Using polygenic scores to assess liability to antisocial behavior
A.V. Kazantseva, D.V. Yakovleva, Yu.D. Davydova, E.K. Khusnutdinova

TL;DR
This study uses genetic and social factors to assess liability to antisocial behavior in a Russian population, finding that social factors have a stronger influence than genetic scores.
Contribution
The study evaluates polygenic scores and social factors for antisocial behavior in a Russian cohort, a population not previously studied in this context.
Findings
Social factors like traumatic brain injury and tobacco smoking significantly increased liability to antisocial behavior.
Polygenic scores alone explained only 1.1–1.5% of the variance in antisocial behavior liability.
The best model combined genetic and social factors, explaining 16.2–21.2% of the variance.
Abstract
To date, several genome-wide association studies (GWAS) of antisocial behavior (ASB) have been conducted in Europeans, which promoted research aimed at evaluating liability to ASB-related phenotypes in independent samples. Such studies implemented a polygenic score (PGS) approach, which represents a composite score considering a number of “risky” alleles. Since no GWAS of ASB has been conducted in Russians, the present study aimed to perform a replication study of liability to severe criminal behavior (homicide) in individuals from Russia using PGS. Moreover, we sought to obtain the best model considering PGS and potential social factors as predictors. Genotyping of the “top” ten SNPs previously identified in GWAS meta-analysis of ASB (CADM2, REV3L, FOXP1, FOXP2, BDNF, FURIN, XKR6, TMEM18, SORCS3, and ZIC4 genes) was conducted via real-time PCR in 227 homicide offenders and 254 healthy…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
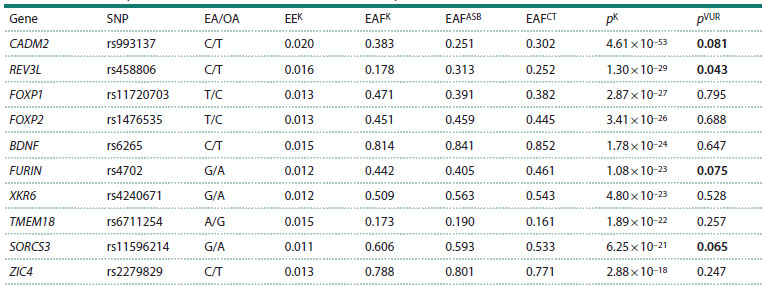 Table 1
Table 1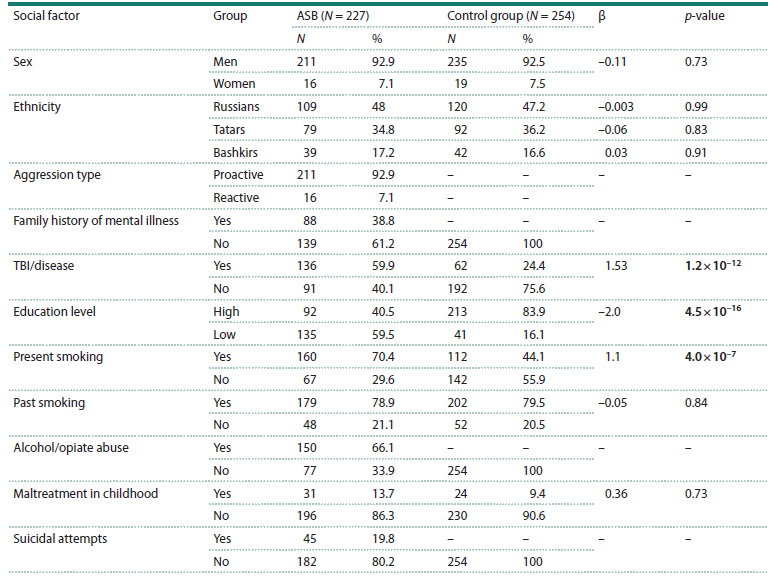 Table 2
Table 2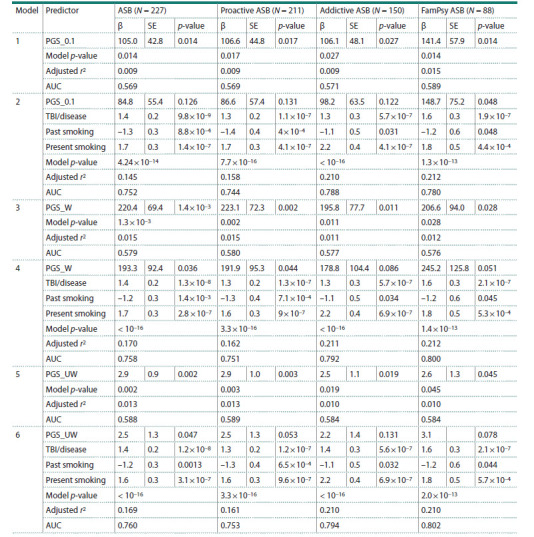 Table 3
Table 3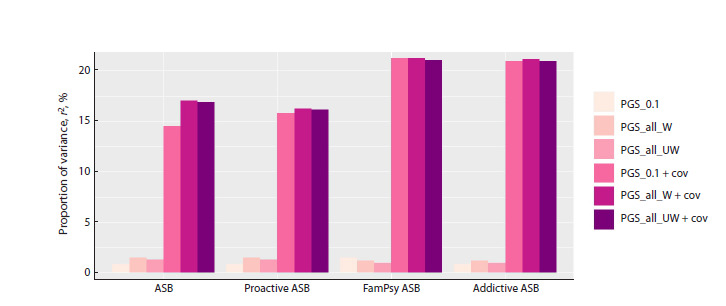 Fig. 1
Fig. 1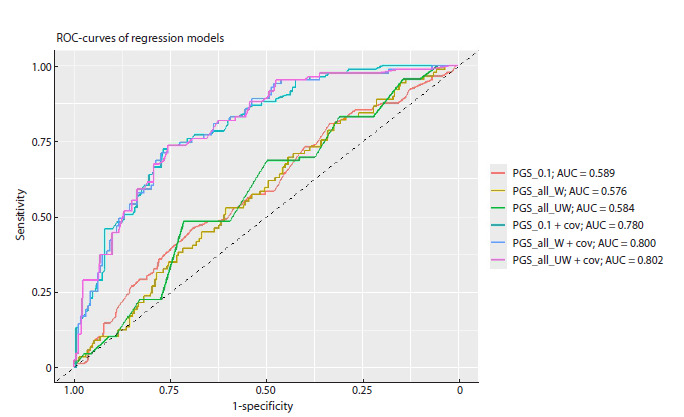 Fig. 2
Fig. 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic Associations and Epidemiology · Cognitive Abilities and Testing · Psychopathy, Forensic Psychiatry, Sexual Offending
Introduction
Aggressive behavior (AB) and antisocial behavior (ASB) represent a destructive form of social interaction aimed at causing damage to another object and resulting in its frustration. From the evolutionary point of view, enhanced aggression was required for the survival of human groups (Baron, Richardson, 2004), thus promoting certain biological benefits. Although it is suggested that the aggression level in modern society is decreased compared with early humans, it still remains significant. To be more precise, the level of severe crimes, including homicides and intentional inflictions of severe harm, accounted for 117.3 and 567.1 thousand cases in 2022 in Russia (according to the data from the Ministry of Internal Affairs of the Russian Federation, http://www.crimestat.ru). According to the data from the World Health Organization (https://www.who.int/data), the homicide rate remains significant worldwide and was estimated at 5.8 cases per 100,000 of population in 2021 in the United States (in comparison, 6.7 cases in Russia; 0.5–4.5 cases in Europe; 5–100 cases in South American countries, and 5–20 cases in Africa).
In turn, during past years, several specifically cruel cases of murder, domestic violence, and antisocial behavior at schools have shocked Russia and the neighboring countries. However, it remains impossible to predict the occurrence of severe cruelty before the crimes have been conducted. In this regard, it seems important to determine significant factors underlying ASB, which can help to predict a higher probability of manifesting cruelty and antisocial behavior. It should be noted that ASB usually manifests in the form of certain psychiatric diseases, including oppositional defiant disorder, conduct disorder, and antisocial personality disorder (Pezzoli et al., 2025). Therefore, these phenotypes can share etiology and underlying factors.
According to previous research, the main factors predisposing to ASB or related phenotypes include biological, psychological, and environmental ones (Fritz et al., 2023). Examination of biological factors, which contribute 50 to 80 % of variance in aggression (Manchia, Fanos, 2017; Odintsova et al., 2023), is mainly focused on the analysis of genetic and epigenetic effects. Logically, genetic variants (SNPs) in the genes attributed to neurotransmitter release, reuptake, and binding (Davydova et al., 2020a; Antón-Galindo et al., 2023), oxytocin and arginine vasopressin signaling (Davydova et al., 2020b; Kazantseva et al., 2021), and others (Pezzoli et al., 2025) have been tested for their relation to individual variance in aggressive behavior. However, the results of multiple studies demonstrate inconsistent findings. Another methodological approach, i. e., genome-wide association studies (GWAS), enables to identify associated SNPs under a hypothesis-free paradigm. Although to date several GWASs of antisocial behavior have been carried out, these studies differ in the examined phenotypes frequently linked with ASB (combined phenotype of externalizing behavior (Karlsson Linnér et al., 2021), impulsivity (Deng et al., 2023), problems with selfregulation (Heilbronner et al., 2021), irritability (Mbatchou et al., 2021), risky behavior (Karlsson Linnér et al., 2019)) or age groups (children (Pappa et al., 2016), adults (Tielbeek et al., 2017)). Moreover, summarized findings from ~ 1.5 million subjects identified more than 500 SNPs related to liability to externalizing behavior, including antisocial behavior, attention- deficit/hyperactivity disorder (ADHD), and addiction in a European cohort (Karlsson Linnér et al., 2021).
One of the possible applications of GWAS findings is to use them for the calculation of polygenic scores (PGS) on the basis of effect estimates obtained for each SNP in the training sample. In turn, inclusion of PGS in mathematical models can gain prediction of enhanced risk of certain complex phenotypes. To date, several attempts seeking to replicate GWAS findings in an independent sample using PGS from ASB phenotype have been made (Karlsson Linnér et al., 2021; Li et al., 2023; Tesli et al., 2024; Acland et al., 2025), which succeeded in determining some proportion of variance in liability to conduct disorder, substance use disorders, smoking, ADHD, criminal behavior, depression, posttraumatic stress disorder, unemployment, and suicidal attempts. One of the possible limitations of using PGS for predicting ASB is the ethnic origin of the examined population, since differences in allele and genotype frequencies between ethnic groups can change SNPs’ effect (Kazantseva et al., 2016). To date, no GWAS of liability to homicidal conduct has been carried out in subjects from Russia. Therefore, it is relevant to check if it is applicable to use the effect estimates obtained from combined ASB phenotype and different ethnic groups to predict the probability of conducting severe ASB in the Russian cohort.
Undoubtedly, specific environmental/social factors acting at various stages of ontogenesis affect genes’ activity via epigenetic changes in regulation of genes responsible for manifesting aggression (Borinskaya et al., 2021). In this context, the analysis of potential social factors together with genetic effects (PGS) can help to increase the prognostic significance of the final model. In addition, it is established that ASB is highly accumulated in certain groups, including subjects with comorbid mental disorders (Ip et al., 2021; Wang et al., 2024; Pezzoli et al., 2025), family history of mental illness (Han et al., 2024), addiction (Karlsson Linnér et al., 2021; Antón-Galindo et al., 2023), and unfavorable rearing conditions (Burt, 2022).
Considering the existing findings of ASB meta-analysis of European populations (Karlsson Linnér et al., 2021) and absent GWAS data for individuals from Russia, the present study aimed to evaluate the applicability of calculated polygenic scores based on existing GWAS data to predict severe ASB (homicide) in the Russian cohort. Moreover, to enhance the prognostic ability of regression models, we sought to obtain the best model with the optimal sensitivity and specificity, which assumes PGS and potential social factors as predictors
Materials and methods
The study sample comprised 227 criminal offenders who conducted homicide and were directed to a forensic examination of present mental disorders in the Republican Clinical Psychiatric Hospital (Ufa, Russia). Only individuals without mental illness who were proven to be sane by the Court were included in the study. The examined sample consisted mainly of men (93 %) with a mean age of 41.5 ± 14.5 years. Ethnic content of the sample was the following: 48 % Russians, 34.8 % Tatars, and 17.2 % Bashkirs. The data on the social/ clinical background of enrolled subjects were obtained via a survey and included the information on present and past tobacco smoking, alcohol/opiate abuse, family history of mental illness or criminal behavior, suicidal attempts, level of education, maltreatment in childhood, severe chronic disease in anamnesis, and type of ASB (proactive or reactive aggression).
The control group was selected on the basis of correspondence to the group of criminal offenders by age, ethnicity, and gender. In total, we examined DNA samples obtained from 254 individuals who reported no family history of mental illness and were non-registered in the psychiatric database of the Republic of Bashkortostan. The study was approved by the local bioethical committee at the Institute of Biochemistry and Genetics – Subdivision of the Ufa Federal Research Centre of the Russian Academy of Sciences (Ufa, Russia) (protocol code 15, date of approval, October 12, 2017) in accordance with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards.
SNP selection for PGS calculation from GWAS meta-analysis of ASB (Karlsson Linnér et al., 2021) was based on the following criteria: the lowest level of significance ( p <10–18); selection of a single SNP from a set of proxy SNPs; minor allele frequency (MAF) above 0.05 in Europeans (based on 1000 Genomes); and known regulatory effect of the SNP based on the RDB (Regulome Database, https://regulomedb. org/regulome-search) and CADD (Combined Annotation Dependent Depletion, https://cadd.gs.washington.edu) databases. The final list of selected SNPs included CADM2 rs993137, REV3L rs458806, FOXP1 rs11720703, FOXP2 rs1476535, BDNF rs6265, FURIN rs4702, XKR6 rs4240671, TMEM18 rs6711254, SORCS3 rs11596214, and ZIC4 rs2279829, which were used for PGS calculation, and is reported in Table 1. Genotyping of previously extracted DNA of the control group and criminal offenders was carried out using real-time PCR with KASP chemistry (LGC Genomics, UK).
Examined top SNPs linked to antisocial behavior: data from previous ASB GWAS and the VUR cohort
All examined SNPs corresponded to the Hardy–Weinberg equilibrium ( p > 0.05). At the second stage, we calculated PGS based on effect estimates obtained from R. Karlsson Linnér et al. (2021). Namely, PGS for each individual from our sample was calculated on the basis of inclusion of 1) SNPs under p < 0.1 (“weighted” effect), 2) all SNPs (using “weighted” effect), 3) all SNPs (using “unweighted” effect). Calculation of “weighted” and “unweighted” PGS was previously explained in detail (Kazantseva et al., 2023a). Briefly, individual PGS was calculated as the weighted/unweighted sum of the number of effect alleles at a certain SNP multiplied by the effect estimate (PLINK v.1.09).
Subsequently, a series of multiple logistic regressions was performed to obtain models that can predict liability to ASB in the total groups of homicide offenders, as well as in subgroups of subjects with proactive forms of aggression, comorbid substance use, or known family history of mental illness or criminal behavior. Initially, only PGS as a predictor was included, which was followed by a backward selection procedure to obtain a list of statistically significant social parameters to be included as predictors together with PGS (R v.4.4.2). To select the best predicting model, we have compared data on the lowest p-value, the highest proportion of variance (Nagelkerke pseudo-R2) explaining liability to ASB, and the highest area under the ROC curve (AUC) for each model.
Results
At the initial stage of the study, we examined the presence of significant differences between the criminal offenders and the control group in the examined social factors (Table 2). We have observed the differences in the proportion of individuals characterized by severe somatic diseases and traumatic brain injuries in anamnesis ( p = 1.2 × 10–12), depending on education level ( p = 4.5 × 10–16) and present smoking ( p = 4.0 × 10–7) between the groups.
Characteristics of the examined groups of criminal offenders and healthy donors and analysis of differences in social factors between the groupsNote. Statistically significant differences between the groups based on p-value < 0.05 are shown in bold. TBI/disease – traumatic brain injury or severe chronic disease in anamnesis. Dashes indicate non-applicable data.
For the genetic part of the present study we selected the “top” ten SNPs ( p < 2.9 × 10–18) identified in the previous meta-analysis GWAS of ASB (Karlsson Linnér et al., 2021). Effect estimates for alleles used for calculation of “weighted PGS” as well as effect allele frequencies in the VUR sample are given in Table 1. In addition, we have tested for statistically significant differences in allele frequencies of examined SNPs between criminal offenders and the control group in the examined cohort from the VUR, which enabled us to confirm a coincidence of four SNPs, although at a trend level ( p < 0.1): CADM2 rs993137, REV3L rs458806, FURIN rs4702, and SORCS3 rs11596214.
Primary logistic regression models that included PGS (based on four SNPs) revealed a small proportion of variance in liability to antisocial behavior in the total group (r2 = 0.9 %, p = 0.014), among subjects with a proactive form of aggression (r2 = 0.9 %, p = 0.017), with comorbid substance abuse (r2 = 0.9 %, p = 0.027), and with a family history of mental illness (r2 = 1.5 %, p = 0.014) (Table 3, Fig. 1). At the initial stage of regression analysis, we have included all social factors, including sex and ethnicity as covariates, together with PGS.
Regression models of liability to ASB based on a polygenic score and social factors as predictorsNote. ASB – antisocial behavior; FamPsy ASB – ASB in individuals with a family history of mental illness; β – regression coefficient for each predictor in the model; SE – standard error of β; AUC – area under curve; TBI/disease – traumatic brain injury or severe chronic disease in anamnesis. PGS_0.1 was based on effect estimates for REV3L rs458806, FOXP1 rs11720703, XKR6 rs4240671, and SORCS3 rs11596214; PGS_W and PGS_UW were PGS based on “weighted” and “unweighted” effect estimates for ten SNPs, correspondingly.
Proportion of variance (adjusted r2) in liability to antisocial behavior in the examined cohort explained by predictors included in multiple regression models based on PGS calculation of SNPs with p < 0.1 (PGS_0.1), “weighted effects” of all SNPs (PGS_all_W), “unweighted effects” of all SNPs (PGS_all_UW) with inclusion of social predictors (PGS_0.1 + cov, PGS_all_W + cov, PGS_all_UW + cov).Examined groups of ASB: total group of homicide offenders (ASB); homicide offenders with a proactive type of ASB (Proactive ASB), family history of mental illness or criminal behavior (FamPsy ASB), or substance abuse (Addictive ASB).
As expected, inclusion of potential social parameters as predictors enabled an increase in the statistical significance of the models, which resulted in 14.5 % (ASB), 15.8 % (proactive ASB), 21.0 % (ASB with comorbid addiction), and 21.2 % of variance (ASB with family history of mental illness) being explained. It should be mentioned that valuable social factors comprised of traumatic brain injury (TBI) or severe chronic disease in anamnesis (β = 1.4, p = 9.8 × 10–9) and present smoking (β = 1.7, p = 1.4 × 10–7) were associated with enhanced liability to aggression, while past smoking demonstrated a positive effect on ASB decrease (β = –1.3, p = 8.8 × 10–4). The impact of other social factors together with sex and ethnicity remained insignificant after the backward selection procedure. Therefore, inclusion of the mentioned social parameters allowed us to explain up to 16.1 % of variance in developing ASB. According to determined models, we can conclude that they possess the highest prediction ability for developing ASB in individuals who have relatives with mental disorders or criminal behavior (AUC = 0.780) or have alcohol/opiate addiction (AUC = 0.788) (Table 3).
At the second stage of our analysis, we calculated PGS based on effect estimates for all examined SNPs, even if they were non-significant in the VUR sample (Table 1). Therefore regression models, which implemented “weighted” (PGS_W) and “unweighted” (PGS_UW) PGS, slightly enhanced the proportion of variance in liability to ASB compared to previous models 1 and 2 (Table 2). Namely, a combined effect of ten genetic variants explained 1.1–1.5 % (“weighted effect”) and 1.0–1.3 % (“unweighted effect”) of variance in predisposition to homicide violence. Previously mentioned social predictors remained significant and, together with PGS, enabled to enhance the proportion of variance explained (16.2–21.2 % in “weighted” PGS, 16.1–21.0 % in “unweighted” PGS).
However, it seems that the inclusion of a larger number of non-significant SNPs had a very small effect on improving the predicting abilities of the models. Nevertheless, models with ten vs. four SNPs in PGS demonstrated slightly higher prognostic ability for ASB in the total sample and in individuals with a proactive form of aggressive behavior or comorbid substance abuse (Table 2, Models 4, 6). We have also constructed ROC curves and calculated comparative areas under the curves (AUCs) for all analyzed models (Fig. 2). Finally, our findings indicate that the best regression model has higher prognostic ability (r2 = 21 %) and a moderate measure of classifier performance (AUC = 0.802) to designate subjects at high risk for developing ASB if they have family history of mental disorders.
ROC curves and areas under the curves (AUCs) for various regression models predicting liability to manifest ASB in individuals with a family history of mental illness or criminal behavior (abbreviations are given as in Fig. 1).
Discussion
In the present study, we tested different logistic regression models, which were based on calculated polygenic scores, aimed at predicting liability to homicide in individuals from the Volga-Ural region of Russia. Based on our findings, the highest prediction ability for developing ASB was observed for individuals with a family history of mental disorders and those with substance abuse comorbidity. The data revealed are not surprising, since externalizing pathology is frequently accumulated in families (Acheson et al., 2018; Han et al., 2024) due to shared genetic profiles between biological parents and offspring. On the other hand, it was reported that the same genes/genetic variants were linked to different psychiatric conditions, addiction, and antisocial behavior (Ip et al., 2021; Antón-Galindo et al., 2023; Li et al., 2023; Wang et al., 2024; Pezzoli et al., 2025), which can be explained by genes’ pleiotropy in various complex traits (Watanabe et al., 2019).
Since no significant difference in predicting ASB risk in the VUR sample was observed among models based on “weighted” and “unweighted” effects of SNPs, it can be concluded that effect estimates from GWAS of Europeans seem to be inappropriate for individuals from Russia. Therefore, future research should be focused on conducting GWAS of ASB in a Russian cohort followed by verification in the same-ethnicity independent sample. Published studies, which sought to replicate findings obtained for different populations, succeeded in using PGS from ASB to predict liability to externalizing behavior in both Europeans and African Americans (Brislin et al., 2024), although representing a small cumulative effect of genetic variants.
Our findings indicate a very small impact of selected SNPs on predicting ASB, which was based on the effect estimates from the study of summarized phenotype of externalizing pathology. The data obtained support previous findings on the small effect (0.1–4.0 %) of analyzed genetic variants (even at a genome-wide level) as polygenic scores on predicting ASB (Tielbeek et al., 2017, 2022; Tesli et al., 2024). Our previous research also revealed a small proportion of variance explained in aggression level in a general population of Russia, which was attributed to the combined effect of 30 genetic variants (Kazantseva et al., 2023b).
It is known that environmental factors play a modulating role in establishing specific patterns of behavior (Kazantseva et al., 2014), including ASB-related ones. In particular, harsh parenting (Burt, 2022), school violence (Acland et al., 2025), and affiliation with delinquent peers (Schwartz et al., 2019) were assumed to increase a risk for manifesting ASB. Regression models designed in the present study also point to a more pronounced effect of environmental factors in establishing ASB than that of the genetic component. These findings are at odds with existing studies, which also depicted valuable impact of such social factors as community violence (Musci et al., 2019), harsh parenting (Acland et al., 2025), and low parental education level (Barnes et al., 2019) under gene-byenvironment interactions
In the present study, we have observed a significant effect of present smoking and history of traumatic brain injury/severe chronic disorders on manifesting criminal behavior. One of the probable links between smoking and ASB is attributed to the influence of nicotine on the CNS via exaggerated stress sensitivity (Weltens et al., 2021) and changed in epigenetic regulation (Gould et al., 2023). It should be noted that the usually accepted environmental effects can also be due to the impact of certain genetic and epigenetic profiles, which are inherited (McAdams et al., 2013). In this regard, present smoking may represent the result of activity of the genes responsible for developing addiction and externalizing behavior. Moreover, the negative effect of smoking promoting the development of ASB later in life was only evident for individuals with predisposing genetic patterns. Namely, individuals who were subjected to prenatal smoking exposure (their mothers smoked during pregnancy) demonstrated an enhanced risk of ASB only if they were genetically related to their mothers. At the same time, no link between maternal smoking and offspring’ ASB was observed if children were developed from a donated egg (van Goozen et al., 2022).
Another significant factor affecting liability to ASB in our cohort is traumatic brain injury in anamnesis, which confirms previous data (Ryan et al., 2021; Theadom et al., 2024). It was suggested that TBI can cause abnormal morphometry of the central executive network in the brain, which can result in worsening of executive functions (Ryan et al., 2021) or exacerbate other valuable triggers, including social deprivation (Guskiewicz et al., 2003), thus promoting ASB. In summary, reported findings on the higher effect of social factors on developing ASB in the Russian cohort can probably capture the effect of other genes on the occurrence of such an “environment”.
Future research should integrate various methodological approaches, including those measuring brain activity and connectivity underlying specificity of individual behavioral responses, and consider the impact of genetic and environmental factors. For instance, there is some evidence of a link between amygdala hyper-reactivity and increased impulsivity and reduced self-regulation as a response to threatening stimuli (Dotterer et al., 2017). Another study identified a link between diminished P3(P300) amplitude of electrical potential, which was obtained as a response to a visual oddball task, and manifestation of externalizing phenotypes (Iacono, 2018; Brislin et al., 2024).
The present study has several limitations. First, the set of SNPs used for PGS calculation is rather small, which can mirror the low proportion of explained variance in liability to ASB attributed to genetic impact. To be more precise, calculated PGS in the previous meta-analysis (Karlsson Linnér et al., 2021) enabled to explain 3–4 % of variance in manifesting a combined phenotype of antisocial behavior when PGS was estimated on genetic data from 579 SNPs at the genome-wide significance level.
In turn, the present study has been focused on biallelic polymorphisms only, while other structural variations in the genome, such as tandem repeats and microdeletions/duplications, which can also contribute to genetically caused manifestations of aggression, remained unstudied within the present research. Although the examined sample represents a specific cohort of individuals with a severe form of aggressive behavior (homicide), the sample size is small, which can result in type I and II errors and requires future enlargement of the examined sample. Moreover, the obtained PGS models are limited to a number of analyzed social factors while other probably relevant factors including child-parent relationship, belonging to a criminal organization, physical or sexual violence, social isolation, personality type, etc. were non-examined. Finally, since the majority of enrolled offenders were characterized by excessive alcohol/opiate use, we cannot rule out whether the reported findings are attributed to present heavy alcohol drinking.
Conclusion
In summary, the present study represents an attempt to create a prognostic model for developing antisocial behavior in a Russian cohort based on genetic data reported for European populations. Revealed findings present evidence for a higher impact of social factors rather than a composite effect of the selected “top” SNPs in predicting liability to ASB. Nevertheless, the best model was able to explain up to 21.2 % of variance in liability to ASB, especially in subjects with a family history of mental illness or criminal behavior, which was based on the genetic profile of ten SNPs and such social parameters as traumatic brain injury, severe chronic disease in anamnesis, and tobacco smoking. Future research in this field has to be focused on performing GWAS in a Russian cohort of criminal offenders (or persons with other types of antisocial behavior) to identify genetic loci and their effect estimates specific to the main ethnic groups from Russia. Obviously, such analyses will enable the design of models of liability to manifest ASB with higher prediction probabilities.
Conflict of interest
The authors declare no conflict of interest.
References
Acheson A., Vincent A.S., Cohoon A.J., Lovallo W.R. Defining the phenotype of young adults with family histories of alcohol and other substance use disorders: studies from the family health patterns project. Addict Behav. 2018;77:247-254. doi 10.1016/j.addbeh. 2017.10.014
Acland E.L., Pocuca N., Paquin S., Boivin M., Ouellet-Morin I., Andlauer T.F.M., Gouin J.P., Côté S.M., Tremblay R.E., Geoffroy M., Castellanos-Ryan N. Polygenic risk and hostile environments: links to stable and dynamic antisocial behaviors across adolescence. Dev Psychopathol. 2025;37(1):464-476. doi 10.1017/S0954579424 00004X
Antón-Galindo E., Cabana-Domínguez J., Torrico B., Corominas R., Cormand B., Fernàndez-Castillo N. The pleiotropic contribution of genes in dopaminergic and serotonergic pathways to addiction and related behavioral traits. Front Psychiatry. 2023;14:1293663. doi 10.3389/fpsyt.2023.1293663
Barnes J.C., Liu H., Motz R.T., Tanksley P.T., Kail R., Beckley A.L., Belsky D.W., Domingue B.W., Moffitt T.E., Pratt T.C., Wertz J. The propensity for aggressive behavior and lifetime incarceration risk: a test for gene-environment interaction (G × E) using whole-genome data. Aggres Violent Behav. 2019;49:101307. doi 10.1016/j.avb. 2019.07.002
Baron R.A., Richardson D.R. Human Aggression. New York: Plenum Press, 2004
Borinskaya S.A., Rubanovich A.V., Larin A.K., Kazantseva A.V., Davydova Y.D., Genetozov E.V., Khusnitdinova E.K., Yankovsky N.K. Epigenome-wide association study of CpG methylation in aggressive behavior. Russ J Genet. 2021;57(12):1454-1460. doi 10.1134/ S1022795421120048
Brislin S.J., Salvatore J.E., Meyers J.M., Kamarajan C., Plawecki M.H., Edenberg H.J., Kuperman S., … Kramer J.R., Chan G., Porjesz B.; COGA Collaborators; Dick D.M. Examining associations between genetic and neural risk for externalizing behaviors in adolescence and early adulthood. Psychol Med. 2024;54(2):267-277. doi 10.1017/S0033291723001174
Burt S.A. The genetic, environmental, and cultural forces influencing youth antisocial behavior are tightly intertwined. Annu Rev Clin Psychol. 2022;18:155-178. doi 10.1146/annurev-clinpsy-072220- 015507
Davydova Y., Kazantseva A., Enikeeva R., Mustafin R., Malykh S., Lobaskova M., Valinurov R., Akhmerova I., Khusnutdinova E. The involvement of hypothalamic-pituitary-adrenal and monoaminergic systems genes in developing aggressive behavior. Eur Neuropsychopharmacol. 2020a;31(S1):S16-S17. doi 10.1016/j.euroneuro.2019. 12.023
Davydova Y.D., Kazantseva A.V., Enikeeva R.F., Mustafin R.N., Lobaskova M.M., Malykh S.B., Gilyazova I.R., Khusnutdinova E.K. The role of oxytocin receptor (OXTR) gene polymorphisms in the development of aggressive behavior in healthy individuals. Russ J Genet. 2020b;56(9):1129-1138. doi 10.1134/S1022795420090057
Deng W.Q., Belisario K., Gray J.C., Levitt E.E., Mohammadi-Shemirani P., Singh D., Pare G., MacKillop J. Leveraging related health phenotypes for polygenic prediction of impulsive choice, impulsive action, and impulsive personality traits in 1534 European ancestry community adults. Genes Brain Behav. 2023;22(3):e12848. doi 10.1111/gbb.12848
Dotterer H.L., Hyde L.W., Swartz J.R., Hariri A.R., Williamson D.E. Amygdala reactivity predicts adolescent antisocial behavior but not callous-unemotional traits. Dev Cogn Neurosci. 2017;24:84-92. doi 10.1016/j.dcn.2017.02.008
Fritz M., Soravia S.M., Dudeck M., Malli L., Fakhoury M. Neurobiology of aggression-review of recent findings and relationship with alcohol and trauma. Biology (Basel). 2023;12(3):469. doi 10.3390/ biology12030469
Gould T.J. Epigenetic and long-term effects of nicotine on biology, behavior, and health. Pharmacol Res. 2023;192:106741. doi 10.1016/ j.phrs.2023.106741
Guskiewicz K.M., McCrea M., Marshall S.W., Cantu R.C., Randolph C., Barr W., Onate J.A., Kelly J.P. Cumulative effects associated with recurrent concussion in collegiate football players: the NCAA Concussion Study. JAMA. 2003;290(19):2549-2555. doi 10.1001/jama.290.19.2549
Heilbronner U., Papiol S., Budde M., Andlauer T.F.M., Strohmaier J., Streit F., Frank J., … Stürmer T., Müller-Myhsok B., Nöthen M.M., Rietschel M., Schulze T.G. “The Heidelberg Five” personality dimensions: genome-wide associations, polygenic risk for neuroticism, and psychopathology 20 years after assessment. Am J Med Genet B Neuropsychiatr Genet. 2021;186(2):77-89. doi 10.1002/ ajmg.b.32837
Iacono W.G. Endophenotypes in psychiatric disease: prospects and challenges. Genome Med. 2018;10(1):11. doi 10.1186/s13073-018-0526-5
Ip H.F., Van der Laan C.M., Krapohl E.M.L., Brikell I., Sánchez- Mora C., Nolte I.M., St Pourcain B., … Lundström S., Plomin R., Bartels M., Nivard M.G., Boomsma D.I. Genetic association study of childhood aggression across raters, instruments, and age. Transl Psychiatry. 2021;11(1):413. doi 10.1038/s41398-021-01480-x
Karlsson Linnér R., Biroli P., Kong E., Meddens S.F.W., Wedow R., Fontana M.A., Lebreton M., … Lee J.J., Cesarini D., Benjamin D.J., Koellinger P.D., Beauchamp J.P. Genome-wide association analyses of risk tolerance and risky behaviors in over 1 million individuals identify hundreds of loci and shared genetic influences. Nat Genet. 2019;51(2):245-257. doi 10.1038/s41588-018-0309-3
Karlsson Linnér R., Mallard T.T., Barr P.B., Sanchez-Roige S., Madole J.W., Driver M.N., Poore H.E., … Waldman I.D., Palmer A.A., Harden K.P., Koellinger P.D., Dick D.M. Multivariate analysis of 1.5 million people identifies genetic associations with traits related to self-regulation and addiction. Nat Neurosci. 2021;24(10):1367- 1376. doi 10.1038/s41593-021-00908-3
Kazantseva A.V., Kutlumbetova Yu.Yu., Malykh S.B., Lobaskova M.M., Khusnutdinova E.K. Arginine-vasopressin receptor gene (AVPR1A, AVPR1B) polymorphisms and their relation to personality traits. Russ J Genet. 2014;50(3):298-307. doi 10.1134/S10227954140 30041
Kazantseva A.V., Malykh S.B., Khusnutdinova E.K. Molecular-genetic studies of personality: from gene candidate analyses to genomewide association studies. In: Malykh S.B., Kovas Y.V., Gaysina D.A. (Eds) Genomics of Behavior: Children Development and Education. Tomsk: Tomsk State University, 2016;178-209 (in Russian)
Kazantseva A.V., Davydova Y.D., Enikeeva R.F., Valinurov R.G., Gareeva A.E., Khusnutdinova N.N., Khusnutdinova E.K. The association study of polymorphic variants of hypothalamic-pituitaryadrenal system genes (AVPR1B, OXTR) and aggressive behavior manifestation: a focus on social environment. Res Results Biomed. 2021;7(3):232-244. doi 10.18413/2658-6533-2021-7-3-0-3
Kazantseva A., Davydova Y., Enikeeva R., Mustafin R.N., Malykh S., Lobaskova M., Kanapin A., Prokopenko I., Khusnutdinova E. A combined effect of polygenic scores and environmental factors on individual differences in depression level. Genes. 2023a;14(7):1355. doi 10.3390/genes14071355
Kazantseva A.V., Davydova Y.D., Enikeeva R.F., Yakovleva D.V., Mustafin R.N., Lobaskova M.M., Malykh S.B., Khusnutdinova E.K. Individual variance in human aggression: a combined effect of polygenic score and social/lifestyle factors. Russ J Genet. 2023b;59(S2): S227-S236. doi 10.1134/s1022795423140065
Li W., Zhou H., Thygesen J.H., Heydtmann M., Smith I., Degenhardt F., Nöthen M., Morgan M.Y., Kranzler H.R., Gelernter J., Bass N., McQuillin A. Genome-wide association study of antisocial personality disorder diagnostic criteria provides evidence for shared risk factors across disorders. Psychiatr Genet. 2023;33(6):233-242. doi 10.1097/YPG.0000000000000352
Manchia M., Fanos V. Targeting aggression in severe mental illness: the predictive role of genetic, epigenetic, and metabolomic markers. Prog Neuropsychopharmacol Biol Psychiatry. 2017;77:32-41. doi 10.1016/j.pnpbp.2017.03.024
Mbatchou J., Barnard L., Backman J., Marcketta A., Kosmicki J.A., Ziyatdinov A., Benner C., … Baras A., Reid J., Abecasis G., Maxwell E., Marchini J. Computationally efficient whole-genome regression for quantitative and binary traits. Nat Genet. 2021;53(7): 1097-1103. doi 10.1038/s41588-021-00870-7
McAdams T.A., Gregory A.M., Eley T.C. Genes of experience: explaining the heritability of putative environmental variables through their association with behavioural and emotional traits. Behav Genet. 2013;43(4):314-328. doi 10.1007/s10519-013-9591-0
Musci R.J., Bettencourt A.F., Sisto D., Maher B., Masyn K., Ialongo N.S. Violence exposure in an urban city: a G×E interaction with aggressive and impulsive behaviors. J Child Psychol Psychiatry. 2019;60(1):72-81. doi 10.1111/jcpp.12966
Odintsova V.V., Hagenbeek F.A., van der Laan C.M., van de Weijer S., Boomsma D.I. Genetics and epigenetics of human aggression. Handb Clin Neurol. 2023;197:13-44. doi 10.1016/B978-0-12- 821375-9.00005-0
Pappa I., St Pourcain B., Benke K., Cavadino A., Hakulinen C., Nivard M.G., Nolte I.M., … Middeldorp C.M., Oldehinkel A.J., Pennell C.E., Boomsma D.I., Tiemeier H. A genome-wide approach to children’s aggressive behavior: the EAGLE consortium. Am J Med Genet B Neuropsychiatr Genet. 2016;171(5):562-572. doi 10.1002/ ajmg.b.32333
Pezzoli P., McCrory E.J., Viding E. Shedding light on antisocial behavior through genetically informed research. Annu Rev Psychol. 2025;76(1):797-819. doi 10.1146/annurev-psych-021524-043650
Ryan N.P., Catroppa C., Hughes N., Painter F.L., Hearps S., Beauchamp M.H., Anderson V.A. Executive function mediates the prospective association between neurostructural differences within the central executive network and anti-social behavior after childhood traumatic brain injury. J Child Psychol Psychiatry. 2021;62(9): 1150-1161. doi 10.1111/jcpp.13385
Schwartz J.A., Solomon S.J., Valgardson B.A. Socialization, selection, or both? The role of gene-environment interplay in the association between exposure to antisocial peers and delinquency. J Quant Criminol. 2019;35:1-26. doi 10.1007/s10940-017-9368-3
Tesli N., Jaholkowski P., Haukvik U.K., Jangmo A., Haram M., Rokicki J., Friestad C., Tielbeek J.J., Næss Ø., Skardhamar T., Gustavson K., Ask H., Fazel S., Tesli M., Andreassen O.A. Conduct disorder – a comprehensive exploration of comorbidity patterns, genetic and environmental risk factors. Psychiatry Res. 2024;331:115628. doi 10.1016/j.psychres.2023.115628
Theadom A., Jones K., Starkey N., Barker-Collo S., Ameratunga S., Faulkner J., Ao B.T., Feigin V. Symptoms and engagement in antisocial behavior 10 years after mild traumatic brain injury within a community civilian sample: a prospective cohort study with age-sex matched control group. Arch Phys Med Rehabil. 2024;105(2):295- 302. doi 10.1016/j.apmr.2023.07.016
Tielbeek J.J., Johansson A., Polderman T.J.C., Rautiainen M.R., Jansen P., Taylor M., Tong X., … Faraone S.V., Popma A., Medland S.E., Posthuma D.; Broad Antisocial Behavior Consortium collaborators. Genome-wide association studies of a broad spectrum of antisocial behavior. JAMA Psychiatry. 2017;74(12):1242-1250. doi 10.1001/jamapsychiatry.2017.3069
Tielbeek J.J., Uffelmann E., Williams B.S., Colodro-Conde L., Gagnon É., Mallard T.T., Levitt B.E., … Fisher S.E., Moffitt T.E., Caspi A., Polderman T.J.C., Posthuma D. Uncovering the genetic architecture of broad antisocial behavior through a genome-wide association study meta-analysis. Mol Psychiatry. 2022;27(11):4453- 4463. doi 10.1038/s41380-022-01793-3
van Goozen S.H.M., Langley K., Hobson C.W. Childhood antisocial behavior: a neurodevelopmental problem. Annu Rev Psychol. 2022; 73:353-377. doi 10.1146/annurev-psych-052621-045243
Wang S., Dan Y.L., Yang Y., Tian Y. The shared genetic etiology of antisocial behavior and psychiatric disorders: insights from pleiotropy and causality analysis. J Affect Disord. 2024;365:534-541. doi 10.1016/j.jad.2024.08.149
Watanabe K., Stringer S., Frei O., Umićević Mirkov M., de Leeuw C., Polderman T.J.C., van der Sluis S., Andreassen O.A., Neale B.M., Posthuma D. A global overview of pleiotropy and genetic architecture in complex traits. Nat Genet. 2019;51(9):1339-1348. doi 10.1038/s41588-019-0481-0
Weltens I., Bak M., Verhagen S., Vandenberk E., Domen P., van Amelsvoort T., Drukker M. Aggression on the psychiatric ward: prevalence and risk factors. A systematic review of the literature. PLoS One. 2021;16(10):e0258346. doi 10.1371/journal.pone.0258346