeling diabetic obesity
Before the start of treatment with 3 % TR solution (after 18 weeks of maintenance on DSt or DCaf), mice from comparison groups 1 and 2, as well as obese animals from groups 3 and 4, did not differ in body weight, food consumption, or taste preferences (see the Table, Fig. 2).
Body weight, water consumption, total energy and dietary component intake in mice in four groupsNote. Mice in groups 1 and 2 maintained on standard diet (DSt) and in groups 3 and 4 maintained on the cafeteria diet (DCaf ) for 18 weeks before the start of treatment with 3 % trehalose (TR) solution in groups 2 and 4. Daily component consumption is indicated. Results are expressed as M ± m (n, number of animals).
Body weight, water consumption, and total energy intake of mice maintained on standard diet (n = 22) or cafeteria diet (n = 19) for 18 weeks before they were given 3 % trehalose solution.** p < 0.001 vs. DSt.*
Mice that consumed DCaf for 18 weeks developed excess body weight (obesity): body weight increased by 25 % (p < 0.001), energy consumption increased by 20 % (p < 0.001), and water consumption, on the contrary, was reduced by 40 % (p < 0.001) compared to mice in the control group (Fig. 2).
In animals kept on DCaf, the indicator of hyperglycemia (blood glucose level) increased by 35 % (see below), and this differs from the effect of DCaf in other studies, where blood glucose level increased more significantly (Parafati et al., 2015), or in genetic models of obesity, ob/ob and db/db mice (Pelletier et al., 2020; Korolenko et al., 2021). In our experiment, excess weight increase was not accompanied by the development of high hyperglycemia.
Effects of TR on metabolic parameters in mice kept on standard laboratory diet
Consumption of a 3 % TR solution for four weeks in control mice did not affect the body weight of the animals (Fig. 3). These mice ate less food (p < 0.05), while they showed a tendency for an increase in fluid consumption by 10 % (Fig. 4). Taking into account the caloric supplementation from TR in drinking, the total kcal consumption in control mice drinking a 3 % TR solution was more than 30 % higher than in control animals drinking water (p < 0.001) (Fig. 3).
Body weight and total energy intake in mice maintained on standard diet and cafeteria diet for four weeks during which the animals consumed water or 3 % trehalose solution.The blue color shows the number of kcal obtained with drinking 3 % TR solution. *** p < 0.001 vs. DSt; &&& p < 0.001 vs. DCaf and water group (t-test).
Total fluid and various food components intake of mice maintained on standard diet or cafeteria diet for four weeks during which the animals consumed water or 3 % trehalose solution. p < 0.05, *** p < 0.001 vs. DSt; & p < 0.05 vs. DCaf and water group (t-test).*
Effects of TR on metabolic parameters in mice kept on cafeteria diet
Despite our expectations, the body weight of mice kept on DCaf did not change under the influence of TR (Fig. 3). TR also had a weak effect on their consumption of standard food, liquid, or lard but increased the consumption of the carbohydrate component, biscuits (Fig. 4). TR significantly increased the number of calories consumed, by 18 % (taking into account the unchanged body weight in these animals compared to the group drinking water and the caloric supplementation from TR itself) (Fig. 3).
TR did not produce significant changes in metabolic blood indices in mice maintained on DCaf or in the control group (except for a noticeable trend (p < 0.07) in blood glucose growth) (Fig. 5). The maintenance of mice on DCaf per se influenced the overall metabolism to a certain extent increasing the levels of glucose (p < 0.01), triglycerides (p < 0.05), and especially those of blood cholesterol (p < 0.001), i. e. changes in carbohydrate and lipid metabolism were registered.
Blood biochemical parameters in mice maintained on standard diet or cafeteria diet and given water or 3 % trehalose solution for four weeks. p <0.05, ** p < 0.01, *** p < 0.001 vs. DSt.*
Effects of TR on transcription of carbohydrate metabolism and autophagy genes in mice kept on cafeteria diet
Neither DCaf nor TR consumption affected the expression of genes regulating glucose uptake from blood (Slc2a2, Insr) or related to autophagy activity (Atg8, Becn1, Park2) in the liver of mice (Fig. 6).
The mRNA levels of autophagy (Park2, Atg8, Becn1) and carbohydrate metabolism (Slc2a2, Insr) genes in the liver of mice kept on standard diet or cafeteria diet and given water or 3 % trehalose solution for four weeks
Discussion
The maintenance of mice on DCaf for 18 weeks was accompanied by an increase in caloric intake (by 20 %) and, as a result, the development of excess body weight (by 25 %). That allowed us to study a normalizing effect of TR on body weight gain found in previous studies in db/db mice (Korolenko et al., 2021). TR as an energy substrate (Sato et al., 1999) increased caloric energy intake both in mice of the control group and in mice given DCaf. Notably, in both groups of mice, an increase in caloric intake mainly associated with TR intake did not augment body mass (Fig. 3). Hence, TR stimulates caloric energy expenditure without affecting body weight. To some extent, this is in line with previous findings that demonstrated the ability of long-term TR treatment to stimulate caloric expenditure in thermogenesis and brown fat burning processes in C57BL/6 mice kept on both standard and high-fat diets (Arai et al., 2013, 2019) and in obese ob/ob mice (Zhang et al., 2018).
Another possible way of regulation by alimentary TR involves the induction of cellular starvation accompanied by activation of cytoprotective autophagy (DeBosch et al., 2016; Mayer et al., 2016; Zhang, DeBosch, 2019). The mechanism is associated with inhibition of the GLUT8 glucose transmembrane transporter, thereby causing energy deficiency in liver cells leading to activation of the adenosine monophosphatedependent kinase AMPK and its pleiotropic metabolic effect including inhibition of biosynthesis, energy consumption, and activation of autophagy. In the present study, TR appeared to lose its ability to simulate the fasting effect (Zhang, DeBosch, 2019), which leads to autophagy activation. The attenuation of the regulatory properties of TR here is consistent with its lack of effect on the transcription of autophagy genes responding to TR by elevation in our other studies (Pupyshev et al., 2022a). The reason for the weakening of the regulatory properties of TR in C57BL/6 mice remains unclear. Such a switch of the effect of TR does not seem to depend on the diet, as the effects of TR were similar both in control mice and in DCaf-given ones. Perhaps, in the conditions in our study (3 % trehalose, 28 days), there is an escape of TR from the quantitative energy cleavage described earlier (Sato et al., 1999), and then the contradiction between the growth of calories consumed and the lack of weight gain is smoothed out.
The loss of regulatory properties of TR in mice in our experiment is not consistent with the effects of TR revealed in db/db mice with diabetic obesity (Korolenko et al., 2021). In db/db mice, TR reduced the body mass (by more than 10 %),reduced blood glucose levels, and produced a general recovery effect by lowering blood cholesterol and triglycerides. However, in studies in rabbits, as here, a weak effect of TR on the blood lipid spectrum was found (Sahebkar et al., 2019).
Given the similarity of the model of obesity in DCaf-given mice with the typical development of obesity in humans, the results question the use of TR to correct this most common type of obesity in humans. At the same time, our results do not exclude the approach of treating patients with severe obesity with TR because such treatment was successful for mice with almost 50 % excess weight (Korolenko et al., 2021).
Conclusion
Based on the well-known effect of TR on simulation of starvation (induction of autophagy) and reduction of excess body weight in db/db mice, a study of its effect in C57BL/6 mice given carbohydrate-fat diet (DCaf), a model an alimentary obesity in humans, was performed. In the control and obese (by 25 %) mice, TR (3 % solution drinking, 28 days) augmented significantly the number of calories consumed while this increase in energy consumption was not accompanied by an increase in body weight gain of the mice. The excess in calories consumed might be spent on enhancing the processes of thermogenesis and brown fat burning (Arai et al., 2013, 2019). Trehalose produced only a tendency of increase in blood metabolic parameters (glucose, cholesterol, triglycerides, HDL) and had no effect on the expression of genes regulating carbohydrate metabolism (Slc2a2, Insr) or autophagy genes (Atg8, Becn1, Park2). In the present study, in mice kept on DCaf, TR did not demonstrate an ability to reduce diabetes and obesity induced by DCaf, nor useful properties for the correction of common dietary type of obesity in humans.
Conflict of interest
The authors declare no conflict of interest.
References
Arai C., Arai N., Mizote A., Kohno K., Iwaki K., Hanaya T., Arai S., Ushio S., Fukuda S. Trehalose prevents adipocyte hypertrophy and mitigates insulin resistance. Nutr Res. 2010;30(12):840-848. doi 10.1016/j.nutres.2010.10.009
Arai C., Miyake M., Matsumoto Y., Mizote A., Yoshizane C., Hanaya Y., Koide K., Yamada M., Hanaya T., Arai S., Fukuda S. Trehalose prevents adipocyte hypertrophy and mitigates insulin resistance in mice with established obesity. J Nutr Sci Vitaminol (Tokyo). 2013;59(5):393-401. doi 10.3177/jnsv.59.393
Arai C., Arai N., Arai S., Yoshizane C., Miyata S., Mizote A., Suyama A., Endo S., Ariyasu T., Mitsuzumi H., Ushio S. Continuous intake of trehalose induces white adipose tissue browning and enhances energy metabolism. Nutr Metab. 2019;16:45. doi 10.1186/ s12986-019-0373-4
DeBosch B.J., Heitmeier M.R., Mayer A.L., Higgins C.B., Crowley J.R., Kraft T.E., Chi M., Newberry E.P., Chen Z., Finck B.N., Davidson N.O., Yarasheski K.E., Hruz P.W., Moley K.H. Trehalose inhibits solute carrier 2A (SLC2A) proteins to induce autophagy and prevent hepatic steatosis. Sci Signal. 2016;9(416):ra21. doi 10.1126/ scisignal.aac5472
Goncharova N.V., Pupyshev A.B., Filyushina E.E., Loktev K.V., Korolenko E.Ts., Lushnikova E.L., Molodykh O.P., Korolenko T.A., Churin B.V. Depression of macrophages modifies serum lipid profile in hyperlipidemia. Bull Exp Biol Med. 2016;160(5):617-621. doi 10.1007/s10517-016-3231-7
Higgins C.B., Zhang Y., Mayer A.L., Fujiwara H., Stothard A.I., Graham M.J., Swarts B.M., DeBosch B.J. Hepatocyte ALOXE3 is induced during adaptive fasting and enhances insulin sensitivity by activating hepatic PPARγ. JCI Insight. 2018;3(16):e120794. doi 10.1172/jci.insight.120794
Hosseinpour-Moghaddam K., Caraglia M., Sahebkar A. Autophagy induction by trehalose: molecular mechanisms and therapeutic impacts. J Cell Physiol. 2018;233(9):6524-6543. doi 10.1002/jcp.26583
Kobayashi M., Yasukawa H., Arikawa T., Deguchi Y., Mizushima N., Sakurai M., Onishi S., Tagawa R., Sudo Y., Okita N., Higashi K., Higami Y. Trehalose induces SQSTM1/p62 expression and enhances lysosomal activity and antioxidative capacity in adipocytes. FEBS Open Bio. 2021;11(1):185-194. doi 10.1002/2211-5463.13055
Korolenko T.A., Dubrovina N.I., Ovsyukova M.V., Bgatova N.P., Tenditnik M.V., Pupyshev A.B., Akopyan A.A., Goncharova N.V., Lin C.L., Zavjalov E.L., Tikhonova M.A., Amstislavskaya T.G. Treatment with autophagy inducer trehalose alleviates memory and behavioral impairments and neuroinflammatory brain processes in db/db mice. Cells. 2021;10(10):2557. doi 10.3390/cells10102557
Liu M., Zhang M., Ye H., Lin S., Yang Y., Wang L., Jones G., Trang H. Multiple toxicity studies of trehalose in mice by intragastric administration. Food Chem. 2013;136(2):485-490. doi 10.1016/j.foodchem. 2012.09.031
Makarova E.N., Chepeleva E.V., Panchenko P.E., Bazhan N.M. Influence of abnormally high leptin levels during pregnancy on metabolic phenotypes in progeny mice. Am J Physiol Regul Integr Comp Physiol. 2013;305(11):R1268-R1280. doi 10.1152/ajpregu.00162.2013
Mayer A.L., Higgins C.B., Heitmeier M.R., Kraft T.E., Qian X., Crowley J.R., Hyrc K.L., Beatty W.L., Yarasheski K.E., Hruz P.W., DeBosch B.J. SLC2A8 (GLUT8) is a mammalian trehalose transporter required for trehalose-induced autophagy. Sci Rep. 2016;6: 38586. doi 10.1038/srep38586
Mizote A., Yamada M., Yoshizane C., Arai N., Maruta K., Arai S., Endo S., Ogawa R., Mitsuzumi H., Ariyasu T., Fukuda S. Daily intake of trehalose is effective in the prevention of lifestyle-related diseases in individuals with risk factors for metabolic syndrome. J Nutr Sci Vitaminol (Tokyo). 2016;62(6):380-387. doi 10.3177/jnsv. 62.380
Parafati M., Lascala A., Morittu V.M., Trimboli F., Rizzuto A., Brunelli E., Coscarelli F., Costa N., Britti D., Ehrlich J., Isidoro C., Mollace V., Janda E. Bergamot polyphenol fraction prevents nonalcoholic fatty liver disease via stimulation of lipophagy in cafeteria diet-induced rat model of metabolic syndrome. J Nutr Biochem. 2015;26(9):938-948. doi 10.1016/j.jnutbio.2015.03.008
Pelletier R.M., Layeghkhavidaki H., Vitale M.L. Glucose, insulin, insulin receptor subunits α and β in normal and spontaneously diabetic and obese ob/ob and db/db infertile mouse testis and hypophysis. Reprod Biol Endocrinol. 2020;18(1):25. doi 10.1186/s12958-020- 00583-2
Pugazhenthi S., Qin L., Reddy P.H. Common neurodegenerative pathways in obesity, diabetes, and Alzheimer’s disease. Biochim Biophys Acta Mol Basis Dis. 2017;1863(5):1037-1045. doi 10.1016/j.bbadis. 2016.04.017
Pupyshev A.B., Belichenko V.M., Tenditnik M.V., Bashirzade A.A., Dubrovina N.I., Ovsyukova M.V., Akopyan A.A., Fedoseeva L.A., Korolenko T.A., Amstislavskaya T.G., Tikhonova M.A. Combined induction of mTOR-dependent and mTOR-independent pathways of autophagy activation as an experimental therapy for Alzheimer’s disease-like pathology in a mouse model. Pharmacol Biochem Behav. 2022a;217:173406. doi 10.1016/j.pbb.2022.173406
Pupyshev A.B., Klyushnik T.P., Akopyan A.A., Singh S.K., Tikhonova M.A. Disaccharide trehalose in experimental therapies for neurodegenerative disorders: molecular targets and translational potential. Pharmacol Res. 2022b;183:106373. doi 10.1016/j.phrs.2022. 106373
Pupyshev A.B., Akopyan A.A., Tenditnik M.V., Ovsyukova M.V., Dubrovina N.I., Belichenko V.M., Korolenko T.A., Zozulya S.A., Klyushnik T.P., Tikhonova M.A. Alimentary treatment with trehalose in a pharmacological model of Alzheimer’s disease in mice: effects of different dosages and treatment regimens. Pharmaceutics. 2024;16(6):813. doi 10.3390/pharmaceutics16060813
Ren H., Wang D., Zhang L., Kang X., Li Y., Zhou X., Yuan G. Catalpol induces autophagy and attenuates liver steatosis in ob/ob and highfat diet-induced obese mice. Aging (Albany NY ). 2019;11(21):9461- 9477. doi 10.18632/aging.102396
Rusmini P., Cortese K., Crippa V., Cristofani R., Cicardi M.E., Ferrari V., Vezzoli G., … Galbiati M., Garrè M., Morelli E., Vaccari T., Poletti A. Trehalose induces autophagy via lysosomal-mediated TFEB activation in models of motoneuron degeneration. Autophagy. 2019; 15(4):631-651. doi 10.1080/15548627.2018.1535292
Sahebkar A., Hatamipour M., Tabatabaei S.A. Trehalose administration attenuates atherosclerosis in rabbits fed a high-fat diet. J Cell Biochem. 2019;120(6):9455-9459. doi 10.1002/jcb.28221
Sarkar S. Regulation of autophagy by mTOR-dependent and mTORindependent pathways: autophagy dysfunction in neurodegenerative diseases and therapeutic application of autophagy enhancers. Biochem Soc Trans. 2013;41(5):1103-1130. doi 10.1042/ BST20130134
Sato S., Okamoto K., Minami R., Kohri H., Yamamoto S. Trehalose can be used as a parenteral saccharide source in rabbits. J Nutrition. 1999;129(1):158-164. doi 10.1093/jn/129.1.158
Stachowicz A., Wiśniewska A., Kuś K., Kiepura A., Gębska A., Gajda M., Białas M., Totoń-Żurańska J., Stachyra K., Suski M., Jawień J., Korbut R., Olszanecki R. The influence of trehalose on atherosclerosis and hepatic steatosis in apolipoprotein E knockout mice. Int J Mol Sci. 2019;20(7):1552. doi 10.3390/ijms20071552
Su S., Liu X., Zhu M., Liu W., Liu J., Yuan Y., Fu F., Rao Z., Liu J., Lu Y., Chen Y. Trehalose ameliorates nonalcoholic fatty liver disease by regulating IRE1α-TFEB signaling pathway. J Agric Food Chem. 2025;73(1):521-540. doi 10.1021/acs.jafc.4c08669
Tamargo-Gómez I., Mariño G. AMPK: regulation of metabolic dynamics in the context of autophagy. Int J Mol Sci. 2018;19(12):3812. doi 10.3390/ijms19123812
Yaribeygi H., Yaribeygi A., Sathyapalan T., Sahebkar A. Molecular mechanisms of trehalose in modulating glucose homeostasis in diabetes. Diabetes Metab Syndr. 2019;13(3):2214-2218. doi 10.1016/ j.dsx.2019.05.023
Yoshizane C., Mizote A., Yamada M., Arai N., Arai S., Maruta K., Mitsuzumi H., Ariyasu T., Ushio S., Fukuda S. Glycemic, insulinemic and incretin responses after oral trehalose ingestion in healthy subjects. Nutr J. 2017;16(1):9. doi 10.1186/s12937-017-0233-x
Zhang Y., DeBosch B.J. Using trehalose to prevent and treat metabolic function: effectiveness and mechanisms. Curr Opin Clin Nutr Metab Care. 2019;22(4):303-310. doi 10.1097/MCO.0000000000000568
Zhang Y., Sowers J.R., Ren J. Targeting autophagy in obesity: from pathophysiology to management. Nat Rev Endocrinol. 2018;14(6): 356-376. doi 10.1038/s41574-018-0009-1
Zhang Y., Higgins C.B., Fortune H.M., Chen P., Stothard A.I., Mayer A.L., Swarts B.M., DeBosch B.J. Hepatic arginase 2 (Arg2) is sufficient to convey the therapeutic metabolic effects of fasting. Nat Commun. 2019;10(1):1587. doi 10.1038/s41467- 019-09642-8