Dynamic Interactions between the Gut Microbiome, Health, and Metabolic Disorders in Preterm Infants
Ki Beom Jang, Eunsol Seo, Younghoon Kim

TL;DR
This review explores how preterm birth disrupts gut microbiome development, leading to health issues and metabolic disorders in infants.
Contribution
The paper integrates recent findings on gut microbiome dynamics and their impact on preterm infant health, emphasizing the gut-brain axis and microbial interventions.
Findings
Preterm infants show gut dysbiosis linked to diseases like NEC and sepsis.
Commensal microbes influence immune balance and gut-brain signaling.
Targeted nutrition and probiotics may improve microbial balance in preterm infants.
Abstract
Preterm birth, which occurs before the entire gestation is completed, causes serious health challenges for newborns. The gastrointestinal tract often shows delayed development and poor function in nutrient utilization and immune response. Preterm neonates are faced with difficulties in growth, are more vulnerable to infections, and have a higher risk of developing inflammatory diseases. One key factor in these outcomes is the abnormal development of intestinal microbiota. In premature neonates, gut microbes tend to show lower diversity and changes in several bacteria. The gut dysbiosis can disrupt gut function and contribute to complications like necrotizing enterocolitis, sepsis, and even long-term metabolic and neurodevelopmental disorders. This review brings together current research on how premature birth affects the growth, gut development, and overall health of infants, with a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
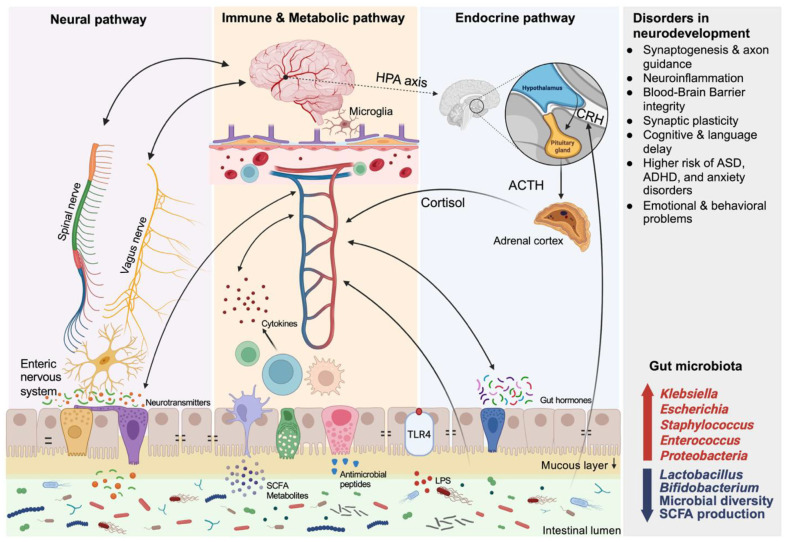 Figure 1
Figure 1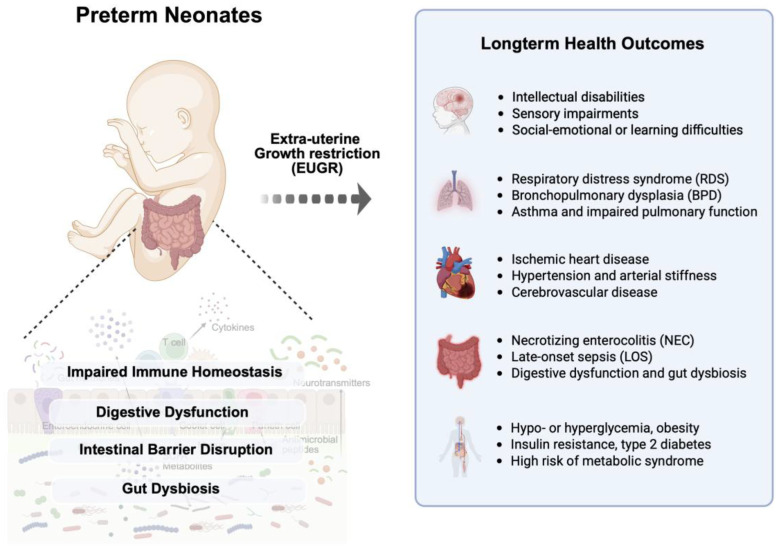 Figure 2
Figure 2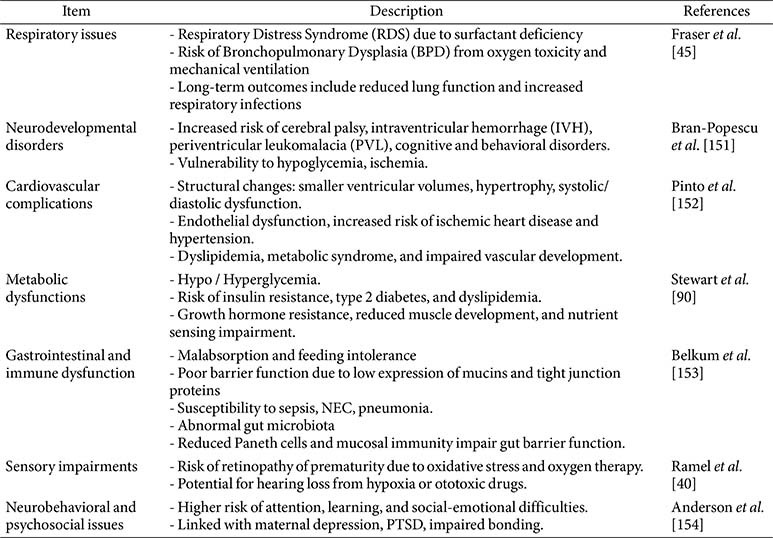 Figure 3
Figure 3 Figure 4
Figure 4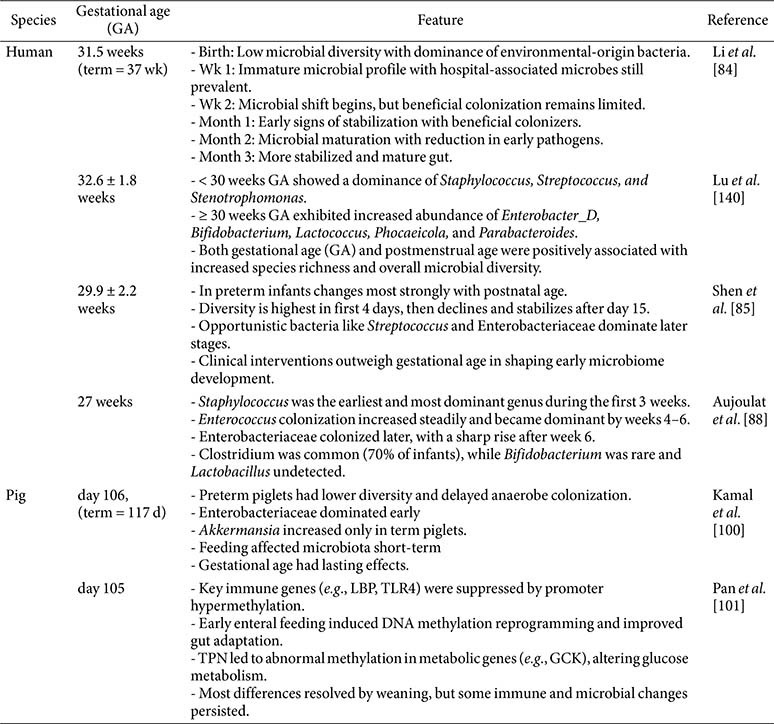 Figure 5
Figure 5- —National Research Foundation of Koreahttp://dx.doi.org/10.13039/501100003725
- —Ministry of Education, Science and Technologyhttp://dx.doi.org/10.13039/501100004085
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDiet and metabolism studies · Hyperglycemia and glycemic control in critically ill and hospitalized patients · Infant Nutrition and Health
Introduction
Prematurity refers to the birth of an animal before the completion of the gestation period. Preterm birth in humans, defined as delivery before 37 weeks of gestation, affects approximately 13.4 million infants annually, approximately 10% of global births according to the World Health Organization [1]. Preterm birth also occurs in livestock production, due to low survival rates and the lack of a standard framework for preterm livestock birth [2]. Preterm birth is not commonly recognized as a distinct category of concern in animal production, and it is mainly highlighted as an animal model for human research. However, preterm birth can occur and negatively affect production by increasing mortality, compromising growth, and raising disease susceptibility [3]. Preterm birth is also recognized as a critical challenge, contributing to various health issues with a higher risk of extra-uterine growth retardation, neurodevelopmental impairment, and metabolic diseases later in life [4]. These adverse outcomes are closely related to the insufficient development of internal organ systems, including the gastrointestinal, respiratory, and immune systems, during the gestational period [5]. Unfortunately, the cause of preterm birth is not clear due to multiple mechanisms of various biological and environmental factors, such as uteroplacental health, maternal condition, viral and bacterial infection, chronic stress, or other immunological responses [6].
The intestine is a major organ of the digestive system that is exposed to the external environment, playing a central role in nutrient digestion and absorption, and maintaining systemic homeostasis through its mucosal immune function [7]. However, preterm birth leads to gastrointestinal immaturity, resulting in impaired nutrient utilization and intestinal barrier function [8]. These vulnerabilities, combined with distinct patterns of microbial colonization, are compared with those of term infants. Such alterations increase susceptibility to neonatal conditions, including sepsis-associated enteropathy and necrotizing enterocolitis (NEC) in the neonatal period, and may also contribute to an increased risk of developing chronic disorders like inflammatory bowel disease (IBD) later in life [9]. The development of intestinal microbiota can also be affected by clinical management practices associated with preterm birth, including delivery method, feeding practices, or early exposure to medication [10, 11]. Extensive research, conducted in humans, animal models, and in vitro systems, has highlighted the pivotal role of the intestinal microbiota in host physiology [12?-14]. In preterm infants, the development of the intestinal microbiota is often altered and suboptimal, typically characterized by delayed colonization, reduced diversity, and gut dysbiosis compared to term infants [15, 16]. Considering the significant impact of early-life microbiota on host development, it is critical to understand how microbial colonization is shaped by prematurity. This review aims to summarize current knowledge on the development of the intestinal microbiota in preterm neonates and examine the implications of microbial development for health and disease.
Effects of Prematurity on Growth and Health
Body Composition Development
Preterm neonates typically have a higher proportion of body fat mass than term neonates, which is negatively correlated with gestational age and positively associated with body weight increase [17, 18]. According to a meta-analysis conducted by Johnson et al. [19], preterm infants had a 3% greater total body fat and much less fat-free mass, about 460 g in the body, compared to full-term infants. These changes may increase the risk of developing metabolic syndrome and cardiovascular diseases in the future [20]. Viswanathan et al. [21] demonstrated that maintaining a healthy balance between fat-free mass and fat mass is crucial for neurodevelopment and metabolic health in preterm infants. Based on previous findings, preterm infants have different body compositions, which can be explained by several reasons. Firstly, immature metabolic systems and challenges hinder effective nutrient utilization, affecting growth and body composition [22, 23]. The immature gastrointestinal system in preterm infants can lead to malabsorption and feeding difficulties, further impacting their nutrient intake and utilization [24]. In addition, hormonal secretion imbalances play a significant role in the altered body composition of preterm infants [22, 25]. One of the causes suggested for the impacts of prematurity is the extreme environmental stress [26] after birth, leading to increased circulating cortisol levels, which in turn affect leptin metabolism and the hypothalamic-pituitary-adrenal (HPA) axis [27]. The endocrine system, which regulates body metabolism, would not be fully developed in preterm infants. Key metabolic hormones may not function optimally, impairing nutrient metabolism [28]. The hormonal dysregulation can result in increased fat deposition and altered muscle development [28]. Understanding these factors is crucial for developing effective nutritional and medical strategies to support the growth and development of preterm infants [29, 30].
Impact of Prematurity on Physiological and Metabolic Development
Prematurity can adversely affect several organ systems, including the neurological, respiratory, and cardiovascular systems (Table 1). During the neonatal period, brain growth and maintenance require a continuous energy supply. However, providing adequate nutrition to premature infants is challenging due to immature gastrointestinal function. Total parenteral nutrition (TPN) is commonly used but is limited in the amount and duration it can be safely administered. Long-term TPN use may impair intestinal development and induce metabolic liver dysfunction [31]. Therefore, early enteral feeding is initiated, and TPN is gradually reduced while carefully introducing digestive feeding [32]. Under nutrient-deficient conditions, tissue and protein breakdown provide alternative energy sources through gluconeogenesis from amino acids or ketogenesis for energy needs [33].
The blood system plays a central role in supporting growth and development by transporting nutrients and oxygen, removing waste products via the liver and kidneys, contributing to systemic immune defense, and facilitating adaptation to environmental changes. Preterm birth is also associated with an increased risk of cardiovascular diseases, including cardiometabolic and cardiopulmonary morbidities later in life [34, 35]. Adults born preterm often exhibit alterations in cardiac structure and function, such as smaller ventricular volumes, increased ventricular mass, and both systolic and diastolic dysfunction [36]. Furthermore, endothelial dysfunction, an early marker of cardiovascular diseases, is more prevalent in preterm infants than in full-term infants [37]. A properly functioning endothelium is crucial for maintaining vascular homeostasis, as it balances factors that regulate both vasoconstriction and vasodilation. Endothelial dysfunction is exacerbated by factors such as dyslipidemia, impaired nutrient metabolism, and atherosclerotic disease. A previous study found that preterm young adults have significantly lower flow-mediated dilation than term-born controls, indicating reduced vascular endothelial function [38]. This suggests an increased risk of cardiovascular disease due to preterm birth. Preterm birth is also linked to an increased risk of developing ischemic heart disease, characterized by reduced blood flow to the heart due to narrowed or blocked coronary arteries [39].
Premature infants have significant challenges in the respiratory system [40]. One contributing factor is the frequent oxygen supply in intensive care management, often due to prematurity or respiratory conditions, which can result in excessive oxygenation of tissues [41]. Mohammadi et al. [42] showed that hyperoxia-induced oxidative stress by high oxygen supply can disrupt mitochondrial function, leading to injury in the developing lungs and brain of extremely premature infants. Preterm birth also significantly increases the risk of long-term respiratory diseases. Preterm infants tended to have respiratory distress syndrome due to a deficiency in pulmonary surfactant, crucial for keeping the alveoli in the lungs stable [43]. Previous research evidence showed that the severity of respiratory distress syndrome and mortality rates are negatively correlated to the gestation day, and preterm models showed immature alveolar architecture and minimal surfactant protein-B expression [44]. Bronchopulmonary dysplasia is another common complication in preterm infants. It is characterized by inflammation, scarring, and impaired lung development, ultimately leading to chronic respiratory disease [44, 45]. Prematurity disrupts normal alveolar development, resulting in fewer and larger alveoli and a reduced surface area for gas exchange, which contributes to chronic lung disease [46]. These impairments often necessitate intensive postnatal interventions, including mechanical ventilation, surfactant replacement, and oxygen therapy [45]. Despite advances in perinatal care, such as antenatal corticosteroids and exogenous surfactants, the incidence of chronic lung disease remains higher in preterm infants compared to those born at term [47]. Chronic lung disease, defined by the need for respiratory support or supplemental oxygen at 36 weeks post-conception, is associated with several factors, including the degree of prematurity, severity of initial lung disease, and duration of mechanical ventilation and oxygen therapy [47]. Prematurity interrupts alveolar development, resulting in reduced surface area for gas exchange and leading to chronic lung disease. Preterm infants have immature immune systems, making them more susceptible to respiratory infections, which can exacerbate chronic respiratory issues [48]. Chronic respiratory conditions can lead to reduced physical activity and endurance, with respiratory muscle weakness further impairing muscle growth and function.
Neurological impairments represent another major consequence of preterm birth. Critical phases of brain development occur between 34 and 40 weeks of postmenstrual age [39]. Preterm birth interrupts these processes, increasing the risk of long-term conditions such as cerebral palsy, cognitive impairments, and behavioral disorders [49]. Structural abnormalities in the brain are common and elevate the risk of intraventricular hemorrhage due to fragile germinal matrix vasculature, unstable cerebral blood flow, and coagulation issues [50]. Although most intraventricular hemorrhages are mild, severe cases may result in permanent brain injury. Periventricular leukomalacia, involving white matter damage due to hypoxia or ischemia, is also associated with cerebral palsy and other motor deficits in preterm infants [51]. Long-term challenges may include intellectual disabilities, sensory impairments, growth delays, and social-emotional or learning difficulties [52]. The brain’s high energy demand—accounting for roughly 60% of total metabolism—is particularly taxing in premature infants and contributes to rapid depletion of energy stores, especially during periods of active synaptogenesis and cortical development [53, 54].
Metabolic dysfunction is common in preterm infants. Immature digestive and endocrine systems can lead to impaired insulin secretion and glucose homeostasis, resulting in hypo- or hyperglycemia [55]. Impaired glucose regulation in early life increases the risk of insulin resistance and type 2 diabetes later in life [55]. In addition, altered lipid metabolism can lead to elevated triglyceride and cholesterol levels, further increasing cardiovascular risk [56]. Muscle development may also be compromised due to disrupted nutrient sensing and insulin signaling, resulting in reduced muscle mass and strength [57, 58]. Growth hormone resistance is another issue that can negatively affect physical development [57]. Finally, preterm infants are at increased risk for deficiencies in micronutrients such as iron, calcium, and vitamin D, which can lead to anemia, poor growth, and developmental delays.
Extrauterine Growth Restriction and Immaturity of Gastrointestinal Tract
Preterm infants are not fully developed in the placenta during gestation and are then born earlier than their designated birth date, resulting in exposure to extra environments with immature digestive and immune systems [59, 60]. In particular, the small intestine has a malfunction in intestinal function and structure, leading to a high risk of inefficiency in nutrient utilization, reduced immune function, and low resistance to disease or viral infection simultaneously [32, 61]. These features are closely related to the vitality and health of preterm infants, making early maturation of intestinal function a critical target in neonatal care strategies [8, 32]. Extrauterine growth restriction (EUGR) commonly occurs in preterm infants after birth with several health challenges [59, 60]. Advancements in neonatal intensive care have significantly improved survival rates, optimizing growth, promoting healthy body composition, and enabling the development of normally functioning organs in premature infants [5, 40]. Without suitable postnatal nutrition, very low birth weight infants struggle with catch-up growth, adversely affecting long-term growth and neurodevelopment [19, 53]. EUGR is a significant issue for very low body weight infants, and their growth retardation results from complex interactions, including health concerns such as endocrine abnormalities, central nervous system damage, difficulties in nutrient intake, and application of medications, or nutritional metabolism [25, 28, 62]. According to Clark et al., EUGR significantly affects the postnatal growth of preterm infants, with about 28% for body weight reduction at discharge from the intensive care unit [63]. Chien et al., have also shown that neurodevelopmental outcomes, considering cognitive and movement impairments in infants with low birth weights, are closely associated with the severity of their EUGR[64]. In a preterm rat model, EUGR elevated pulmonary arterial pressure in adults and genome-wide epigenetic modifications in pulmonary vascular endothelial cells by increased CpG methylation and H3K27me3 in the Notch1 gene promoter region [65]. Interactions of several environmental factors can exacerbate negative outcomes caused by EUGR. In particular, inadequate early postnatal nutrition is recognized as a leading cause of EUGR. Malnutrition during the critical period of brain development is associated with deficiencies in behavior, learning, and memory, and reduced brain cells [66]. Early and aggressive nutrition aims to minimize cumulative caloric and protein deficits during the acute phase after birth to improve cognitive and neurodevelopmental outcomes related to extra-uterine growth restriction [33, 40].
The stomachs of preterm infants have relatively low acid secretion and decreased proteolytic enzyme activity, resulting in unusually high pH levels and reduced pepsin activity, which can impair protein digestion [67, 68]. This may restrict the absorption of essential amino acids for growth and potentially increase the uptake of allergenic antigens [67]. In addition, insufficient gastric acidity weakens the first line of defense against pathogens, increasing the risk of infection [68, 69]. The intestinal mucosa is also premature, resulting in a structurally weak barrier and significantly lower expression of protective proteins such as mucins, trefoil factor family peptides, and antimicrobial peptides compared to term infants or adults [70, 71]. In particular, the expression of tight junction proteins, critical components for intestinal barrier function, is insufficient, causing a “leaky gut condition characterized by increased intestinal permeability [72]. Increased intestinal permeability allows bacteria and toxins to more easily translocate into the body. Such translocation is a major mechanism underlying systemic inflammatory responses and the development of NEC [72].
Regarding an immunological perspective, preterm infants also have an underdeveloped population of Paneth cells and a lack of the innate immune system, limiting their ability to regulate intestinal microbial balance [73, 74]. The initial colonization of gut microbiota is delayed, and there is often an abnormal predominance of pathogenic Gram-negative bacteria, such as Proteobacteria [11, 15, 16]. This dysbiosis triggers inflammatory responses, leading to epithelial cell damage and disruption of immune homeostasis [9, 15, 16]. Notably, NEC, a severe complication that typically occurs within the first 2–3 weeks of life, is strongly associated with such gut dysbiosis [9, 15, 16]. In addition, the proportions and functions of the four major epithelial cell types—enterocytes, goblet cells, enteroendocrine cells (EECs), and Paneth cells—that actively activate in the intestine remain poorly defined in preterm neonates [8, 32]. Enterocyte efficiency is low, and the number and function of goblet cells, which are responsible for mucus secretion, are insufficient [75]. As a result, the intestinal barrier against pathogens is incomplete. Immune cells, such as lymphocytes, macrophages, and plasma cells, in the lamina propria are not fully established, which limits the function of gut-associated lymphoid tissue [48].
Intensive Management for Preterm Neonates
Preterm infants need various medical interventions for growth and development, resulting in physiological conditions distinct from those in utero. Non-invasive respiratory support, such as early continuous positive airway pressure (CPAP) and nasal intermittent positive pressure ventilation (NIPPV), is commonly preferred to avoid endotracheal intubation and reduce the risk of bronchopulmonary dysplasia (BPD) [76, 77]. However, when mechanical ventilation becomes necessary, it is associated with increased systemic inflammation and a higher risk of impaired growth and neurodevelopment, highlighting the importance of a timely transition back to non-invasive support [78].
Nutritional management is crucial for early growth and long-term development in preterm infants. Preterm neonates initially require parenteral nutrition until their digestive tract can sufficiently tolerate enteral feeds, as prolonged use of parenteral nutrition can have negative impacts on the development of the liver and digestive tract [79]. Enteral feeding modalities, which deliver milk either intermittently in specific volumes or continuously at a steady rate, have distinct physiological effects and clinical implications [80]. Researchers have compared bolus versus continuous feeding modalities, demonstrating distinct clinical characteristics [79, 80]. In addition to feeding modality, the importance of breast milk feeding is well established because human milk is superior to infant formula in terms of preterm nutrition, primarily due to its bioactive compounds, which include immunomodulatory factors, growth factors, vitamins, and trace elements [81]. These components enhance initial nutritional status and positively influence developmental trajectories in preterm infants. Antibiotic treatment is used to prevent infections in preterm infants. However, antibiotics can also elevate the risk of nephrotoxicity, hepatotoxicity, and antimicrobial resistance, highlighting the importance of careful monitoring and appropriate duration of treatment [82]. Overall, interventions implemented in the neonatal intensive care unit (NICU), including respiratory support, nutritional management strategies, and antibiotic use, significantly impact both immediate survival and the longer-term growth and developmental outcomes of preterm infants.
Microbial Colonization in Preterm Infants
Postnatal Progression of Microbial Establishment
The intestinal microbiota in preterm infants develops through a dynamic, sequential process (Table 2) that begins at birth and continues over the first two to three months of life [83, 84]. This early microbial colonization plays a fundamental role in shaping host immunity, metabolic programming, and gut maturation [74]. Shen et al.[85] showed that the development of gut microbiota in preterm infants follows a clear temporal pattern that correlates more strongly with postmenstrual age than with postnatal age, delivery mode, or antibiotic exposure, and progresses in defined stages dominated sequentially by Staphylococcus, Enterococcus, Enterobacter, and finally Bifidobacterium. In contrast to full-term infants, preterm neonates experience delayed and aberrant microbial succession, with significant clinical implications [86]. Initial colonization typically occurs at the time of birth. In preterm infants, the meconium microbiota is characterized by low diversity and is dominated by Proteobacteria and Firmicutes, with minimal representation of strict anaerobes such as Bifidobacterium and Bacteroides [74, 87]. Aujoulat et al. [88] observed that very preterm infants were largely colonized by facultative anaerobes such as Staphylococcus, Enterococcus, and Enterobacteriaceae, with diversity developing slowly and remaining distinct from that of term infants. Moreover, differences in birth weight, gestational age, and clinical exposures contribute to variability. Such cohort-specific differences are often difficult to interpret because various factors, including antibiotic exposure, feeding practices, and NICU management, simultaneously affect microbial development. For instance, infants weighing less than 1.1 kg often remain longer in the Enterococcus-dominant phase, indicating delayed maturation [89]. By 4 to 6 weeks, several preterm infants can begin to acquire more stable microbiota structures, with increasing Bifidobacterium and Lactobacillus and a gradual decline in Proteobacteria [86]. Still, extreme prematurity, prolonged antibiotic exposure, or extended NICU stay can sustain dysbiosis and delay convergence toward a term-like microbiota. Although microbial diversity remains low in early life, beta diversity shows convergence across infants over time [84]. Stewart et al. [90] further showed that functional maturation proceeds slowly, with early stages dominated by carbohydrate metabolism and later weeks shifting toward amino acid and lipid metabolic functions [86].
After birth, microbial colonization is readily influenced by extrinsic factors, including delivery mode, antibiotic exposure, feeding method, and the neonatal intensive care environment [91, 92]. Zeng et al. [93] further emphasized that gut dysbiosis in preterm infants is strongly associated with neonatal intensive care practices, including antibiotic use and parenteral nutrition, particularly in extremely low birth weight infants. Interestingly, the NICU management can influence the shape of early colonization patterns. Infants delivered by cesarean section typically acquire a microbiota enriched in skin‐derived Staphylococcus, whereas those delivered vaginally harbor maternal gut–derived taxa such as Escherichia and Bacteroides [89]. Cesarean delivery has also been linked to delayed establishment of Bifidobacterium and reduced Bacteroides colonization, whereas vaginal delivery promotes faster microbial succession and higher overall diversity [89]. In addition, continuous exposure to the NICU environment, including medical equipment, surfaces, and staff contact, promotes the establishment of hospital-associated microbes [91]. NICU stay has been associated with persistence of Klebsiella, Enterococcus, and other Enterobacteriaceae, often delaying the transition toward beneficial anaerobes. With these influences, subsequent modulation of the gut microbiota is also strongly influenced by clinical interventions such as antibiotic use and feeding strategies. Intrapartum or postnatal antibiotic administration markedly reduces microbial diversity and delays the expansion of beneficial commensals [89]. Specifically, antibiotics diminish early Bifidobacterium colonization and favor expansion of Proteobacteria, particularly Enterobacteriaceae, and in some cases increase fungal colonization such as Candida. Feeding mode can also affect the microbial colonization: breast milk containing human milk oligosaccharides can support the dominance of Bifidobacterium, whereas formula feeding is associated with delayed or altered microbial colonization. Breast milk promotes enrichment of B. breve and B. longum, which produce short-chain fatty acids that acidify the lumen and suppress pathogens. In contrast, formula-fed infants show higher relative abundance of Clostridium, Enterobacter, and Streptococcus, taxa often linked to necrotizing enterocolitis risk. Moreover, donor milk and formula feeding policies can either accelerate or hinder the functional maturation of the microbiota, influencing the timing of transition toward adult-like communities [92]. Although donor milk confers more protection than formula, pasteurization and processing reduce microbial transfer and alter the metabolic profile compared to breast milk.
Feeding modality has a more distinct influence on microbial development. Tauchi et al. [94] also found that breast milk feeding in preterm infants was associated with higher levels of Bifidobacterium and lower levels of potentially pathogenic Proteobacteria, reinforcing the protective and microbiota-stabilizing effects of human milk. Direct breastfeeding promotes the transmission of maternal strains such as B. breve and B. longum, which are not efficiently transferred through formula [95]. Breast milk has also been shown to mitigate antibiotic-induced disruptions in microbial development [74]. In addition, fecal samples from preterm infants reveal strong taxonomic overlap across time, suggesting that early microbial imprinting can persist for several weeks [16, 84, 96]. Studies that follow babies over time show that the amount of weight a baby gains and the duration since pregnancy began are better indicators of gut bacteria development than simply counting the days after birth [83, 89]. Despite individual variation, the overall trajectory in preterm infants reveals delayed establishment of strict anaerobes, high initial dominance by facultative pathogens, and prolonged dysbiosis in the absence of supportive interventions [97, 98]. These findings highlight the neonatal period as a critical window in which targeted interventions could redirect microbial development and improve long-term outcomes.
Microbial Composition and Diversity in Preterm Infants
The intestinal microbiota of preterm infants differs significantly from that of term infants, influenced by developmental immaturity and postnatal management. These differences lead to taxonomic shifts, altered functional capabilities, and modified metabolic activities, carrying profound implications for the development of the immune system and long-term health outcomes. Preterm infants predominantly exhibit intestinal microbiota characterized by facultative anaerobes, notably members of Proteobacteria such as Enterobacteriaceae, Klebsiella, and Escherichia coli, accompanied by reduced colonization by obligate anaerobes including Bifidobacterium, Bacteroides, and Clostridia [87, 96, 99]. Table 3 summarizes key features of gut microbiota development based on previous findings. Preterm piglets are a valuable model for prematurity study, as they have also been shown reduced microbial diversity, delayed anaerobe colonization, and epigenetic alterations in immune and metabolic genes influenced by feeding strategies [100, 101]. The dysbiosis results from delayed microbial succession, which is exacerbated by clinical practices such as cesarean delivery, antibiotic use, and limited breastfeeding opportunities inherent in neonatal intensive care units [97, 102].
Longitudinal studies consistently indicate lower microbial diversity in preterm infants, particularly during the initial postnatal weeks. Chu et al., [103] reported a significant reduction in Shannon diversity indices by 0.45 units in preterm infants compared to term infants within the first 10 days post-birth, a phenomenon independent of feeding modality or antibiotic exposure but closely associated with the length of NICU management. Although alpha-diversity tends to increase over time, it often remains lower than that observed in term infants even beyond early childhood [96, 104, 105]. Beta-diversity, which indicates differences in microbial composition between individuals, initially exhibits considerable variability among preterm infants due to diverse clinical interventions and environmental exposures, but typically converges as infants mature [84, 96]. Enterotype transitions, characterized by shifts from dominance of Staphylococcus or Enterobacter to stable Bifidobacterium- or Bacteroides-rich profiles, occur later in preterm infants, particularly in cases of extreme prematurity or prolonged antibiotic exposure [83, 86, 91]. Importantly, although microbial diversity and composition tend to stabilize over time, distinct microbial imprints persist into later childhood, characterized by delayed acquisition of adult-like microbiota and associated functional maturity [104, 105]. In addition, maternal microbiota during pregnancy has been shown to influence neonatal microbiota development, with mothers delivering preterm showing reduced gut diversity and decreased levels of beneficial bacteria such as Bifidobacterium and Streptococcus compared to mothers delivering at term [99]. Therefore, these findings underscore the complex interplay of developmental immaturity, clinical practices, and environmental exposures that shape the microbiota composition and diversity in preterm infants. Persistent dysbiosis and altered maturation pathways highlight the critical importance of targeted interventions during neonatal care to promote optimal microbial development and support long-term physiological health.
Feeding practices and probiotic supplementation have a significant impact on the microbiota. Breastfeeding promotes beneficial colonization by B. breve and B. longum, enhancing carbohydrate metabolism pathways [95, 106]. In contrast, formula feeding and cow’s milk-based fortifiers may encourage Proteobacteria proliferation and reduce microbial evenness [106]. Although probiotic interventions have demonstrated benefits such as increased SCFAs, lowered intestinal pH, and reduced abundance of pathogenic taxa, previous studies highlight variability in outcomes influenced by strain, dosage, and administration timing [16, 107]. According to previous studies, supplementation of Lactobacillus rhamnosus has been associated with reduced NEC incidence and improved barrier function [108, 109], whereas B. breve supplementation showed more modest benefits, with inconsistent effects on NEC prevention but improvements in metabolic outcomes for preterm infants [110, 111]. Probiotic effects are widely considered strain-specific, and recent meta-analyses confirm that L. rhamnosus GG used as a single strain reduced NEC risk, but showed no significant effects on late-onset sepsis or mortality, whereas observational studies did not replicate these benefits [112]. Despite these promising findings, results from randomized controlled trials remain inconsistent. Some large trials and network meta-analyses report significant reductions in NEC and mortality with certain probiotic regimens, whereas others show no clear benefit, reflecting heterogeneity in trial design, product formulations, and patient populations [113]. A review of probiotic-related sepsis in preterm infants also demonstrated that although probiotics may reduce the risk of NEC and death, but rare cases of probiotic-associated sepsis and fatal infections from contaminated products highlight safety concerns, highlighting the need for strict quality control and careful monitoring in extremely preterm infants [114].
Roles and Health of Intestinal Microbiota in Preterm Infants
Immune and Intestinal Development
In preterm infants, an underdeveloped gastrointestinal tract is often accompanied by gut dysbiosis, disrupting the synchrony between immune maturation and intestinal epithelial development [102, 115]. Preterm infants often show microbiome compositions characterized by reduced microbial diversity and dominance of facultative anaerobes, such as Klebsiella, Escherichia, Staphylococcus, and Enterococcus, significantly impairing mucosal defense and barrier integrity [115]. These bacteria are also known to produce little or no short-chain fatty acids and are primarily associated with pathogenicity, dysbiosis, and inflammation. Microbial metabolites, such as short-chain fatty acids (SCFAs), play a pivotal role in regulating immune responses and reinforcing epithelial defense [116]. Butyrate has been shown to strengthen tight junction integrity by upregulating claudin-1, occludin, and ZO-1 gene expression and promoting oxygen-sensing pathways through GPR43-mediated HIF-1α stabilization [117, 118]. Butyrate suppresses histone deacetylases (HDACs), enhancing FOXP3 transcription and thereby promoting the expansion of regulatory T cells [119]. It could also be related to IL-10 production, suppressing IL-6 and TNF-α, which are two cytokines frequently elevated in preterm infants with intestinal inflammation [120]. Intestinal cell types, such as Paneth cells and goblet cells, exhibit both reduced number and function in preterm infants [121]. Paneth cells, located at the base of intestinal crypts, are crucial for secreting antimicrobial peptides such as lysozyme, defensins, and RegIIIγ, which help to control microbial colonization and maintain intestinal homeostasis [122]. In preterm infants, Paneth cells display delayed differentiation, decreased granule formation, and significantly reduced antimicrobial peptide secretion [123]. Goblet cells, responsible for mucus production, also exhibit impaired functionality in preterm neonates [124]. The mucus layer produced by goblet cells serves as a primary physical barrier against microbial invasion [125]. Reduced mucus secretion, decreased mucin gene (MUC2) expression, and altered mucus composition in preterm infants lead to compromised barrier function and heightened susceptibility to pathogens and inflammatory stimuli [124]. This combined impairment of Paneth and goblet cells intensifies vulnerability to microbial invasion and inflammation, contributing to a higher incidence of severe intestinal conditions, such as NEC, in preterm infants [115].
Gut microbiota in full-term infants is more effective at inducing tolerogenic dendritic cells and anti-inflammatory immune profiles different from preterm neonates [126]. Dysbiosis in preterm infants often features Proteobacteria dominance and reduced microbial diversity, both of which correlate with heightened Th17/Th1 immune responses and impaired cell development [11, 126]. Elevated levels of IL-6 and IL-8 in serum samples of infants with NEC further highlight the pro-inflammatory bias associated with a disrupted microbiota [126]. Conversely, colonization by B. breve has been linked to enhanced mucin gene expression (MUC2, TFF3), improved barrier function, and a more regulated immune response [127]. Probiotic supplementation with beneficial strains has shown potential to enhance mucosal immunity, stimulate regulatory T-cell differentiation, increase IgA production, and restore barrier function by improving epithelial proliferation and differentiation [128, 129]. Moreover, breastmilk provides critical bioactive components such as milk oligosaccharides, immunoglobulins, and lactoferrin, which facilitate beneficial microbial colonization and reinforce intestinal barrier integrity [81, 130]. However, preterm infants frequently have limited capacity for breastfeeding, thus significantly reducing their intake of the biological benefits from maternal nutrition. This restricted access to breastfeeding exacerbates gut dysbiosis and further impairs immune and intestinal development. These findings collectively indicate that microbial signals are integral to coordinating both physical and immunological maturation of the neonatal gut, and interventions that support beneficial colonization may offer long-term protective benefits in premature populations.
Necrotizing enterocolitis and neonatal sepsis are two of the most devastating complications associated with prematurity, both closely linked to intestinal dysbiosis. NEC is predominantly observed in very-low-birth-weight infants, with a mortality rate reaching up to 30% in severe cases [131]. A hallmark of NEC pathogenesis is the aberrant activation of innate immune signaling in the immature intestine, especially through Toll-like receptor 4 (TLR4) in response to lipopolysaccharide (LPS) derived from Gram-negative bacteria. This leads to exaggerated inflammatory cascades, epithelial injury, and compromised barrier function, ultimately resulting in intestinal necrosis [131?-133]. Previous studies have demonstrated that TLR4 is overexpressed in the premature gut and its activation by endotoxin impairs intestinal perfusion via endothelial nitric oxide synthase signaling [134]. Additionally, mice deficient in TLR4 show resistance to NEC, underscoring its central role in disease onset [132]. Clinically, NEC is preceded by a characteristic shift in the gut microbiota that typically a dominance of Proteobacteria and a depletion of Firmicutes and Bifidobacterium occurring 2–3 weeks before symptom onset [135, 136]. This pattern was also observed in a previous study, which showed that neonates who developed NEC or feeding intolerance had reduced microbial diversity and significantly lower levels of Bifidobacterium and Streptococcus in their fecal samples [84]. These microbial patterns are further exacerbated by clinical practices such as prolonged antibiotic use, cesarean section, and formula feeding, which suppress commensal colonization and amplify pathogenic proliferation [131]. Neonatal sepsis shares overlapping microbial and immunological features with NEC. Intestinal dysbiosis facilitates the translocation of gut-derived pathogens into the bloodstream, especially under conditions of barrier disruption. In a multi-site prospective study, over 80% of bloodstream isolates in septic neonates matched strains found in their stool samples, indicating gut-origin bacteremia [137]. Antibiotic-induced depletion of protective commensals such as Bifidobacterium and Lactobacillus allows for opportunistic expansion of Enterobacteriaceae, increasing sepsis risk [131]. Furthermore, the presence of specific microbial signatures, including elevated abundance of Klebsiella and Enterococcus prior to sepsis onset, has been reported in stool samples of infants who subsequently developed sepsis [138]. This suggests the potential of gut microbial profiling as a predictive biomarker for both NEC and sepsis.
Gut-Brain Axis and Neurodevelopment
The development of the gut-brain axis during early life plays a critical role in shaping long-term neuro-developmental outcomes, especially in preterm infants (Fig. 1). Communication between the gut and the brain occurs through three interconnected pathways: the neural pathway mediated by the vagus and spinal nerves, the immune and metabolic pathway involved in cytokines, microglial activation, blood–brain barrier regulation, and microbial metabolites such as SCFAs, and the endocrine pathway by the hypothalamic–pituitary–adrenal (HPA) axis and cortisol release. Immaturity or dysregulation of these pathways in preterm infants disrupts critical neurodevelopmental processes, resulting in impaired synaptogenesis, heightened neuroinflammation, reduced blood–brain barrier integrity, and diminished synaptic plasticity. Such alterations increase susceptibility to cognitive and language delays, autism spectrum disorder (ASD), attention-deficit/hyperactivity disorder (ADHD), anxiety, and a range of emotional or behavioral difficulties. These infants often exhibit delayed colonization of commensals, reduced microbial diversity, and increased proportions of pathobionts. Gut microbiota can be a key factor in this process, as preterm infants typically display an overrepresentation of pathobionts such as Klebsiella, Escherichia, Staphylococcus, Enterococcus, and members of Proteobacteria, alongside a reduction in beneficial taxa such as Lactobacillus and Bifidobacterium. This gut dysbiosis, characterized by decreased microbial diversity and reduced SCFA production, further compromises both intestinal and neurodevelopmental health. Such dysbiosis has been linked to neurodevelopmental impairments, including autism spectrum disorder, ADHD, and anxiety [139]. Lu et al. [140] demonstrated that transplanting fecal microbiota from preterm infants of lower postmenstrual age into germ-free mice led to significantly reduced associative fear learning and altered gene expression in the brain. Similarly, Carlson et al. [141] found that higher microbial diversity in infants under one year of age was positively correlated with enhanced cognitive scores based on the Bayley Scales of Infant and Toddler Development, a standardized assessment that evaluates cognitive, language, and motor skills in infants and toddlers.
The gut microbiota modulates brain development via immune regulation, microbial metabolite signaling, and neuroinflammatory pathways [142]. Among these, short-chain fatty acids have been shown to influence blood-brain barrier integrity, microglial maturation, and synaptic plasticity [143, 144]. Lu et al. [144] reported that mice receiving preterm microbiota displayed reduced brain IGF-1 levels, lower NeuN expression, and diminished myelin basic protein (MBP), indicating impaired brain maturation. In addition, Wang et al. [145] used Mendelian randomization analysis to reveal that SCFA-producing Faecalibacterium is causally associated with a reduced risk of ADHD, further supporting a role for microbial metabolites in neurodevelopment. Early-life dysbiosis also impacts tryptophan metabolism and serotonin biosynthesis, both of which influence synaptogenesis and axon guidance [139]. Mhanna et al. [146] reviewed how gut microbes regulate biosynthetic pathways of neurotransmitters such as GABA, dopamine, and serotonin, implicating these in the pathophysiology of ASD and anxiety. Beghetti et al. [139] specifically noted that dysbiosis in preterm infants suppresses tryptophan–serotonin pathways, thereby disrupting neural connectivity and neuron maturation.
The physiological architecture of the gut–brain axis includes peripheral and central components such as the enteric nervous system, vagus nerve, HPA axis, and limbic structures. In preterm neonates, stress-related signals originating in the gut may be transduced via vagal or humoral routes, activating the HPA axis and influencing emotional regulation circuits. Environmental factors such as maternal separation, intensive care unit stress, or intestinal inflammation can exacerbate these responses, particularly affecting the hippocampus and long-range neural connectivity.
The connection between gut dysbiosis and neuroinflammation is especially evident in models of NEC. Xia and Claud [142] demonstrated that gut-derived IFN-γ-producing CD4^+^ T cells migrate to the brain, where they activate microglia, leading to neurotoxicity and white matter injury. Previous studies [131, 147] further described that dysbiosis in preterm neonates impairs myelination and triggers microglial activation, contributing to impaired motor outcomes. Sherman et al. [148] also reported that inflammatory cytokines and microbial toxicants derived from the dysbiosis can enter systemic circulation and reach the brain, leading to white matter injury and long-term neurodevelopmental damage even in the absence of intracranial hemorrhage. These microbe–immune–neural interactions also influence stress circuitry through the hypothalamic-pituitary-adrenal (HPA) axis. Beghetti et al. [139] suggested that preterm infants experiencing early dysbiosis exhibit HPA hyperactivation, potentially leading to long-term emotional and behavioral dysregulation. Dysregulation of gut hormones and neuropeptides (e.g., ghrelin, peptide YY, GLP-1) in response to gut dysbiosis can exacerbate these disorders [148]. Krupa-Kotara et al. [149] provided evidence that such dysbiosis-induced alterations in stress circuits can be reversed through probiotic intervention.
Human studies have shown specific microbial patterns, such as increased Bacteroides and Lachnospiraceae abundance, to improve cognitive and language scores at two years of age [150]. Prtty et al. [150] reported that very preterm infants with higher alpha diversity and lower Proteobacteria levels in early fecal samples showed significantly higher Bayley cognitive and language scores at age two. Colonization by beneficial commensals such as Faecalibacterium prausnitzii and Bacteroides thetaiotaomicron promotes mucosal barrier integrity and anti-inflammatory signaling, thereby protecting the developing brain from microbial translocation and neuroimmune stress [148]. Moreover, maternal and neonatal probiotic supplementation has demonstrated neurodevelopmental benefits in animal models, suggesting possible benefits in preterm human infants [139]. Probiotic bacteria can inhibit proinflammatory signaling cascades (e.g., TNF-α and NF-κB), enhance IL-10 production, and reduce gut-derived systemic inflammation, ultimately preventing brain injury in preterm models.
Conclusion
In conclusion, gut dysbiosis is not only an acute challenge of prematurity but also an underlying cause of long-term health outcomes (Fig. 2). These microbial alterations increase the risk of severe neonatal diseases, including NEC and sepsis, and contribute to persistent neurodevelopmental and metabolic disorders. Commensal microbes and their metabolites play a pivotal role in epithelial maturation, immune regulation, and gut–brain signaling. Therefore, advancing neonatal care will require precision strategies that integrate microbial profiling with targeted nutritional interventions and a deeper understanding of host–microbe interactions to promote healthier outcomes in preterm infants.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ohuma EO Moller AB Bradley E Chakwera S Hussain-Alkhateeb L Lewin A 2023 National, regional, and global estimates of preterm birth in 2020, with trends from 2010: a systematic analysis Lancet 4021261127110.1016/S 0140-6736(23)00878-437805217 · doi ↗ · pubmed ↗
- 2Inskeep EK 2004 Preovulatory, postovulatory, and postmaternal recognition effects of concentrations of progesterone on embryonic survival in the cow 1,2J. Anim. Sci.82suppl_13E 24E 3910.2527/2004.8213_suppl E 24x 15471804 · doi ↗ · pubmed ↗
- 3Swanson AM David AL 2015 Animal models of fetal growth restriction: considerations for translational medicine Placenta 3662363010.1016/j.placenta.2015.03.00325819810 · doi ↗ · pubmed ↗
- 4Lammertink F Vinkers CH Tataranno ML Benders M 2020 Premature birth and developmental programming: mechanisms of resilience and vulnerability Front. Psychiatry 111153157110.3389/fpsyt.2020.53157133488409 PMC 7820177 · doi ↗ · pubmed ↗
- 5Ramel SE Gray HL Ode KL Younge N Georgieff MK Demerath EW 2011 Body composition changes in preterm infants following hospital discharge: comparison with term infants J. Pediatr. Gastroenterol. Nutr.5333333810.1097/MPG.0b 013e 3182243 aa 721602717 PMC 7680641 · doi ↗ · pubmed ↗
- 6Daskalakis G Psarris A Koutras A Fasoulakis Z Prokopakis I Varthaliti A 2023 Maternal infection and preterm birth: from molecular basis to clinical implications Children (Basel)1090710.3390/children 1005090737238455 PMC 10217143 · doi ↗ · pubmed ↗
- 7Takiishi T Fenero CIM Câmara NOS 2017 Intestinal barrier and gut microbiota: shaping our immune responses throughout life Tissue Barriers 5e 137320810.1080/21688370.2017.137320828956703 PMC 5788425 · doi ↗ · pubmed ↗
- 8Indrio F Neu J Pettoello-Mantovani M Marchese F Martini S Salatto A 2022 Development of the gastrointestinal tract in newborns as a challenge for an appropriate nutrition: a narrative review Nutrients 14140510.3390/nu 1407140535406018 PMC 9002905 · doi ↗ · pubmed ↗