Morphological and molecular identification of new species and records of Daldinia (Hypoxylaceae, Xylariales) from Guizhou Province, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
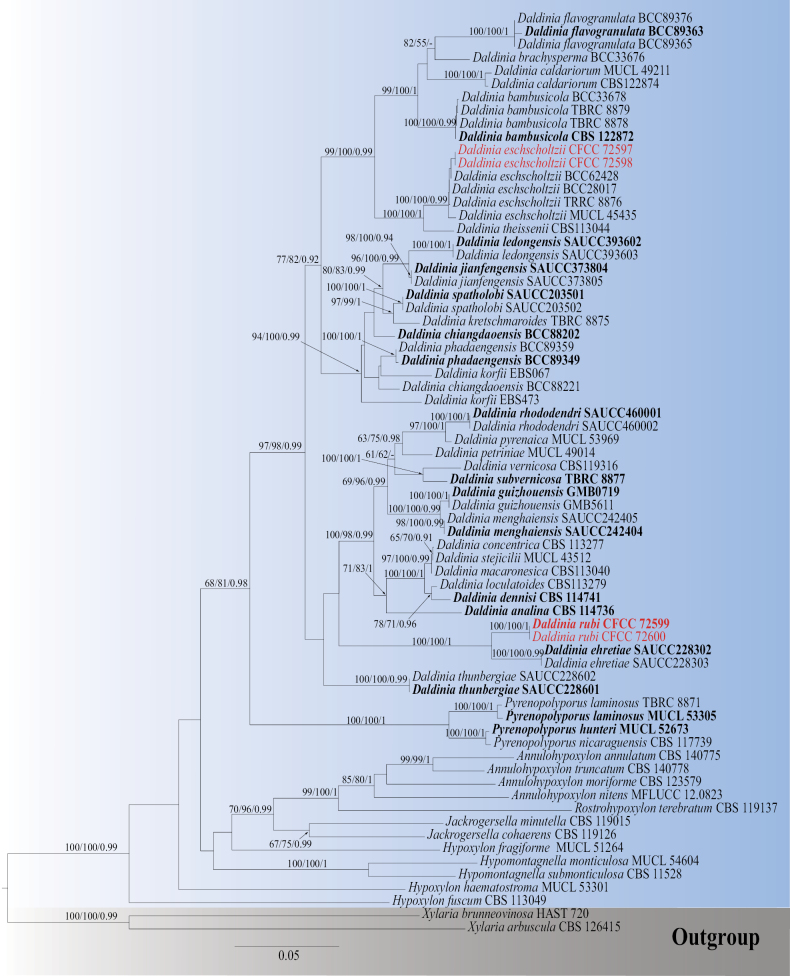 Figure 1
Figure 1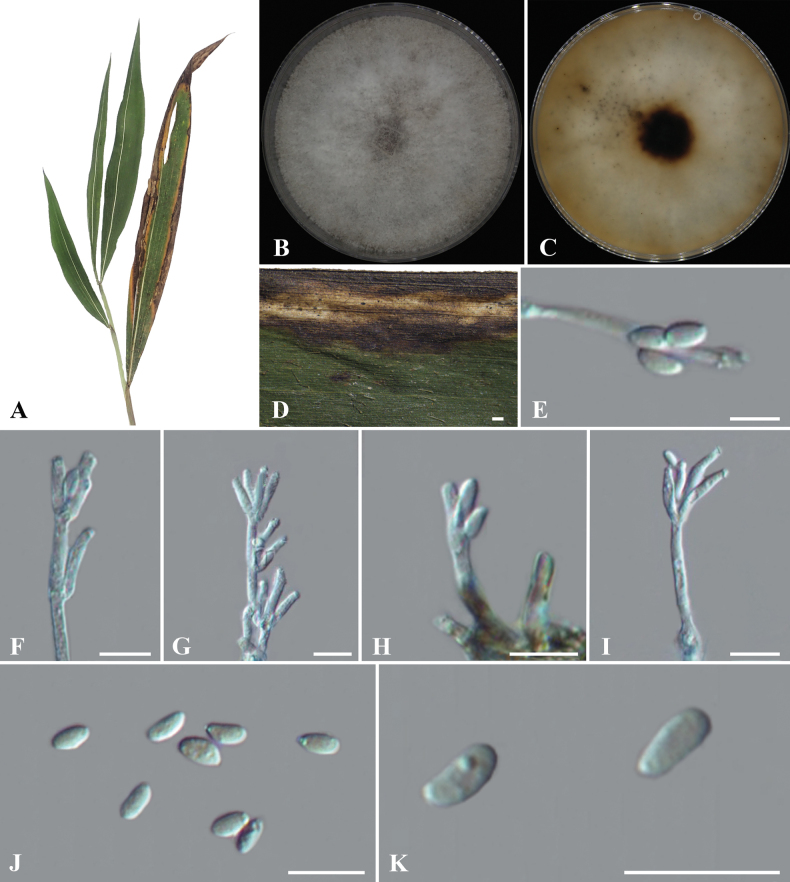 Figure 2
Figure 2 Figure 3
Figure 3| Locus | PCR primers | PCR: thermal cycles (annealing temp. in bold) | References |
|---|---|---|---|
| ITS | ITS1/ITS4 | (95 °C: 30 s, 51 °C: 30 s, 72 °C: 1 min) × 35 cycles |
|
| 28S | LROR/LR5 | (95 °C: 45 s, 55 °C: 30 s, 72 °C: 1 min) × 35 cycles |
|
|
| fRPB2-5F/fRPB2-7cR | (95 °C: 15 s, 55 °C: 30 s, 72 °C: 1 min) × 35 cycles |
|
|
| T1/T22 | (95 °C: 35 s, 52 °C: 55 s, 72 °C: 2 min) × 35 cycles |
|
| Species | Strains | Country | GenBank Accession Numbers | |||
|---|---|---|---|---|---|---|
| ITS | 28S | rpb2 | tub2 | |||
|
| CBS 140775 | USA |
|
|
|
|
|
| CBS 123579 | France |
|
|
|
|
|
| MFLUCC 12.0823 | Thailand |
|
|
|
|
|
| CBS 140778 | USA |
|
|
|
|
|
| CBS 114736T | Ecuador |
|
|
|
|
|
| CBS 122872T | Thailand |
|
|
|
|
|
| TBRC 8878 | Thailand |
|
|
|
|
|
| TBRC 8879 | Thailand |
|
|
|
|
|
| Thailand |
|
|
|
| |
|
| Thailand |
|
|
|
| |
|
| MUCL 49211 | France |
|
|
|
|
|
| CBS 122874 | USA |
|
|
|
|
|
| Thailand |
|
|
|
| |
|
| Thailand |
|
|
|
| |
|
| CBS 113277 | Germany |
|
|
|
|
|
| CBS 114741T | Australia |
|
|
|
|
|
| SAUCC228302T | China |
|
|
|
|
|
| SAUCC228303 | China |
|
|
|
|
|
| China |
|
|
|
| |
|
| China |
|
|
|
| |
|
| MUCL 45435 | Benin |
|
|
|
|
|
| TRRC 8876 | Thailand |
|
|
|
|
|
| Thailand |
|
|
|
| |
|
| Thailand |
|
|
|
| |
|
| Thailand |
|
|
|
| |
|
| Thailand |
|
|
|
| |
|
| Thailand |
|
|
|
| |
|
| GMB0719T | China |
|
|
|
|
|
| GMB5611 | China |
|
|
|
|
|
| SAUCC373804T | China |
|
|
|
|
|
| SAUCC373805 | China |
|
|
|
|
|
| EBS067 | Argentina |
|
|
|
|
|
| EBS473 | Argentina |
|
|
|
|
|
| TBRC 8875 | Thailand |
|
|
|
|
|
| SAUCC393602T | China |
|
|
|
|
|
| SAUCC393603 | China |
|
|
|
|
|
| CBS 113279 | UK |
|
|
|
|
|
| CBS 113040 | Spain |
|
|
|
|
|
| SAUCC242404T | China |
|
|
|
|
|
| SAUCC242405 | China |
|
|
|
|
|
| MUCL 49014 | Austria |
|
|
|
|
|
| Thailand |
|
|
|
| |
|
| Thailand |
|
|
|
| |
|
| MUCL 53969 | France |
|
|
|
|
|
| SAUCC460001T | China |
|
|
|
|
|
| SAUCC460002 | China |
|
|
|
|
|
| China |
|
|
|
| |
|
| China |
|
|
|
| |
|
| SAUCC203501T | China |
|
|
|
|
|
| SAUCC203502 | China |
|
|
|
|
|
| MUCL 43512 | Papua New |
|
|
|
|
|
| TBRC 8877T | Thailand |
|
|
|
|
|
| CBS 113044 | Argentina |
|
|
|
|
|
| SAUCC228601T | China |
|
|
|
|
|
| SAUCC228602 | China |
|
|
|
|
|
| CBS 119316 | Germany |
|
|
|
|
|
| MUCL 54604 | French Guiana |
|
|
|
|
|
| CBS 115280 | France |
|
|
|
|
|
| MUCL 51264 | Germany |
|
|
|
|
|
| CBS 113049 | France |
|
|
|
|
|
| MUCL 53301 | Martinique |
|
|
|
|
|
| CBS 119126 | Germany |
|
|
|
|
|
| CBS 119015 | Portugal |
|
|
|
|
|
| MUCL 52673T | Ivory Coast |
|
|
|
|
|
| TBRC 8871 | Thailand |
|
|
|
|
|
| MUCL 53305T | Martinique |
|
|
|
|
|
| CBS 117739T | Burkina Faso |
|
|
|
|
|
| CBS 119137T | Thailand |
|
|
|
|
|
| CBS 126415 | Germany |
|
|
|
|
|
| HAST 720 | Martinique |
|
|
|
|
| Species | Conidiophores (Branching/Conidiogenous Cells/Size) | Conidiogenous Cells (Shape/Size) | Conidia (Shape/Size) | References |
|---|---|---|---|---|
|
| Dichotomously or trichotomously branched, 2–3 conidiogenous cells per terminus, 110–160 × 2.1–2.7 μm | Cylindrical, 10.1–15.3 × 2.5–3.1 μm | Subglobose or ellipsoid, 3.4–4.5 × 2.5–3.1 μm |
|
|
| Dichotomously or trichotomously branched, 2–3 conidiogenous cells per terminus, 150–220 × 2.5–3 μm | Clavate (apically enlarged), 14.9–32.7 × 2.8–4.4 μm | Subglobose or ellipsoid, 5.8–9.1 × 4.1–5.8 μm |
|
|
| Mononematous or dichotomously branched, 1–2 conidiogenous cells per terminus, 100–210 × 3.1–4.3 μm | Cylindrical, 16.8–24.5 × 2.7–4.1 μm | Ellipsoid or cylindrical, 4.2–6.6 × 1.7–2.8 μm |
|
|
| Mononematous, dichotomously or trichotomously branched, 2–3 conidiogenous cells per terminus, 120–214 × 2.3–4.1 μm | Cylindrical, 14.9–22.7 × 2.1–3.6 μm | Ellipsoid or dacryoid, 4.9–6.8 × 2.3–3.5 μm |
|
|
| Mononematous, dichotomously or trichotomously branched, 1–4 conidiogenous cells per terminus, 70–120 × 2.9–4.4 μm | Cylindrical, 12.1–16.9 × 2.6–3.6 μm | Subglobose or ellipsoid, 3.2–5.5 × 2.6–3.7 μm |
|
|
| Rarely mononematous or dichotomously branched, 1 conidiogenous cell per terminus, 120–200 × 1.7–2.1 μm | Clavate, 8.6–15.1 × 1.2–3.4 μm | Ellipsoid or fusiform, 3.2–4.0 × 1.4–2.0 μm |
|
|
| Dichotomously or trichotomously branched (occasionally), 1–2 conidiogenous cells per terminus, 80–150 × 1.9–3.4 μm | Cylindrical or clavate, 16.9–23.5 × 2.0–3.5 μm | Ellipsoid, subglobose or dacryoid, 4.7–8.2 × 3.1–4.0 μm |
|
|
| Rarely mononematous or dichotomously branched, conidiogenous cell number unclear, 40–90 × 1.4–2.0 μm | Cylindrical or ampulliform, 5.9–11.6 × 1.1–2.9 μm | Ellipsoid, cylindrical or banana - shaped, 3.2–5.1 × 1.1–2.3 μm |
|
|
| Mononematous or dichotomously branched, 2–3 conidiogenous cells per terminus, 16–36 × 2.5–4 μm | Cylindrical or laterally cylindrical, (11.5–)13–20.5 × 2–3.5 μm | Ellipsoid to dacryoid, 4.5–8 × 3–4.5 μm | This study |
|
| Mononematous, dichotomously or trichotomously branched, 1–4 conidiogenous cells per terminus, 70–220 × 1.8–4.1 μm | Cylindrical or clavate, 6.7–17.3 × 1.9–2.5 μm | Ellipsoid or teardrop - shaped, 3.2–5.0 × 2.2–3.1 μm |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Mycorrhizal Fungi and Plant Interactions · Yeasts and Rust Fungi Studies
Introduction
Daldinia was originally introduced by Cesati and De Notaris (Cesati and De Notaris 1863). In the early 20^th^ century, Child (1932) conducted the first comprehensive global study on Daldinia, describing 13 species and introducing anamorphic features and physiological traits for classification purposes. However, her work contained inaccuracies in species descriptions, leading to confusion. Later, Ju et al. (1997) conducted the second global revision, clarifying some errors and providing detailed descriptions of D. bakeri, D. caldariorum, and D. loculata. Rogers et al. (1999) redefined the type species D. concentrica and held that it was actually D. childiae. In the early 21^st^ century, the integration of molecular biology and chemotaxonomic techniques significantly advanced the taxonomic study of Daldinia. Johannesson et al. (2000) and Stadler et al. (2001a) constructed preliminary phylogenetic relationships using nuc rDNA ITS sequence analysis. The application of HPLC (high-performance liquid chromatography) and scanning electron microscopy (SEM) enabled the examination of chemical profiles and ultrastructures of historical specimens, revealing the diversity of secondary metabolites and their taxonomic significance in Daldinia (Stadler et al. 2001b, c, 2002; Bitzer et al. 2007). Subsequent ecological studies uncovered its endophytic lifestyle and rapid colonization ability following host damage (Rogers 2000; Stadler 2011). Stadler et al. (2014) integrated morphological, molecular phylogenetic, and chemotaxonomic approaches, analyzing data from thousands of specimens and cultures to elucidate the complex diversity of secondary metabolites and phylogenetic relationships in Daldinia. Overall, the taxonomy of Daldinia has evolved from a morphology-based approach to a multidisciplinary research framework, gradually resolving earlier taxonomic confusions and providing a foundation for understanding its ecological and evolutionary relationships.
In recent years, the classification framework of Daldinia has stabilized. Wendt et al. (2018) confirmed through multi-gene phylogenetic analysis (ITS, 28S, rpb2, and tub2) that Daldinia and its relatives form a distinct clade within Hypoxylaceae, clearly differentiated from Hypoxylon and Pyrenopolyporus. The application of a polyphasic approach has significantly increased species diversity, revealing multiple cryptic species. For instance, Stadler et al. (2004) described five new species from the Canary Islands, Channel Islands, and Sicily, while Wongkanoun et al. (2019) reported a new record and a new species from northern Thailand. Additionally, Pažoutová et al. (2013) identified D. hawksworthii, an insect-associated endophytic species, based on molecular data and conidial structures. Yin et al. (2024) isolated and identified seven new anamorphic species from diseased leaves in southern China, analyzing their geographical distribution and host specificity, which suggests that warm, humid, and vegetatively rich regions may harbor more Daldinia and fungal resources.
The stromata of Daldinia fungi are prominent and persistent, often forming dense clusters on woody plants, making them easily noticeable. Internally, the stromata exhibit horizontal zonation, a key feature distinguishing this genus from other pyrenomycete fungi (Stadler et al. 2014). Traditionally, Daldinia was considered a saprophyte, primarily causing white rot on dead angiosperm wood. However, recent studies suggest they may persist as endophytes within host tissues, forming stromata only when the host is damaged or stressed, leading to their previous misidentification as “rare” despite potentially widespread distribution (Stadler and Hellwig 2005). In this paper, during the identification of plant pathogenic fungi from diseased leaves in Guizhou Province, China, two Daldinia species were unexpectedly isolated from Indocalamus hirsutissimus and Rubus idaeus. Molecular and morphological analyses confirmed the establishment of a new species, D. rubi sp. nov., and reported a new host record species, D. eschscholtzii. This study provides detailed descriptions, illustrations, and DNA-based phylogenetic analyses to validate their taxonomic classification.
Materials and methods
Sample collection and fungal isolation
Diseased leaves of Indocalamus hirsutissimus and Rubus idaeus were sampled from Guizhou, China, and sampling information was recorded (Rathnayaka et al. 2025). For sampling positions on the leaf, the junction of diseased and healthy tissue was targeted. The diseased leaf tissues were cut into small pieces of approximately 5 mm × 5 mm using a sterile scalpel. These tissue pieces were then subjected to surface sterilization (rinsed in 75% ethanol for 30 s, followed by 2% sodium hypochlorite for 2 min, and finally rinsed three times in sterile distilled water). The surface-sterilized tissues were plated onto potato dextrose agar (PDA), which consisted of 20% diced potatoes, 2% agar, and 2% glucose. Petri dishes were incubated at 25 °C in the dark for 2–3 days. After colony formation, hyphal tips were carefully picked under a stereomicroscope and transferred to fresh PDA plates to obtain pure cultures, following the method described by Crous et al. (2019a). Type specimens of the new species were deposited in the Museum of Beijing Forestry University (BJFC), and ex-type living cultures were preserved at the China Forestry Culture Collection Center (CFCC), Beijing, China.
Morphological observation
Cultures were grown on PDA at 25 °C under a 12 h light/dark cycle (Crous et al. 2019b). After 14 days, colony measurements were taken, and characteristics such as color, shape, and aerial mycelium density were observed and recorded. Slide mounts were prepared in water from sporulating colonies on PDA. Observations were performed using a LEICA DM 2500 dissecting microscope (Wetzlar, Germany) and a NIKON ECLIPSE 80i compound microscope with differential interference contrast (DIC) illumination. Images were captured using a NIS DS-RI2 camera with Nikon NIS-Elements F4.30.01 software. Conidial length was measured from the base of the basal cell to the base of the apical appendage, and conidial width was measured at the widest point. A random selection of 50 conidia was used for measurements. Taxonomic novelties were deposited in MycoBank (Crous et al. 2004).
DNA extraction, PCR amplification, and sequencing
When mycelia spread fully on PDA, genomic DNA was extracted using the cetyltrimethylammonium bromide (CTAB) method. PCR primers (forward and reverse) and amplification conditions are detailed in Table 1. PCR amplification was performed on a BIO-RAD PTC-200 thermal cycler. Each 20 μL reaction system contained 10 μL of Master Mix (Promega Corporation), 7 μL of double-deionized water, 1 μL each of forward and reverse primers, and 1 μL of DNA template. PCR products were analyzed by 2% agarose gel electrophoresis and sent to Tsingke Biotechnology Co., Ltd. (Beijing, China) for sequencing.
Phylogenetic analyses
The sequences obtained were assembled using SEQMAN software, and reference sequences from related publications (Yin et al. 2024; Liu et al. 2025) were retrieved from the National Center for Biotechnology Information (NCBI; https://www.ncbi.nlm.nih.gov). All sequences generated in this study were submitted to GenBank (Table 2). Sequences were aligned in MAFFT on the web server (https://mafft.cbrc.jp/alignment/server/) (Katoh and Standley 2013; Katoh et al. 2019), and further adjustments and editing were made with MEGA (Tamura et al. 2013). The multigene sequence alignments and the resulting trees were deposited in TreeBASE (https://treebase.org; study ID S32158). Maximum parsimony (MP), maximum likelihood (ML), and Bayesian inference (BI) were selected to construct phylogenetic trees using PAUP, PHYML, and MRBAYES (Huelsenbeck and Ronquist 2001; Swofford 2003; Silvestro and Michalak 2012). Phylograms were visualized with FIGTREE (http://tree.bio.ed.ac.uk/software/figtree/) and further edited with ADOBE ILLUSTRATOR CS (Adobe Systems Inc., USA). Maximum-parsimony bootstrap values (MPBP) and maximum-likelihood bootstrap values (MLBP) ≥ 50% and Bayesian posterior probabilities (BYPP) ≥ 0.90 were shown on the tree.
Maximum parsimony (MP) analysis used the tree bisection and reconnection (TBR) branch-swapping algorithm with 1,000 random-addition sequences in a heuristic search (Swofford 2003). The maximum number of trees was set at 5,000 branches of zero length, and all parsimonious trees were saved. Tree length (TL), consistency index (CI), retention index (RI), and rescaled consistency index (RC) were calculated (Swofford 2003). For maximum likelihood (ML) analysis, the GTR GAMMA model of site substitution was applied, estimating gamma-distributed rate heterogeneity and the proportion of invariant sites (Guindon et al. 2010). Branch support in MP and ML was evaluated using a 1,000-replicate bootstrap (BS) method (Hillis and Bull 1993). Bayesian inference (BI) analysis, using a Markov chain Monte Carlo (MCMC) algorithm, was employed to calculate Bayesian posterior probabilities (Rannala and Yang 1996). MRMODELTEST (Posada and Crandall 1998) was used to estimate the nucleotide substitution model for weighted Bayesian analysis. Two MCMC chains, starting from random trees, were run for 1,000,000 generations until the average standard deviation of split frequencies dropped below 0.01, sampling trees every 100^th^ generation. The first 25% of trees were discarded as burn-in, and Bayesian posterior probabilities (BPP) were calculated from the remaining 7,500 trees.
Result
Phylogenetic analyses
The BLAST results indicated that the four isolates belong to Daldinia. In this genus, the combined ITS, 28S, rpb2, and tub2 dataset consisted of 3,145 characters, including alignment gaps (408 for ITS, 801 for 28S, 830 for rpb2, and 1,106 for tub2), of which 2,006 were constant and 228 were variable but parsimony-uninformative characters. MP analysis with the remaining 911 parsimony-informative characters resulted in one equally parsimonious tree: tree length (TL) = 4,936; consistency index (CI) = 0.369; retention index (RI) = 0.682; and rescaled consistency index (RC) = 0.251. In ML analysis based on the combined gene dataset, the matrix had 239 distinct alignment patterns. Estimated base frequencies were as follows: A = 0.238203, C = 0.267126, G = 0.257477, T = 0.237194, AC = 0.732392, AG = 5.679375, AT = 0.806583, CG = 0.787827, CT = 4.670947, GT = 1.000000, gamma distribution shape parameter α = 0.178803, and likelihood value ln = −26014.402666. The phylogenetic analysis revealed that the four isolates (CFCC 72597, CFCC 72598, CFCC 72599, CFCC 72600) were categorized into two clades, representing one new species, D. rubi, and one known species, D. eschscholtzii (Fig. 1). The single-gene trees for ITS, 28S, and tub2 of Daldinia are shown in Suppl. material 1.
Phylogram generated from RAxML analysis based on ITS, 28S, rpb2, and tub2 sequence data of Daldinia isolates. The tree was rooted with Xylaria brunneovinosa (HAST 720) and X. arbuscula (CBS 126415). MP, ML (≥ 50%), and BI (≥ 0.9) bootstrap supports are shown near the nodes. Isolates from this study are marked in red, and ex-type strains are marked in bold.
Taxonomy
Daldinia
eschscholtzii
Taxon classificationFungiXylarialesHypoxylaceae
(Ehrenb.: Fr.) Rehm, Annls mycol. 2(2): 175. 1904.
06AB00A0-0394-5B4C-A691-DA9707D1738B
858704
Description.
Sexual morph: not observed. Asexual morph: Conidiophores with Virgariella-like to Nodulisporium-like branching, mononematous or dichotomous, bearing 1–4 conidiogenous cells per terminus, smooth to finely roughened, hyaline, aseptate, 23.5–28.5 × 2.5–3 (av. ± S.D.= 26 ± 1.9 × 3 ± 0.2 µm, n = 30) μm. Conidiogenous cells terminal or lateral, cylindrical to phialidic, smooth-finely roughened, hyaline, aseptate, apical, 9–14.5 × 2–3 μm (av. ± S.D.= 11.5 ± 1.5 × 2.5 ± 0.3 µm, n = 30). Conidia ellipsoid, cylindrical, oval in shape, smooth, hyaline to pale yellow, aseptate, solitary, holoblastic-sympodial, 4.5–6.5 × 2–4 μm (av. ± S.D.= 5.5 ± 0.5 × 3.0 ± 0.4 µm, n = 50).
Culture characters.
Colonies were dense and uniform, with entire margins and velvety texture, fluffy, surface grayish-white, slightly darker centrally, reverse pale yellow to buff, dark brown at the center. Colonies developed abundant aerial hyphae and reached 60 mm in diameter after 7 days on PDA at 25 °C.
Daldinia eschscholtzii (BJFC-S2541) A, D. Leaf of host Rubus idaeus; B. Front colony morphology on PDA at 14 days; C. Reverse colony morphology on PDA at 14 days; E–I. Conidiogenous cells and conidia; J, K. Conidia. Scale Bars: 500 μm (D); 10 μm (E–K).
Materials examined.
China • Guizhou Province, Xingyi City, Maling River Canyon Scenic Area, 25°7'49"N, 104°57'19"E, on the leaf spots of Indocalamus hirsutissimus, 26 Jul 2024, C.M. Tian, N. Jiang, S.J. Li and Y.Y. Wu, BJFC-S2541, living culture CFCC 72597; *ibid.*BJFC-S2542, living culture CFCC 72598.
Notes.
Based on multi-locus phylogenetic analysis, two strains (CFCC 72597 and CFCC 72598) formed a highly supported clade with Daldinia eschscholtzii (100% MP/100% ML/0.99 BYPP). D. eschscholtzii is a multifunctional wood-inhabiting fungus exhibiting endophytic, saprophytic, and pathogenic traits (Stadler et al. 2014). Daldinia eschscholtzii exhibits endophytic, saprophytic, and pathogenic traits (Stadler et al. 2014) and has a broad host range spanning 56 plant families across 31 countries (Suppl. material 2), mainly colonizing decaying dicotyledonous wood and occasionally occurring on marine algae (Karnchanatat et al. 2007; Zhang et al. 2008; Tarman et al. 2012). Its human pathogenic potential is also confirmed (Ng et al. 2012, 2016; Yew et al. 2014; Chan et al. 2015). This study represents the first documented record of D. eschscholtzii on Indocalamus, supported by phylogenetic congruence with known D. eschscholtzii and alignment with its generalist ecology of colonizing lignocellulosic substrates, like Indocalamus.
Daldinia
rubi
Taxon classificationFungiXylarialesHypoxylaceae
Y.Y. Wu & C.M. Tian sp. nov.
744209AA-4038-5ED0-B724-3F97FA17E91A
858703
Type.
China • Guizhou Province, Guiyang City, Yunyan District, Qianlingshan Forest Park, 26°36'06"N, 106°41'42"E, on the leaf spots of Rubus idaeus, 26 Jul 2024, C.M. Tian, N. Jiang, S.J. Li, and Y.Y. Wu (holotypeBJFC-S2543). Ex-type culture CFCC 72599.
Etymology.
Named after the host genus, Rubus.
Description.
Sexual morph: not observed. Asexual morph: Conidiophores are mononematous or dichotomously branched, displaying a Virgariella-like to Nodulisporium-like branching pattern. Conidiophores smooth to finely roughened, hyaline, aseptate, with 2–3 conidiogenous cells at each terminus, measuring (14–)16–36(–39) × 2.5–4(–7) μm (av. ± S.D.= 27 ± 8.4 × 3.5 ± 1.2 µm, n = 30). Conidiogenous cells terminal or lateral, cylindrical, hyaline to pale yellow, smooth to finely roughened, with flattened base, producing conidia apically, measuring (11.5–)13–20.5 × 2–3.5 μm (av. ± S.D.= 16 ± 2.5 × 3 ± 0.4 µm, n = 30). Conidia ellipsoid to dacryoid, hyaline, aseptate, smooth to finely roughened, solitary, mostly flat-based, holoblastic-sympodial, measuring 4.5–8 × 3–4.5 μm (av. ± S.D.= 7 ± 0.8 × 4 ± 0.3 µm, n = 50).
Daldinia rubi (BJFC-S2543) A, D. Leaf of host Rubus idaeus; B. Front colony morphology on PDA at 14 days; C. Reverse colony morphology on PDA at 14 days; E. Conidiomata development on medium; F–J. Conidiogenous cells and conidia; K. Conidia. Scale Bars: 500 μm (D); 200 μm (E); 10 μm (F–K).
Culture characters.
Colonies showed sparse, cobweb-like mycelium, appearing semi-transparent, pale gray, with small brown central structures. The reverse was pale grayish-brown with scattered black conidial masses. Aerial hyphae were sparse, growing to 60 mm on PDA in 7 days at 25 °C.
Other material examined.
China • Guizhou Province, Guiyang City, Yunyan District, Qianlingshan Forest Park, 26°36'06"N, 106°41'42"E, on the leaf spots of Rubus idaeus, 26 Jul 2024, C.M. Tian, N. Jiang, S.J. Li and Y.Y. Wu, BJFC-S2544, living cultures CFCC 72600.
Notes.
Based on multi-locus phylogenetic analysis, the two isolates (CFCC 72599, CFCC 72600) formed an independent clade with 100% MP, 100% ML, and 1.00 BYPP values, clearly distinct from Daldinia ehretiae in the multi-locus analyses (Fig. 1). To further substantiate the recognition of D. rubi as a new species, a comprehensive comparison of asexual morphological traits within the Daldinia genus in China was conducted (Table 3). Morphologically, D. rubi can be readily distinguished from D. ehretiae by multi-trait divergence: as shown in Table 3, D. rubi produces larger conidia (ellipsoid to dacryoid, 4.5–8 × 3–4.5 μm) compared to D. ehretiae (ellipsoid or cylindrical, 4.2–6.6 × 1.7–2.8 μm). For conidiophores, D. rubi has shorter and narrower structures (mononematous or dichotomously branched, 16–36 × 2.5–4 μm) with more conidiogenous cells per terminus (2–3 cells) than D. ehretiae (mononematous or dichotomously branched, 100–210 × 3.1–4.3 μm, 1–2 cells per terminus). Conidiogenous cells of D. rubi are shorter in length (cylindrical or laterally cylindrical, (11.5–)13–20.5 × 2–3.5 μm) versus D. ehretiae (cylindrical, 16.8–24.5 × 2.7–4.1 μm). Molecularly, D. rubi also shows clear divergence from D. ehretiae. There is a 12 bp difference in ITS sequences (376 characters, 96.8% similarity, including one gap) and a 26 bp difference in tub2 sequences (745 characters, 96.5% similarity, no gaps). Collectively, the independent phylogenetic position, distinct morphological traits (as detailed in Table 3 for a comparison of asexual characteristics among Daldinia species in China), and molecular divergence confirm that D. rubi represents a new species.
Discussion
In this study, two Daldinia species, D. eschscholtzii and D. rubi, were discovered on leaf spot samples of Indocalamus hispidus and Rubus idaeus in Guizhou Province, China. This represents the first record of Daldinia on host plants belonging to Rubus (Rosaceae) and Indocalamus (Poaceae). Globally, Daldinia exhibits an exceptionally broad host range, with the USDA database documenting over 600 host species (https://fungi.ars.usda.gov/). Within China, previously reported Daldinia species have primarily been found on woody plants of Fagaceae, Lauraceae, Sapindaceae, Moraceae, and Betulaceae (https://fungi.ars.usda.gov/). By contrast, records on Rosaceae and Poaceae hosts remain scarce. The present study significantly expands the known host range of Daldinia in China.
Notably, Daldinia exhibits distinctive biological characteristics in its sexual morph, most remarkably through the formation of large stromata on woody branches. These structures characteristically display concentric zonation patterns internally and produce ellipsoidal, brownish to dark brown ascospores (Stadler et al. 2001a; Stadler et al. 2004; Stadler et al. 2014; Liu et al. 2025). However, our investigations, consistent with previous studies, have failed to observe a sexual stage in Daldinia specimens collected from diseased foliage in Southwest China (Yin et al. 2024). This phenomenon suggests that the life cycle transition in Daldinia may be influenced by an intricate interplay of environmental conditions, climatic factors, and host-specific characteristics. Moreover, Daldinia may initially colonize leaves as endophytes or latent pathogens. When host vigor declines or environmental conditions become favorable, these fungi may exhibit pathogenicity. Subsequent studies should conduct artificial inoculation experiments to confirm Daldinia’s pathogenicity and determine how environmental factors and host conditions affect its virulence.
This study systematically compiled the global host range of D. eschscholtzii, revealing its ability to parasitize over 50 plant families while demonstrating distinct host preferences (Suppl. material 2). These fungi primarily colonize dicotyledonous plants, with a particular affinity for the Rosales and Fabales orders, showing the highest frequency on the Fabaceae, Lauraceae, and Moraceae families. Notably, D. eschscholtzii exhibits cross-clade infectivity, capable of rarely colonizing gymnosperms (Pinaceae) and ferns. As shown in Suppl. material 2, tropical-affiliated families (e.g., Lauraceae and Myrtaceae) are disproportionately represented among hosts, many of which include economically significant crops such as citrus and soybean. These characteristics reflect an evolutionary balance between broad-spectrum infectivity and specialized host adaptation in D. eschscholtzii.
Current studies reveal a wide distribution of Daldinia fungi around the world, demonstrating their strong adaptability to diverse ecological environments (Chen 2002; Stadler et al. 2014; Yin et al. 2024; Liu et al. 2025). This suggests potential undiscovered Daldinia species across various ecosystems, warranting more systematic investigations. Targeted sampling in special habitats (e.g., karst formations) and non-conventional hosts would particularly advance our understanding of the biodiversity and ecological functions of this genus.
Supplementary Material
XML Treatment for Daldinia eschscholtzii
XML Treatment for Daldinia rubi
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bitzer JKöpcke B Stadler M Hellwig V Ju YM Seip S Henkel T (2007) Accelerated dereplication of natural products, supported by reference libraries.Chimia 51: 332–338. 10.2533/chimia.2007.332 · doi ↗
- 2Cesati V De Notaris G (1863) Schema di classificazione degle sferiacei italici aschigeri piu’ o meno appartenenti al genere Sphaeria nell’antico significato attribuitoglide Persono.Commentario della Società Crittogamologica Italiana 1(4): 177–420.
- 3Chan CL Yew SM Ngeow YF Na SL Lee KW Hoh CC Yee WY Ng KP (2015) Genome analysis of Daldinia eschscholtzii strains UM 1400 and UM 1020, wood-decaying fungi isolated from human hosts. BMC Genomics 16: 966. 10.1186/s 12864-015-2200-2PMC 465094226581579 · doi ↗ · pubmed ↗
- 4Chen MM (2002) Forest fungi phytogeography: Forest fungi phytogeography of China, North America, and Siberia and international quarantine of tree pathogens. California: Pacific Mushroom Research and Education Center, Sacramento.
- 5Child M (1932) The genus Daldinia. Annals of the Missouri Botanical Garden 19: 429–496. 10.2307/2394130 · doi ↗
- 6Crous PW Gams W Stalpers JA Robert V Stegehuis G (2004) Myco Bank: An online initiative to launch mycology into the 21st century.Studies in Mycology 50(1): 19–22.
- 7Crous PW Verkleij GJM Groenewald JZ Houbraken J (2019 a) Fungal Biodiversity. Westerdijk Laboratory Manual Series No. 1. Utrecht: Westerdijk Fungal Biodiversity Institute.
- 8Crous PW Verkleij GJM Groenewald JZ Houbraken J (2019 b) Westerdijk Laboratory Manual Series No. 1. Fungal biodiversity. CBS Laboratory Manual Series, 2nd edn. Utrecht: Westerdijk Fungal Biodiversity Institute.