Association of the rs823144 variant of the RAB29 gene with the activity of lysosomal hydrolases in blood cells and risk of Parkinson’s disease
K.S. Basharova, A.I. Bezrukova, K.A. Senkevich, G.V. Baydakova, A.V. Rybakov, I.V. Miliukhina, A.A. Timofeeva, E.Yu. Zakharova, S.N. Pchelina, T.S. Usenko

TL;DR
A genetic variant in the RAB29 gene is linked to lower Parkinson’s disease risk and changes in lysosomal enzyme activity and sphingolipid levels in blood.
Contribution
First demonstration of RAB29 rs823144 C allele association with PD risk and altered lysosomal enzyme activity and sphingolipid profiles.
Findings
The RAB29 rs823144 C allele is associated with reduced PD risk in the Northwestern Russian population.
The C allele correlates with increased GLA activity and decreased LysoGb3 and LysoSM levels in PD patients.
No significant link was found between the C allele and RAB29 mRNA expression in blood cells.
Abstract
Recent genome-wide association studies have identified a link between the RAB29 gene and Parkinson’s disease (PD). The Rab29 protein encoded by RAB29 regulates leucine-rich repeat kinase 2 (LRRK2). Mutations in the LRRK2 gene increase its kinase activity and contribute to autosomal dominant forms of PD. Previous research has shown that altered LRRK2 kinase activity may correlate with the activity of lysosomal hydrolases and the concentration of sphingolipids. This study aimed to assess the association of the rs823144 variant in the promoter region of the RAB29 gene with PD risk, and to evaluate RAB29 expression, lysosomal hydrolase activity, and sphingolipid concentrations in the blood of PD patients. We screened the rs823144 variant of the RAB29 gene in a cohort of PD patients (N = 903) and controls (N = 618) using next-generation sequencing (NGS) and polymerase chain reaction (PCR)…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
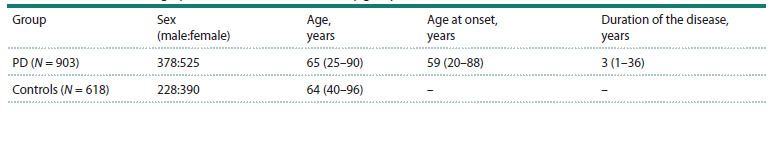 Table 1
Table 1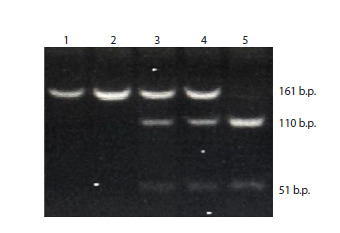 Fig. 1
Fig. 1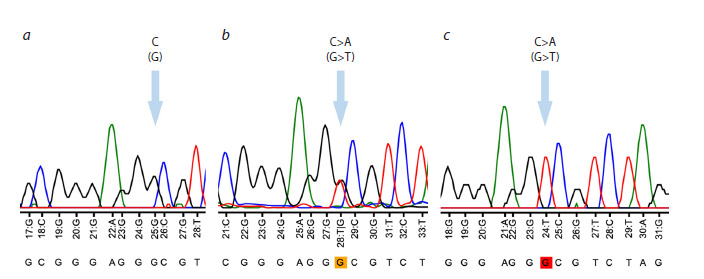 Fig. 2
Fig. 2 Table 2
Table 2 Table 3
Table 3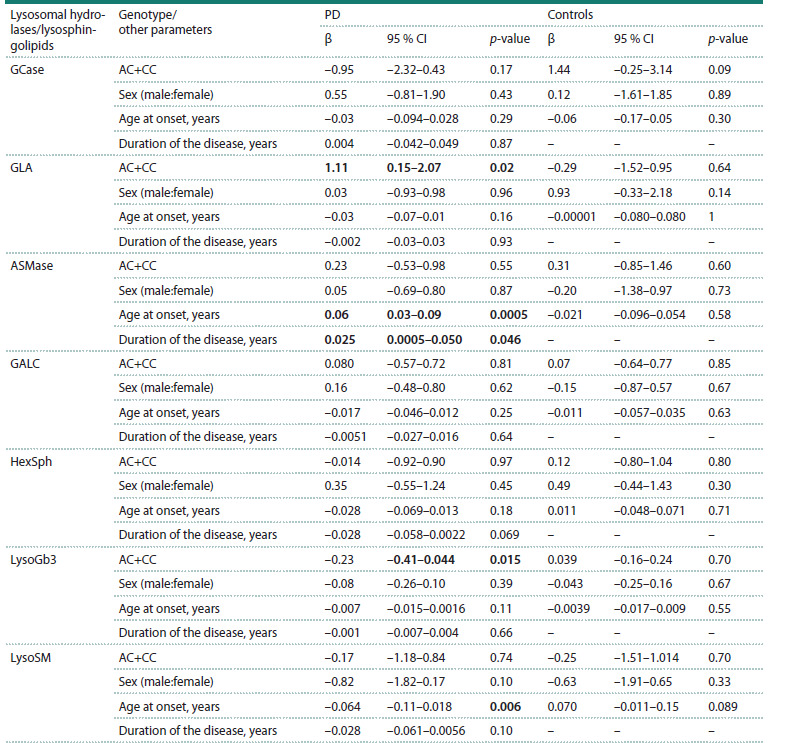 Table 4
Table 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLysosomal Storage Disorders Research · Parkinson's Disease Mechanisms and Treatments · Cellular transport and secretion
Introduction
Parkinson’s disease (PD) is a common, slowly progressive neurodegenerative disorder characterized by the loss of dopaminergic neurons in the substantia nigra (SN) of the brain (Lill, 2016). A central pathological mechanism in PD pathogenesis is the accumulation and aggregation of the alpha-synuclein protein in the SN. Although PD is primarily sporadic, approximately 15 % of patients report a positive family history. The molecular mechanisms underlying PD remain largely unclear; however, increasing evidence implicates lysosomal dysfunction as a key contributor to disease pathogenesis (Nechushtai et al., 2023). In particular, our group and others have demonstrated reduced lysosomal hydrolase activity and altered sphingolipid levels in the peripheral fluids of patients with idiopathic PD (Alcalay et al., 2015; Galper et al., 2022; Usenko et al., 2022). Additionally, changes in lysosomal enzyme activity and sphingolipid concentrations have been observed in PD cases associated with mutations in the LRRK2 gene, one of the most common monogenic forms of the disease (Alcalay et al., 2015; Usenko et al., 2023, 2024).
The LRRK2 gene encodes leucine-rich repeat kinase 2 (LRRK2), a multidomain protein implicated in Parkinson’s disease pathogenesis (Zimprich et al., 2004). A key group of LRRK2 substrates comprises small Rab GTPases, which are critical regulators of vesicular trafficking, particularly within the endolysosomal system (Steger et al., 2016; Wang et al., 2014). Dysregulated LRRK2 kinase activity disrupts the trafficking of lysosomal hydrolases to their proper destinations, thereby impairing lysosomal function (MacLeod et al., 2013; Ysselstein et al., 2019; Rivero-Ríos et al., 2020; Kedariti et al., 2022).
Among the LRRK2 substrates, Rab29 – encoded by the RAB29 gene – has attracted particular interest (Steger et al., 2016). Rab29 has been identified as a key upstream regulator of LRRK2, responsible for its activation (Liu et al., 2018; Madero-Pérez et al., 2018; Purlyte et al., 2018; Kuwahara Iwatsubo, 2020). Rab29 localizes to the membranes of lysosomes and the Golgi apparatus, where it recruits inactive cytoplasmic LRRK2 monomers and promotes their oligomerization into active dimers or tetramers (Purlyte et al., 2018; Zhu et al., 2023). The RAB29 gene is located within the PARK16 locus, which has previously been associated with reduced PD risk (Satake et al., 2009; Pihlstrøm et al., 2015; Nalls et al., 2019). A recent multi-trait analysis of genomewide association studies (MTAG) further confirmed the association between RAB29 and PD at both the transcriptomic and proteomic levels (Shi et al., 2024).
Several studies have identified variants in the promoter region of RAB29 that are associated with a reduced risk of Parkinson’s disease. These variants are thought to influence RAB29 gene expression levels (Gan-Or et al., 2012; Khaligh et al., 2017; Sun et al., 2021), potentially modulating the activation of LRRK2 and thereby affecting lysosomal hydrolase activity in PD
The aim of this study was to investigate the association of the rs823144 single nucleotide polymorphism (SNP), located in the promoter region of RAB29, with PD risk, RAB29 gene expression, lysosomal hydrolase activity – including glucocerebrosidase (GCase), α-galactosidase (GLA), galactocerebrosidase (GALC), and acid sphingomyelinase (ASMase) – and the concentrations of lysosphingolipids in the blood. The lysosphingolipids analyzed included hexosylsphingosine (HexSph), a mixture of glucosylsphingosine (GlcSph) and galactosylsphingosine (GalSph); lysosphingomyelin (LysoSM); and lysoglobotriaosylsphingosine (LysoGb3). These parameters were evaluated in both PD patients and healthy control subjects
Materials and methods
Characteristics of the study groups. The study included 903 patients with sporadic PD and 618 control individuals matched for age and gender. All patients were recruited from the clinic of the N.P. Bechtereva Institute of the Human Brain of the Russian Academy of Sciences. The control group comprised individuals seen at the consultative and diagnostic center of the First St. Petersburg State Medical University named after academician I.P. Pavlov. To exclude PD and other neurodegenerative disorders, all control participants underwent neurological examination. Clinical and demographic characteristics of the study groups are presented in Table 1. No significant differences in age or gender distribution were observed between the groups (p > 0.05).
Clinical and demographic characteristics of the study groupsNote. PD – Parkinson’s disease; data are presented as median (min–max).
All procedures involving human participants were conducted in accordance with the ethical standards of the National Research Ethics Committee and the 1964 Declaration of Helsinki and its later amendments or comparable ethical standards. Written informed consent was obtained from each participant prior to inclusion in the study. The study protocol was approved by the Ethics Committee of the First St. Petersburg State Medical University named after academician I.P. Pavlov (Protocol No. 275, dated September 04, 2023).
Genetic analysis. Two methods were used to genotype the rs823144 variant in the RAB29 gene: massively parallel sequencing (next-generation sequencing, NGS) and polymerase chain reaction (PCR) followed by restriction fragment length polymorphism (RFLP) analysis. Peripheral blood samples were collected from all study participants, and genomic DNA was extracted using phenol-chloroform extraction, as previously described (Maniatis et al., 1994).
NGS genotyping of the rs823144 variant in the RAB29 gene was performed in a subset of 521 PD patients and 420 control individuals using molecular inversion probes, as previously described (Rudakou et al., 2021). Sequencing was conducted on the Illumina NovaSeq 6000 SP PE100 platform. Sequence alignment was performed using the Burrows–Wheeler Aligner (BWA) with the hg19 human genome reference (Li, Durbin, 2009). Variant calling and post-alignment quality control were carried out using the Genome Analysis Toolkit (GATK, v3.8) (McKenna et al., 2010). Variants were filtered based on coverage depth and quality metrics; only those with a minimum read depth >30 and a quality score >20 were included in the analysis.
An additional 473 PD patients and 384 controls were screened for rs823144 using PCR followed by restriction analysis. Primer sequences were designed using Primer3 v. 0.4.0 (http://bioinfo.ut.ee/primer3-0.4.0/) (FOR: 5ʹ-CCCTGCA CGTGACGCTTG-3ʹ, REV: 5ʹ-GAATCCCAGTCAGCTC CTTACA-3ʹ). Restriction enzyme selection was performed using the NEBCutter tool (Vincze et al., 2003), and BstAC I was chosen for RFLP analysis (Fig. 1).
Electropherogram showing the results of genotyping of the rs823144 variant in the RAB29 gene1, 2 – homozygote for the A allele (genotype AA), 3, 4 – heterozygote (genotype AC), 5 – homozygote for the C allele (genotype CC).
A total of 91 PD patients and 186 control individuals were genotyped for the rs823144 variant in the RAB29 gene using both PCR with restriction analysis and NGS. These overlaps were accounted for in subsequent statistical analyses
To validate the results obtained from both methods, a subset of samples was confirmed by Sanger sequencing using a Nanofor-05 genetic analyzer (Synthol, Russia). The Sanger sequencing data were visualized and analyzed using Tracy software (Rausch et al., 2020) (Fig. 2).
Graphical representation of Sanger sequencing results (electropherogram displaying the results of genotyping of the rs823144 variant of the RAB29 gene).a – homozygote for the C allele (genotype CC); b – heterozygote (genotype AC); c – homozygote for the A allele (genotype AA).
Evaluation of the relative expression level of the RAB29 gene in peripheral blood mononuclear cells of PD patients and controls. Peripheral venous blood samples were collected from PD patients (N = 30) and control individuals (N = 43). Peripheral blood mononuclear cells (PBMCs) were isolated by density gradient centrifugation using Ficoll-Paque PLUS (GE Healthcare) at 400g for 40 minutes, as described by Böyum (1968). The resulting mononuclear fraction was washed twice with phosphate-buffered saline (PBS; Biolot, St. Petersburg) and centrifuged at 3,000 rpm for 10 minutes. Total RNA was extracted from PBMCs using the RNeasy Mini Kit (Qiagen, 74104, USA). Complementary DNA (cDNA) was synthesized via reverse transcription using the RevertAid First Strand cDNA Synthesis Kit (K1622, Thermo Scientific, Lithuania).
The relative expression level of the RAB29 gene in PBMCs of PD patients (N = 30) and control individuals (N = 43) was quantified by real-time PCR using SYBR Green I as the intercalating dye. The housekeeping genes RPLP0 and GAPDH, which are constitutively expressed in PBMCs, were used as internal reference genes for normalization. Primer sequences were designed using the Primer3 v. 0.4.0 program (https://bioinfo.ut.ee/primer3-0.4.0) (FOR: 5ʹ-CGGTTTCACAGGTTGGACAG-3ʹ, REV: 5ʹ-CC CTTGGGTGGACAAAGACA-3ʹ). The relative mRNA level for each gene was calculated by comparing the threshold amplification levels ΔΔCt (Livak, Schmittgen, 2001).
Evaluation of lysosomal hydrolase activities and lysosphingolipid concentrations in peripheral blood of PD patients and controls. Peripheral venous blood samples were collected from PD patients and controls into EDTA tubes. To obtain dried blood spots, 40 μl of whole blood was applied to each spot on a filter paper test blank, after which the spots were allowed to air dry at room temperature for 2 hours and were then stored at +4 °C until extraction. Activities of four lysosomal hydrolases: glucocerebrosidase (GCase), α-galactosidase (GLA), galactocerebrosidase (GALC), and sphingomyelinase (ASMase), as well as the concentration of three lysosphingolipids: hexazyl sphingosine (HexSph) (a mixture of glycosyl sphingosine (GlcSph) and galactosyl sphingosine (GalSph)), lysosphingomyelin (LysoSM) and lysoglobotriaosyl sphingosine (LysoGb3) were estimated by high-performance liquid chromatography coupled with tandem mass spectrometry (HPLC-MS/MS) according to our previously published protocol (Pchelina et al., 2018).
Statistical data processing. Statistical analyses were performed using the R programming environment (version 4.0.5). Odds ratios (OR) with 95 % confidence intervals (CIs) were calculated using logistic regression models adjusted for age and gender. The nonparametric Mann–Whitney U-test was applied to compare the relative expression levels of the RAB29 gene, lysosomal hydrolase activities, and lysosphingolipid concentrations between study groups. To evaluate the association between the rs823144 variant of the RAB29 gene and lysosomal hydrolase activity, multiple linear regression analysis was conducted, adjusting for age, gender, and disease duration. Statistical significance was set at p < 0.05. Data are presented as median (min–max).
Results
Association between rs823144 of the RAB29 gene and PD risk
Genotyping of the rs823144 variant in the RAB29 gene among PD patients and controls from the Northwestern Russian population revealed that the major allele is A. Hardy–Weinberg equilibrium (HWE) analysis confirmed that the genotype distributions in both groups were in equilibrium (p > 0.05). The genotype frequencies are summarized in Table 2. Logistic regression analysis demonstrated that the rs823144 variant is significantly associated with a reduced risk of Parkinson’s disease in this population (OR: 0.7806, 95 % CI: 0.6578–0.9263, p = 0.0046).
Frequencies of rs823144 genotypes and alleles in the study groupsNote. PD – Parkinson’s disease; OR – odds ratio.
Evaluation of the relative level of RAB29 gene mRNA in the mononuclear fraction of peripheral blood cells of PD patients and controls
This study assessed for the first time the association between the rs823144 variant of the RAB29 gene and the relative expression level of RAB29 mRNA in peripheral blood mononuclear cells (PBMCs) from PD patients and controls. The median relative expression of RAB29 mRNA in PBMCs was 1.00 (0.22–1.75) in PD patients and 0.96 (0.13–1.79) in controls, with no statistically significant difference between groups ( p > 0.05). When stratified by rs823144 genotype, the relative RAB29 mRNA levels were as follows: in PD patients, AA genotype carriers had a median expression of 1.09 (0.–1.68), while carriers of AC and CC genotypes combined showed 1.13 (0.36–1.75); in controls, AA carriers had 1.05 (0.58–1.28) and AC + CC carriers had 0.97 (0.55–1.33). No significant association was found between the rs823144 genotype and RAB29 mRNA levels in either group ( p > 0.05).
Association of the rs823144 variant of the RAB29 gene with the activity of lysosomal hydrolases and the concentration of lysosphingolipids in the peripheral blood of patients with PD and controls
This study is the first to investigate the association of the rs823144 variant of the RAB29 gene with lysosomal hydrolase activities and lysosphingolipid concentrations in the blood of PD patients and controls. Patients with PD exhibited significantly increased GALC activity and decreased LysoSM concentrations compared to controls (p = 0.008 and p = 0.01, respectively) (Table 3). Stratification by rs823144 genotype revealed that carriage of the C allele of the rs823144 variant in the RAB29 gene was associated with increased GLA activity and decreased levels of LysoGb3 – a substrate of GLA – in PD patients (p = 0.038 and p = 0.022, respectively) (Table 3). These associations were confirmed by multiple linear regression adjusted for gender, age, and disease duration (GLA: β = 1.11, p = 0.024; LysoGb3: β = –0.23, p = 0.015) (Table 4). No significant differences were observed in GCase, GLA and ASMase activities, or HexSph and LysoGb3 concentrations when analyzing the combined groups of PD patients and controls, regardless of rs823144 genotype. Similarly, no genotype-dependent differences were detected in GCase, ASMase, GALC activities, or HexSph concentrations.
*Lysosomal hydrolase activity and lysosphingolipid levels in the peripheral blood of PD patients and controlsNote. PD – Parkinson’s disease; GCase – glucocerebrosidase; GLA – α-galactosidase; GALC – galactocerebrosidase; ASMase – sphingomyelinase; HexSph – hexasylsphingosine; LysoSM – lysosphingomyelin; LysoGb3 – lysoglobotriaosylsphingosine.
- Compared with the combined control group; ** compared with PD patients with the AA genotype. Data are presented as median (min–max).*
Lysosomal hydrolase activities and lysosphingolipid concentrations in the peripheral blood of patients with PD (regression analysisNote. PD – Parkinson’s disease; GCase – glucocerebrosidase; GLA – α-galactosidase; GALC – galactocerebrosidase; ASMase – sphingomyelinase; HexSph – hexasylsphingosine; LysoSM – lysosphingomyelin; LysoGb3 – lysoglobotriaosylsphingosine.
Notably, PD patients carrying the C allele exhibited decreased blood concentrations of LysoGb3 and LysoSM compared to the combined control group ( p = 0.045 and p = 0.015, respectively), whereas PD patients with the AA genotype showed increased LysoSM levels relative to controls ( p = 0.022). The increase in GLA activity and corresponding decrease in its substrate LysoGb3 were specific to carriers of C allele of the rs823144 variant in the RAB29 gene within the PD group and were not observed in controls
Discussion
The RAB29 gene is one of five genes located within the PARK16 locus on chromosome 1q32, previously implicated in Parkinson’s disease risk (Simón-Sánchez et al., 2009; Tucci et al., 2010). Multiple association studies have identified the minor allele C of rs823144, positioned in the promoter region of RAB29, as protective against PD across various populations (Gan-Or et al., 2012; Xia et al., 2015; Khaligh et al., 2017; Sun et al., 2021). It has been hypothesized that this allele modulates transcription factor binding (Gan-Or et al., 2012; Khaligh et al., 2017).
In silico analyses suggest that the C allele of the rs823144 variant in the RAB29 gene abolishes the binding site for c- Ets-1 and introduces binding sites for p300, GATA-1, and Sp1, potentially enhancing RAB29 expression (Gan-Or et al., 2012). However, data from the GTEx database (https://www. gtexportal.org/) indicate that the CC genotype of rs823144 correlates with reduced RAB29 expression in whole blood and brain tissues ( p < 0.0001). In turn, increased expression of the RAB29 gene, encoding one of the main regulators of LRRK2 kinase, may potentially lead to impaired activation of this kinase. LRRK2, by phosphorylating Rab family proteins, regulates endolysosomal transport (Reczek et al., 2007; Wei et al., 2023). Dysregulation of LRRK2 activation can impair lysosomal hydrolase transport and function, contributing to PD pathogenesis
In our study of the Northwestern Russian population, we confirmed the association of the C allele of rs823144 with a reduced risk of PD, which is consistent with global findings (Gan-Or et al., 2012; Khaligh et al., 2017; Sun et al., 2021). Notably, we did not observe any association between the C allele of rs823144 genotype and RAB29 mRNA levels in peripheral blood mononuclear cells from either PD patients or controls.
Lysosomal dysfunction is widely recognized as a central mechanism in PD. Previous work by our group and others has demonstrated altered lysosomal hydrolase activity and sphingolipid metabolism in peripheral fluids and postmortem brain regions of PD patients (Alcalay et al., 2018; Nelson et al., 2018; Huebecker et al., 2019; Chang et al., 2022; Usenko et al., 2022, 2024). Specifically, we reported increased GALC activity and decreased LysoSM – a substrate of acid ASMase – concentration in PD peripheral blood (Usenko et al., 2022, 2024). In turn, disruption of lysosphingolipid metabolism, including through altered lysosomal hydrolase activity, may contribute to the aggregation of α-synuclein (Mazzulli et al., 2011; Marie et al., 2015).
Accumulating lysosphingolipids in neurons can stabilize neurotoxic alpha-synuclein oligomers (Battis et al., 2023). Prior brain autopsy studies revealed correlations between LysoGb3 isoform concentrations and pathological phosphorylated alpha-synuclein as well as an negative correlation of GLA activity with levels of α-synuclein phosphorylated at serine 129 – the pathological form of the protein that predominates in aggregates in PD (Nelson et al., 2018). Our novel findings linking the C allele of rs823144 to increased GLA activity and decreased LysoGb3 concentration, alongside elevated LysoSM levels in AA genotype carriers, underscore a potential role for RAB29 in sphingolipid metabolism in PD.
It should be noted that a marked reduction in GLA activity due to mutations in the GLA gene leads to a rare lysosomal storage disorder, Fabry disease. Notably, elevated LysoGb3 concentration is a risk factor for white matter lesions in Fabry disease. (Rombach et al., 2010). We have also observed LysoGb3 accumulation in neuronopathic mucopolysaccharidoses (Baydakova et al., 2020), suggesting that LysoGb3 elevation may not be exclusive to Fabry disease. Therefore, the association of the RAB29 rs823144 C allele with increased GLA activity and decreased LysoGb3 in PD patients may influence disease progression and clinical phenotype.
The present study has several limitations. The sample sizes of PD patients and the control group included in the experiment assessing the association of the RAB29 rs823144 variant with RAB29 mRNA levels were limited. The compared groups were population-wise heterogeneous. Furthermore, we did not directly assess the impact of rs823144 on LRRK2 kinase activity, which warrants investigation in future studies.
Conclusion
This study demonstrates for the first time worldwide that the C allele of the rs823144 variant in the RAB29 gene, previously identified as protective against PD in other populations, is associated with a reduced risk of PD in the North-West region of Russia. Additionally, we report for the first time that carriage of the C allele correlates with increased GLA activity and decreased LysoGb3 concentration in the blood of PD patients.
These findings suggest a potential role for RAB29 in lysosphingolipid metabolism and imply that the rs823144 variant may influence the clinical course of PD. Further research is warranted to elucidate the relationship between the PARK16 locus, RAB29 gene, lysosphingolipid metabolism, and PD progression.
Conflict of interest
The authors declare no conflict of interest.
References
Alcalay R.N., Levy O.A., Waters C.C., Fahn S., Ford B., Kuo S.H., Mazzoni P., … Wolf P., Oliva P., Keutzer J., Marder K., Zhang X. Glucocerebrosidase activity in Parkinson’s disease with and without GBA mutations. Brain. 2015;138(9):2648-2658. doi 10.1093/brain/ awv179
Alcalay R.N., Wolf P., Levy O.A., Kang U.J., Waters C., Fahn S., Ford B., … Chung W.K., Oliva P., Keutzer J., Marder K., Zhang X.K. Alpha galactosidase A activity in Parkinson’s disease. Neurobiol Dis. 2018;112:85-90. doi 10.1016/j.nbd.2018.01.012Battis K., Xiang W., Winkler J. The bidirectional interplay of α-synuclein with lipids in the central nervous system and its implications for the pathogenesis of Parkinson’s disease. Int J Mol Sci. 2023; 24(17):13270. doi 10.3390/ijms241713270
Baydakova G., Ilyushkina A., Gaffke L., Pierzynowska K., Bychkov I., Ługowska A., Wegrzyn G., Tylki-Szymanska A., Zakharova E. Elevated LysoGb3 concentration in the neuronopathic forms of mucopolysaccharidoses. Diagnostics (Basel). 2020;10(3):155. doi 10.3390/diagnostics10030155
Böyum A. Isolation of mononuclear cells and granulocytes from human blood. Isolation of monuclear cells by one centrifugation, and of granulocytes by combining centrifugation and sedimentation at 1 g. Scand J Clin Lab Invest Suppl. 1968;97:77-89
Chang K.H., Cheng M.L., Tang H.Y., Huang C.Y., Wu H.C., Chen C.M. Alterations of sphingolipid and phospholipid pathways and ornithine level in the plasma as biomarkers of Parkinson’s disease. Cells. 2022;11(3):395. doi 10.3390/cells11030395
Galper J., Dean N.J., Pickford R., Lewis S.J.G., Halliday G.M., Kim W.S., Dzamko N. Lipid pathway dysfunction is prevalent in patients with Parkinson’s disease. Brain. 2022;145(10):3472-3487. doi 10.1093/brain/awac176
Gan-Or Z., Bar-Shira A., Dahary D., Mirelman A., Kedmi M., Gurevich T., Giladi N., Orr-Urtreger A. Association of sequence alterations in the putative promoter of RAB7L1 with a reduced Parkinson disease risk. Arch Neurol. 2012;69(1):105-110. doi 10.1001/arch neurol.2011.924
Huebecker M., Moloney E.B., Van Der Spoel A.C., Priestman D.A., Isacson O., Hallett P.J., Platt F.M. Reduced sphingolipid hydrolase activities, substrate accumulation and ganglioside decline in Parkinson’s disease. Mol Neurodegener. 2019;14(1):40. doi 10.1186/ s13024-019-0339-z
Kedariti M., Frattini E., Baden P., Cogo S., Civiero L., Ziviani E., Zilio G., … Di Fonzo A., Alcalay R.N., Rideout H., Greggio E., Plotegher N. LRRK2 kinase activity regulates GCase level and enzymatic activity differently depending on cell type in Parkinson’s disease. NPJ Parkinsons Dis. 2022;8(1):92. doi 10.1038/s41531- 022-00354-3
Khaligh A., Goudarzian M., Moslem A., Mehrtash A., Jamshidi J., Darvish H., Emamalizadeh B. RAB7L1 promoter polymorphism and risk of Parkinson’s disease; a case-control study. Neurol Res. 2017; 39(5):468-471. doi 10.1080/01616412.2017.1297558
Kuwahara T., Iwatsubo T. The emerging functions of LRRK2 and Rab GTPases in the endolysosomal system. Front Neurosci. 2020;14: 227. doi 10.3389/fnins.2020.00227
Li H., Durbin R. Fast and accurate short read alignment with Bur-rows– Wheeler transform. Bioinformatics. 2009;25(14):1754-1760. doi 10.1093/bioinformatics/btp324
Lill C.M. Genetics of Parkinson’s disease. Mol Cell Probes. 2016; 30(6):386-396. doi 10.1016/j.mcp.2016.11.001
Liu Z., Bryant N., Kumaran R., Beilina A., Abeliovich A., Cookson M.R., West A.B. LRRK2 phosphorylates membrane-bound Rabs and is activated by GTP-bound Rab7L1 to promote recruitment to the trans-Golgi network. Hum Mol Genet. 2018;27(2):385- 395. doi 10.1093/hmg/ddx410
Livak K.J., Schmittgen T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods. 2001;25(4):402-408. doi 10.1006/meth.2001.1262
MacLeod D.A., Rhinn H., Kuwahara T., Zolin A., Di Paolo G., MacCabe B.D., Marder K.S., Honig L.S., Clark L.N., Small S.A., Abeliovich A. Rab7L1 interacts with LRRK2 to modify intraneuronal protein sorting and Parkinson’s disease risk. Neuron. 2013; 77(3):425-439. doi 10.1016/j.neuron.2012.11.033
Madero-Pérez J., Fdez E., Fernández B., Lara Ordóñez A.J., Blanca Ramírez M., Gómez-Suaga P., Waschbüsch D., … Beilina A., Gonnelli A., Cookson M.R., Greggio E., Hilfiker S. Parkinson disease-associated mutations in LRRK2 cause centrosomal defects via Rab8a phosphorylation. Mol Neurodegener. 2018;13(1):3. doi 10.1186/S13024-018-0235-
Maniatis T., Fritsch E.F., Sambrook J. Methods of Genetic Engineering. Molecular Cloning. Moscow: Mir Publ., 1984 (in Russian)
Marie G., Dunning C.J., Gaspar R., Grey C., Brundin P., Sparr E., Linse S. Acceleration of α-synuclein aggregation by exosomes. J Biol Chem. 2015;290(5):2969. doi 10.1074/jbc.M114.585703
Mazzulli J.R., Xu Y.H., Sun Y., Knight A.L., McLean P.J., Caldwell G.A., Sidransky E., Grabowski G.A., Krainc D. Gaucher disease glucocerebrosidase and α-synuclein form a bidirectional pathogenic loop in synucleinopathies. Cell. 2011;146(1):37-52. doi 10.1016/j.cell.2011.06.001
McKenna A., Hanna M., Banks E., Sivachenko A., Cibulskis K., Kernytsky A., Garimella K., Altshuler D., Gabriel S., Daly M., DePristo M.A. The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010;20(9):1297-1303. doi 10.1101/gr.107524.110
Nalls M.A., Blauwendraat C., Vallerga C.L., Heilbron K., Bandres- Ciga S., Chang D., Tan M., … Silburn P.A., Vallerga C.L., Wallace L., Wray N.R., Zhang F. Identification of novel risk loci, causal insights, and heritable risk for Parkinson’s disease: a meta-genome wide association study. Lancet Neurol. 2019;18(12):1091-1102. doi 10.1016/S1474-4422(19)30320-5
Nechushtai L., Frenkel D., Pinkas-Kramarski R. Autophagy in Parkinson’s disease. Biomolecules. 2023;13(10):1435. doi 10.3390/biom 13101435
Nelson M.P., Boutin M., Tse T.E., Lu H., Haley E.D., Ouyang X., Zhang J., Auray-Blais C., Shacka J.J. The lysosomal enzyme alpha- Galactosidase A is deficient in Parkinson’s disease brain in association with the pathologic accumulation of alpha-synuclein. Neurobiol Dis. 2018;110:68-81. doi 10.1016/j.nbd.2017.11.006
Pchelina S., Baydakova G., Nikolaev M., Senkevich K., Emelyanov A., Kopytova A., Miliukhina I., Yakimovskii A., Timofeeva A., Berkovich O., Fedotova E., Illarioshkin S., Zakharova E. Blood lysosphingolipids accumulation in patients with Parkinson’s disease with glucocerebrosidase 1 mutations. Mov Disord. 2018;33(8):1325-1330. doi 10.1002/mds.27393
Pihlstrøm L., Rengmark A., Bjørnarå K.A., Dizdar N., Fardell C., Forsgren L., Holmberg B., Larsen J.P., Linder J., Nissbrandt H., Tysnes O.B., Dietrichs E., Toft M. Fine mapping and resequencing of the PARK16 locus in Parkinson’s disease. J Hum Genet. 2015; 60(7):357-362. doi 10.1038/jhg.2015.34
Purlyte E., Dhekne H.S., Sarhan A.R., Gomez R., Lis P., Wightman M., Martinez T.N., Tonelli F., Pfeffer S.R., Alessi D.R. Rab29 activation of the Parkinson’s disease-associated LRRK2 kinase. EMBO J. 2018;37(1):1-18. doi 10.15252/EMBJ.201798099
Rausch T., Fritz M.H.Y., Untergasser A., Benes V. Tracy: basecalling, alignment, assembly and deconvolution of sanger chromatogram trace files. BMC Genomics. 2020;21(1):230. doi 10.1186/S12864- 020-6635-8
Reczek D., Schwake M., Schröder J., Hughes H., Blanz J., Jin X., Brondyk W., Van Patten S., Edmunds T., Saftig P. LIMP-2 is a receptor for lysosomal mannose-6-phosphate-independent targeting of betaglucocerebrosidase. Cell. 2007;131(4):770-783. doi 10.1016/j.cell. 2007.10.018
Rivero-Ríos P., Romo-Lozano M., Fernández B., Fdez E., Hilfiker S. Distinct roles for Rab10 and Rab29 in pathogenic LRRK2-mediated endolysosomal trafficking alterations. Cells. 2020;9(7):1719. doi 10.3390/cellS9071719
Rombach S.M., Dekker N., Bouwman M.G., Linthorst G.E., Zwinderman A.H., Wijburg F.A., Kuiper S., vd Bergh Weerman M.A., Groener J.E., Poorthuis B.J., Hollak C.E., Aerts J.M. Plasma globotriaosylsphingosine: diagnostic value and relation to clinical manifestations of Fabry disease. Biochim Biophys Acta. 2010;1802(9): 741-748. doi 10.1016/j.bbadis.2010.05.003
Rudakou U., Yu E., Krohn L., Ruskey J.A., Asayesh F., Dauvilliers Y., Spiegelman D., … Rouleau G.A., Hassin-Baer S., Fon E.A., Alcalay R.N., Gan-Or Z. Targeted sequencing of Parkinson’s disease loci genes highlights SYT11, FGF20 and other associations. Brain. 2021;144(2):462-472. doi 10.1093/brain/awaa401
Satake W., Nakabayashi Y., Mizuta I., Hirota Y., Ito C., Kubo M., Kawaguchi T., … Yamamoto M., Hattori N., Murata M., Nakamura Y., Toda T. Genome-wide association study identifies common variants at four loci as genetic risk factors for Parkinson’s disease. Nat Genet. 2009;41(12):1303-1307. doi 10.1038/ng.485
Shi J.J., Mao C.Y., Guo Y.Z., Fan Y., Hao X.Y., Li S.J., Tian J., … Zuo C.Y., Liang Y.Y., Xu Y.M., Yang J., Shi C.H. Joint analysis of proteome, transcriptome, and multi-trait analysis to identify novel Parkinson’s disease risk genes. Aging. 2024;16(2):1555-1580. doi 10.18632/aging.205444
Simón-Sánchez J., Schulte C., Bras J.M., Sharma M., Gibbs J.R., Berg D., Paisan-Ruiz C., … Chen H., Riess O., Hardy J.A., Singleton A.B., Gasser T. Genome-wide association study reveals genetic risk underlying Parkinson’s disease. Nat Genet. 2009;41(12):1308- 1312. doi 10.1038/ng.487
Steger M., Tonelli F., Ito G., Davies P., Trost M., Vetter M., Wachter S., … Fell M.J., Morrow J.A., Reith A.D., Alessi D.R., Mann M. Phosphoproteomics reveals that Parkinson’s disease kinase LRRK2 regulates a subset of Rab GTPases. eLife. 2016;5:e12813. doi 10.7554/eLife.12813.001
Sun J., Deng L., Zhu H., Liu M., Lyu R., Lai Q., Zhang Y. Meta-analysis on the association between rs11868035, rs823144, rs3851179 and Parkinson’s disease. Meta Gene. 2021;30:100949. doi 10.1016/ j.mgene.2021.100949
Tucci A., Nalls M.A., Houlden H., Revesz T., Singleton A.B., Wood N.W., Hardy J., Paisán-Ruiz C. Genetic variability at the PARK16 locus. Eur J Hum Genet. 2010;18(12):1356-1359. doi 10.1038/ejhg.2010.125
Usenko T.S., Senkevich K.A., Bezrukova A.I., Baydakova G.V., Basharova K.S., Zhuravlev A.S., Gracheva E.V., … Palchikova E.I., Zalutskaya N.M., Emelyanov A.K., Zakharova E.Y., Pchelina S.N. Impaired sphingolipid hydrolase activities in dementia with Lewy bodies and multiple system atrophy. Mol Neurobiol. 2022;59(4): 2277-2287. doi 10.1007/S12035-021-02688-0
Usenko T.S., Senkevich K.A., Basharova K.S., Bezrukova A.I., Baydakova G.V., Tyurin A.A., Beletskaya M.V., Kulabukhova D.G., Grunina M.N., Emelyanov A.K., Miliukhina I.V., Timofeeva A.A., Zakharova E.Y., Pchelina S.N. LRRK2 exonic variants are associated with lysosomal hydrolase activities and lysosphingolipid alterations in Parkinson’s disease. Gene. 2023;882:147639. doi 10.1016/j.gene.2023.147639
Usenko T.S., Timofeeva A., Beletskaia M., Basharova K., Baydakova G., Bezrukova A., Grunina M., Emelyanov A., Miliukhina I., Zakharova E., Pchelina S. The effect of p.G2019S mutation in the LRRK2 gene on the activity of lysosomal hydrolases and the clinical features of Parkinson’s disease associated with p.N370S mutation in the GBA1 gene. J Integr Neurosci. 2024;23(1):16. doi 10.31083/ j.jin2301016
Vincze T., Posfai J., Roberts R.J. NEBcutter: a program to cleave DNA with restriction enzymes. Nucleic Acids Res. 2003;31(13):3688- 3691. doi 10.1093/nar/gkg526
Wang S., Ma Z., Xu X., Wang Z., Sun L., Zhou Y., Lin X., Hong W., Wang T. A role of Rab29 in the integrity of the trans-golgi network and retrograde trafficking of mannose-6-phosphate receptor. PLoS One. 2014;9(5):e96242. doi 10.1371/journal.pone.0096242
Wei Y., Awan M.U.N., Bai L., Bai J. The function of Golgi apparatus in LRRK2-associated Parkinson’s disease. Front Mol Neurosci. 2023; 16:1097633. doi 10.3389/fnmol.2023.1097633
Xia H., Luo Q., Li X.X., Yang X.L. Association between PARK16 gene polymorphisms and susceptibility of Parkinson’s disease in a Chinese population. Genet Mol Res. 2015;14(2):2978-2985. doi 10.4238/2015.april.10.7
Ysselstein D., Nguyen M., Young T.J., Severino A., Schwake M., Merchant K., Krainc D. LRRK2 kinase activity regulates lysosomal glucocerebrosidase in neurons derived from Parkinson’s disease patients. Nat Commun. 2019;10(1):5570. doi 10.1038/s41467-019- 13413-w
Zhu H., Tonelli F., Turk M., Prescott A., Alessi D.R., Sun J. Rab29- dependent asymmetrical activation of leucine-rich repeat kinase 2. Science. 2023;382(6677):1404-1411. doi 10.1126/science.adi9926
Zimprich A., Biskup S., Leitner P., Lichtner P., Farrer M., Lincoln S., Kachergus J., Hulihan M., Uitti R.J., Calne D.B., Stoessl A.J. Mutations in LRRK2 cause autosomal-dominant parkinsonism with pleomorphic pathology. Neuron. 2004;44(4):601-607. doi 10.1016/ j.neuron.2004.11.005