Telipogon rojasiae (Orchidaceae, Oncidiinae), a new species from relict forests on the western slopes of the northern Peruvian Andes

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
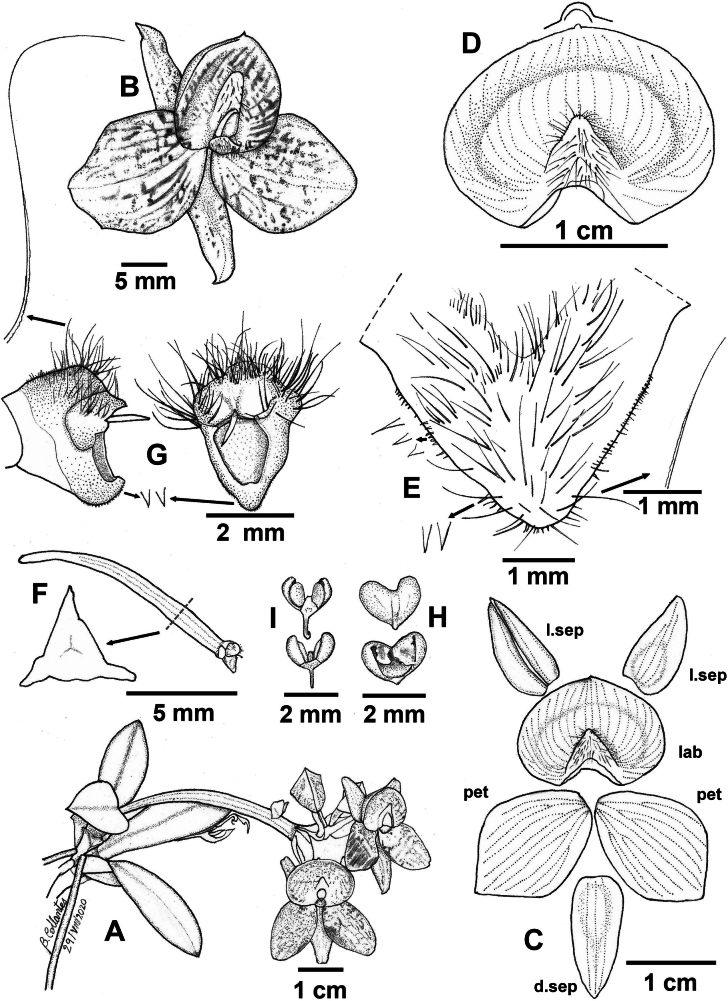 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3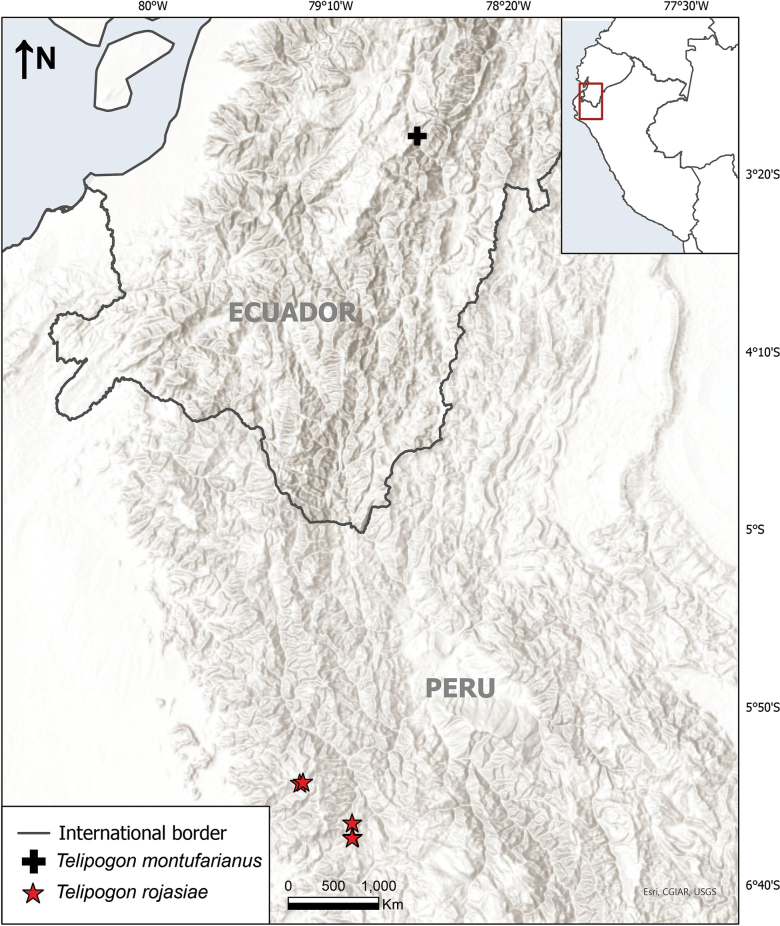 Figure 4
Figure 4 Figure 5
Figure 5|
|
|
| |
|---|---|---|---|
| Flower colour | cream-yellow with red vinaceous stains | bright yellow with brown striations | yellow-brown with brown veins |
| Petals | sub-rhombic to obovate, 10-veined | elliptic, 5-veined | elliptic, 7-veined |
| Labellum | reniform, 26-veined | reniform, 16-19-veined | broadly ovate, subcordate at the base, 19-23-veined |
| Callus | sagittate, hirsute on the sides, setose apically | widely sub-cordiform, hirsute on the sides, setose apically | sub-triangular, lingulate, puberulent |
| Column | sides of the stigmatic cavity smooth | sides of the stigmatic cavity smooth | borders of the stigmatic cavity with incurved teeth |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and animal studies · Ecology and Vegetation Dynamics Studies · Plant and Fungal Species Descriptions
Introduction
Telipogon Kunth is a Neotropical orchid genus distributed from southern Mexico and the Caribbean to Bolivia (Martel and Nauray Huari 2013). Telipogon plants grow at mid and high elevations (500–3800 m), although most species are found in mountain forests between 2200 and 3500 m. In its current circumscription, Telipogon includes 258 species (POWO 2025). However, many more remain undescribed, since Telipogon species often have highly restricted distributions (Nauray and Galán 2008; Martel 2016), and new explorations continue to reveal new taxa, with several proposed each year (e.g. Pupulin 2022; Iturralde et al. 2023; Jiménez et al. 2024). Most of this richness is found in the Andean cloud forests of Colombia, Ecuador, and Peru, where around 70% of the known Telipogon species occur (POWO 2025).
In Peru, Telipogon is currently represented by more than 50 species (Martel et al. 2020), most of which occur between 2000 and 3500 m on the eastern slopes of the Andes, characterised by year-round humidity and predominantly forested environments. Conversely, the western slopes of the Andes are dominated by the Andean shrubland ecosystem (Matorral Andino in Spanish; Ministerio del Ambiente 2019), which is characterised by a dry climate and predominance of sparse shrubland along with grassland. However, due to the Huancabamba depression – an east–west depression through the Andes between southern Ecuador and northern Peru – humid air from the east reaches some areas of the western slopes above 8° latitude south. This has made possible the occurrence of other types of ecosystems, such as the relict montane forests (“bosque relicto montano de vertiente occidental” in Spanish; Ministerio del Ambiente 2019), which are characterised by sparse humid forests. These forest remnants are biodiversity-rich ecosystems, which are unfortunately disappearing rapidly due to the expansion of human activities (Weigend et al. 2005b). Efforts to better characterise the plant diversity of this ecosystem type in Peru have taken place over the last few decades (Weigend et al. 2005a), and orchids have gained more attention during the last 10 years (e.g. Chamaya Gonzáles 2023).
As part of a survey of orchid diversity in the relict montane forests of northern Peru, one of the authors (AGDH) recorded and collected material of an unknown Telipogon species. After careful revision, we concluded that the material belonged to a previously undescribed species. Therefore, we describe and illustrate it here.
Materials and methods
Specimens of the new species were found during botanical expeditions on the western slopes of the Peruvian Andes, in the departments of Lambayeque and Cajamarca, in 2016 and 2023. Flowers were collected and preserved in spirit, whereas vegetative parts were pressed and dried and later used to prepare the line drawing and composite plate. In situ photographs were taken using a Nikon D5000 camera fitted with macro lenses of 18–55 mm and 105 mm. The collected specimens were compared with those deposited in Peruvian and Ecuadorian herbaria, as well as with descriptions and photographs from relevant literature (e.g. Dodson 1984, 2002; Medina et al. 2019). The specimens were subsequently deposited at the herbarium of the Universidad Nacional Pedro Ruiz Gallo (PRG). A preliminary conservation assessment was prepared following IUCN (2024) criteria, based on area of occupancy and extent of occurrence data, using the GeoCAT tool (Bachman et al. 2011). Descriptions of morphological characters followed the botanical terminology established by Beentje (2012).
Results
Taxonomic treatment
Telipogon
rojasiae
Taxon classificationPlantaeAsparagalesOrchidaceae
C.Martel, A.Diaz & Iturralde sp. nov.
76BB6CB2-35B1-5DDB-842D-13C67C3AD11E
urn:lsid:ipni.org:names:77370794-1
Type.
Peru • Lambayeque. Prov. Ferreñafe, [Dist. Incahuasi,] Tungula, [geographical coordinates omitted for conservation purposes], 3369 m, 29 Jan 2023, Alex G. Diaz H. 493 (holotype: PRG! [Accession N° 20392]. isotype: PRG! [Accession N° 20392]).
Diagnosis.
Telipogon rojasiae is most similar to Telipogon montufarianus but differs by its cream-yellow flowers heavily stained with red vinaceous (vs. bright yellow flowers), the sub-rhombic to obovate petals, 15–17 × 11–12 mm (vs. elliptic petals, 12 × 8 mm), the greater number of veins in the petals (9–10 veins vs. 5 veins) and labellum (26 veins vs. 16–19 veins), and its sagittate callus (vs. a widely sub-cordiform callus).
Telipogon rojasiae A. Habit; B. Flower, ¾ view; C. Dissected flower with expanded perianth (d.sep: dorsal sepal; l.sep: lateral sepal; pet: petal; lab: labellum), all in frontal view except one lateral sepal; D. Expanded labellum, frontal view with details of the apex; E. Callus, frontal view with details of the setae; F. Pedicel, ovary and column, lateral view with details of the ovary cross-section; G. Column, frontal and lateral views with details of the setae; H. Anther cap, frontal and ventral views; I. Pollinarium, frontal and ventral views.
Description.
Epiphytic herb, erect, sympodial, acaulous in young plants and with a visible short stem in older plants, up to 15 cm including the inflorescence. Roots terete, 1.6–2.2 mm in diameter. Stem abbreviated, forming pseudobulb-like structures on mature plants. Leaves 2.5–5.9 × 0.6–1.1 cm, 3–9, subcoriaceous, lanceolate-oblong to narrowly sub-elliptic, entire, acute apex, base articulated to a conduplicate leaf sheath that covers the stem; the basal leaves smaller than the upper leaves. Inflorescence an apical and lateral raceme; scape 5–7 × 0.2 cm, triquetrous, up to 5 flowers, one to two flowers opening in succession. Floral bract 15–16 × 5–6 mm (progressively decreasing in size in the following bracts), conduplicate, decurrent, falcate in natural position, widely ovate when expanded, acute, carinate. Ovary 20–21 × 3.0–3.1 mm, triquetrous, winged, pedicellate. Pedicel 6–11 mm long, triquetrous. Flowers 20–25 mm in diameter, non-resupinate. Sepals greenish-yellow with red vinaceous, spots, semitranslucent; lateral sepals 11–12 × 5–6 mm, ovate, entire, basally concave, acute apex, curved in natural position, with entire margin, 3-veined, carinate abaxially; dorsal sepal 13–14 × 5–6 mm, narrowly ovate, entire, concave towards the base, acute apex, 3-veined, carinate abaxially. Petals 15–17 × 11–12 mm, red-vinaceous with spots, sub-rhombic to obovate, apex acute to sub-acute, 9–10-veined. Labellum 12–13 × 16–17 mm when expanded, red-vinaceous with spots, reniform, concave, the base embracing almost the entire column, the apex rounded, protruding, 26-veined. Callus 3.2–3.6 × 5.7–6.1 mm, elevated, dark purple towards the apex, yellow towards the base, sagittate with a rounded apex, setose at the centre of the apex, fimbriate margin. Column 3.4 mm long, 2.4 mm wide, sub-rhomboid, dark yellow, ventral surface densely covered by minute conical papillae with acute apex, 3 tufts of setae; setae dark purple to white, simple, capilliform, up to 2.2 mm long. Clinandrium concave, with rounded edges, with a dorsal projection covering up to half of the anther cap. Stigma suborbicular. Rostellum erect. Anther cap 1.7 × 2.1 mm, red, cordiform, bilocular. Pollinarium 2.4 × 1.9 mm; pollinia 4 in 2 unequal pairs; outer pair obovoid, convex-flat; inner pair ellipsoid, laterally complanate; caudicle hyaline; viscidium uncinate. Fruit not seen.
Telipogon rojasiae A. Habit; B. Floral bract, lateral view; C. Flower, frontal view; D. Dissected flower, frontal view with details of the abaxial side of the sepals and the labellum surface; E. Callus and column, frontal view; F. Pedicel, ovary and column, lateral view with details of the ovary cross-section; G. Column, frontal, top and lateral views; H. Anther cap and pollinarium, ventral and dorsal views. Note that all the floral segments are in their natural position, not expanded.
Telipogon rojasiae. Holotype. GPS coordinates hidden for conservation purposes.
Phenology.
Plants of T. rojasiae have been recorded in flower between August and January, the dry season in the southern hemisphere.
Eponymy.
Telipogon rojasiae is named after Dr Consuelo Rojas, professor of Botany at the Universidad Nacional Pedro Ruiz Gallo, for her work on biodiversity and plant conservation in Northern Peru.
Distribution, habitat and ecology.
Telipogon rojasiae is only known from the western slopes of the northern Peruvian Andes between 2800 and 3400 m asl (Fig. 4). The ecosystem of this area has been categorised as relict montane forests of the western slope (Ministerio del Ambiente 2019). These relict forests are located in the western Andean slopes of northwest Peru and southwest Ecuador and possess a rich biodiversity, including many endemic species and genera (Weigend et al. 2005a, 2005b).
Distribution map of Telipogon rojasiae and the morphologically similar T. montufarianus.
We have so far recorded five small subpopulations, which occur in small forest patches. In these areas, Miconia sp. is the dominant plant species, followed by Brachyotum ledifolium (Desr.) Triana, Hedyosmum scabrum (Ruiz & Pav.) Solms, Grosvenoria jelskii (Hieron.) R.M.King & H.Rob., and Myrica pubescens Humb. & Bonpl. ex Willd. In terms of orchids, many species co-occur with T. rojasiae, including Cyrtochilum loesenerianum (Schltr.) Dalström, Epidendrum capitellatum C.Schweinf., E. hemiscleria Rchb.f., E. scutella Lindl., Fernandezia ionanthera (Rchb.f. & Warsz.) Schltr., Liparis elegantula Kraenzl., and Telipogon papilio Rchb.f. & Warsz.
The flower morphology resembles the general morphology of flowers in Telipogon species pollinated by sexual deception involving tachinid male flies (e.g. T. peruvianus T.Hashim., T. bowmanii Rchb.f.; Martel et al. 2016, 2019), which suggests T. rojasiae may have a similar pollination mechanism.
Preliminary conservation status.
Telipogon rojasiae was first observed in 2016; since then, only five small subpopulations, each comprising fewer than 20 individuals, have been recorded. The five subpopulations occur in disturbed and patchy forests, which are threatened by deforestation. Its EOO is estimated to be 132.5 km^2^, falling into the Endangered category under subcriterion B1, whereas its AOO is estimated at 5 km^2^ (a cell of 1 km^2^ due to the small forest remnants), falling into the Critically Endangered category under subcriterion B2. The known habitat of T. rojasiae, the relict forests, is severely fragmented (a), and we project that the five known subpopulations will decline over time and the AOO and EOO will be reduced. Therefore, following the IUCN (2024) criteria, we informally assessed the conservation status of T. rojasiae as Critically Endangered [CR B2ab(i,ii,iii,iv)].
Taxonomic notes.
Telipogon rojasiae is morphologically most similar to T. montufarianus H.Medina, J.Portilla & Hirtz, for being a medium-sized plant (< 10–15 cm including the inflorescence) and having small flowers of similar size (20–25 mm in diam.), a similar colouration in the sepals, the concave labellum, an elevated callus, and a column with three tufts of capilliform setae (Fig. 5). Some of the differences between both species have been detailed in the diagnosis; however, the most notable are the colour of the flowers (cream-yellow flowers heavily stained with red vinaceous in T. rojasiae vs. bright yellow flowers in T. montufarianus), the shape of the petals (sub-rhombic to obovate petals vs. elliptic), the shape of the callus (sagittate in T. rojasiae vs. widely sub-cordiform in T. montufarianus), the number of veins in the petals (10 veins in T. rojasiae vs. 5 veins in T. montufarianus) and the labellum (26 veins in T. rojasiae vs. 16–19 veins in T. montufarianus) (Fig. 4). Additionally, T. rojasiae inhabits the montane forests of the eastern Andean slopes of northwestern Peru, while T. montufarianus is distributed in the western slopes of southeastern Ecuador (Table 1). Telipogon rojasiae seems to be also morphologically similar to Telipogon frymirei Dodson, from southeastern Ecuador, as they both have a concave labellum and an elevated callus; however, T. rojasiae can be easily discriminated by the sub-rhombic to sub-obovate petals (vs. elliptic petals in T. frymirei), the reniform labellum (vs. broadly ovate in T. frymirei), the sagittate callus (vs. sub-triangular, lingulate callus in T. frymirei), and the rounded margins of the stigmatic cavity (vs. a stigmatic cavity with incurved teeth in T. frymirei) (Table 1).
Comparison between Telipogon rojasiae and T. montufarianus; A. Plant of T. rojasiae; B. Plant of T. montufarianus; C. Flower of T. rojasiae, frontal view; D. Flower of T. montufarianus, frontal view.
Table 1.: Summary of principal differences between T. rojasiae and morphologically similar species.
There are at least two Telipogon species that might be closely related to T. rojasiae: T. montufarianus and an undescribed Telipogon species presented as T. frymirei Dodson by Dodson (2002) in his book Native Ecuadorian Orchids, volume III. The latter displays the typical concave labellum and a hairy elevated callus, as T. rojasiae and T. montufarianus do; in addition, it has dark red to purplish flowers. These and other characters of the specimen portrayed by Dodson (2002) differ strongly from the characters presented in the original description and drawing of T. frymirei (Dodson 1984). The misidentified species resembles T. montufarianus in petal shape and T. rojasiae in labellum shape. Unfortunately, no material is available to compare our new entity with the actual and misidentified T. frymirei. Moreover, in the same book, Dodson depicted a picture of a flower of T. montufarianus – which was still undescribed at that time – as T. ecuadorensis Schltr., despite these two species being morphologically very distinct. Explorations in the type locality of T. frymirei are being carried out, which we hope will shed light on the true identity of this and other Telipogon species, as well as the potential occurrence of T. rojasiae in Ecuador.
As a side note, T. montufarianus was originally described as T. montufariana (Medina et al. 2019), which is an incorrect declension, since the genus name Telipogon is masculine, and therefore the species epithet should be declined with the suffix “ianus”. Similarly, T. humboldtianus H.Medina, J.Portilla & Hirtz was originally named T. humboldtiana (Medina et al. 2019), which we correct here.
Other records.
Peru • Cajamarca. Prov. Chota, [Dist. Querocoto,] Querocoto, 3144 m, 20 Aug 2017, Alex G. Diaz H. 271 (PRG! [Accession N° 20391]); • ibid. 3148 m, 20 Sep 2018, Alex G. Diaz H. 270 (PRG! [Accession N° 20390]); • ibid. Pagaibamba, 2808 m, 11 Aug 2018, Alex G. Diaz H. 268 (PRG! [Accession N° 20389]); • ibid. Lambayeque. Prov. Ferreñafe, [Dist. Incahuasi,] Tungula, 3400 m, 17 Oct 2016, Alex G. Diaz H. 128 (PRG! [Accession N° 18754]).
Supplementary Material
XML Treatment for Telipogon rojasiae
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bachman S Moat J Hill Ade la Torre J Scott B (2011) Supporting Red List threat assessments with Geo CAT: Geospatial conservation assessment tool.Zoo Keys 150: 117–126. 10.3897/zookeys.150.2109 PMC 323443422207809 · doi ↗ · pubmed ↗
- 2Beentje H (2012) The Kew Plant Glossary: An Illustrated Dictionary of Plant Terms. Royal Botanic Gardens, Kew.Kew Publishing, Richmond, UK, 160 pp.
- 3Chamaya Gonzáles JA (2023) Diversidad de especies de la familia Orchidaceae en el bosque La Palma – Chota – Cajamarca. M Sc thesis. Universidad Nacional de Cajamarca, Peru.
- 4Dodson CH (1984) Telipogon frymirei. In: Dodson CH, Dodson PM (Eds) Icones Plantarum Tropicarum, Fascicle 10. The Marie Selby Botanical Gardens, Sarasota, Plate 991.
- 5Dodson CH (2002) Native Ecuadorian Orchids. Volume III: Lepanthopsis-Oliveriana.Dodson Trust, Sarasota, 231 pp.
- 6Iturralde GA Monteros MF Jiménez MM Martel C Baquero LE (2023) Telipogon pillaropatatensis (Oncidiinae): A new species from the east-central Andes of Ecuador.Lankesteriana 23: 495–509. 10.15517/lank.v 23i 3.58100 · doi ↗
- 7IUCN (2024) IUCN Standards and Petitions Committee 2024. Guidelines for Using the IUCN Red List Categories and Criteria. Version 16. Prepared by the Standards and Petitions Committee. https://www.iucnredlist.org/resources/redlistguidelines.pdf [Accessed 01 July 2025]
- 8Jiménez MM Martel C Garzón-Suárez HX Baquero LE Mashendo V Iturralde GA (2024) Telipogon leisberthvelezii (Orchidaceae: Oncidiinae), a new orchid species from the Cordillera del Cóndor in Ecuador.Kew Bulletin 79: 97–106. 10.1007/s 12225-023-10145-5 · doi ↗