Antimicrobial Activity of Actinomycetes Isolated From Hot Springs in Western Uganda
Joel Bazira, Karuhanga Naume Joyce, Nalumaga Pauline Petra, James Mwesigye, Kennedy Kassaza, Kawuma Simon

TL;DR
This study isolates Actinomycetes from Ugandan hot springs and finds that half of them show antimicrobial activity against harmful bacteria and fungi.
Contribution
The study identifies Actinomycetes from Ugandan hot springs as a new potential source of antimicrobial compounds.
Findings
Fifty percent of Actinomycetes isolates showed antimicrobial activity against at least one pathogen.
Isolates HB003 and HB026 were the most effective against Gram-negative and Gram-positive bacteria, respectively.
Genome analysis confirmed all isolates belonged to the Actinomycete species.
Abstract
Introduction: Actinomycetes are Gram-positive filamentous bacteria belonging to the phylum Actinobacteria. This study aimed to isolate and characterize Actinomycetes from Ugandan hot springs and evaluate their antimicrobial activity against pathogenic microorganisms. Materials and methods: A total of 30 Actinomycetes isolates were retrieved from two major hot springs in Uganda and identified using morphological and molecular techniques, including PCR amplification of the 16S rRNA gene and whole-genome sequencing. Antimicrobial activity was assessed against Staphylococcus aureus, Escherichia coli, and Candida species using cross-streak and agar well diffusion methods. Results: Out of the 30 Actinomycetes isolates, 15 (50%) exhibited antimicrobial activity against at least one of the test pathogens; six isolates (20%) exhibited antibacterial activity against Gram-negative bacteria, four…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
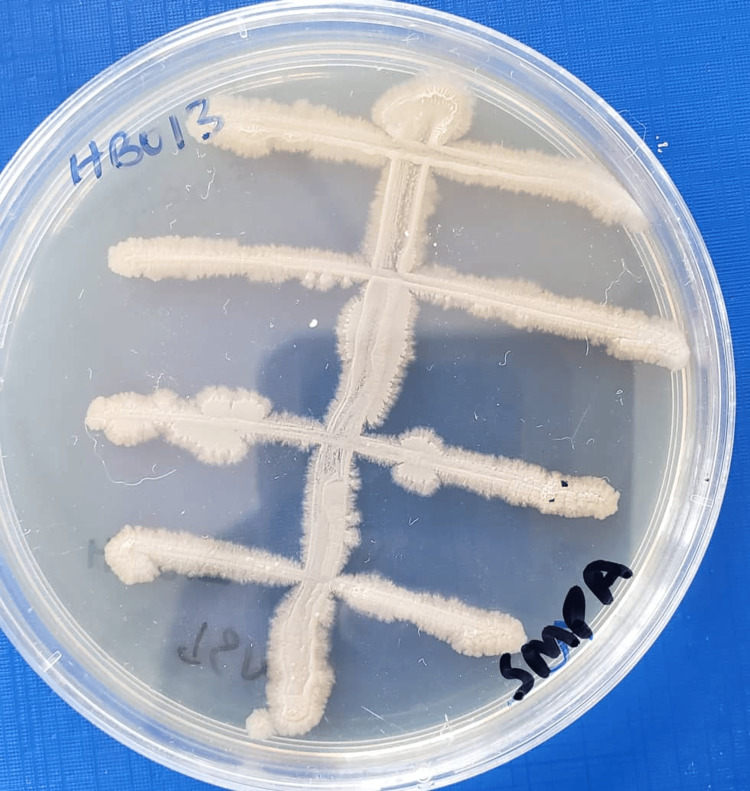 Figure 1
Figure 1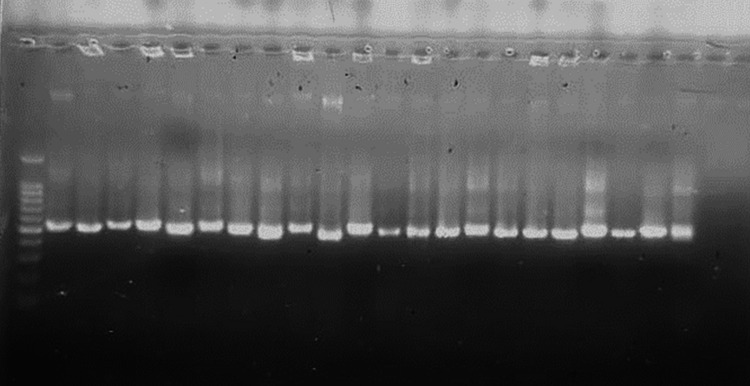 Figure 2
Figure 2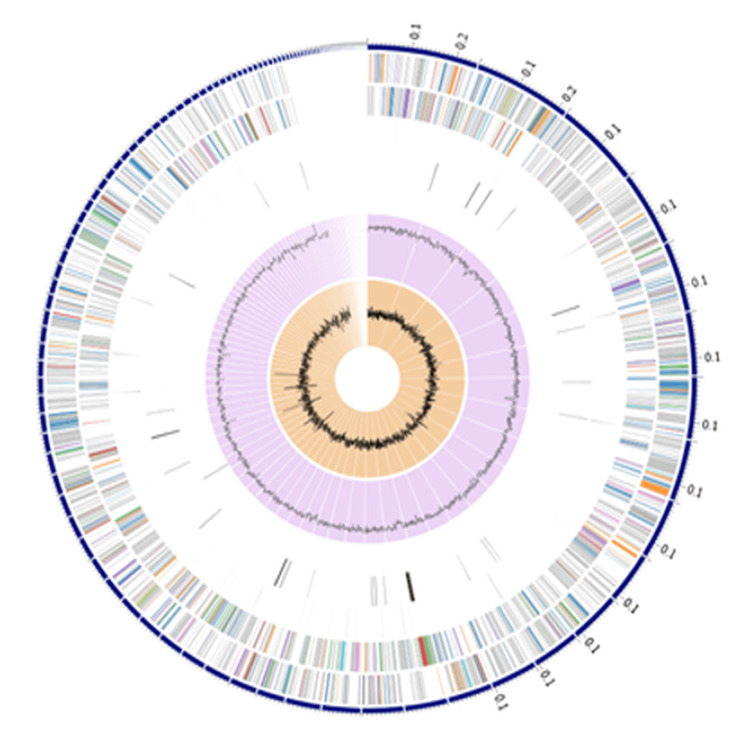 Figure 3
Figure 3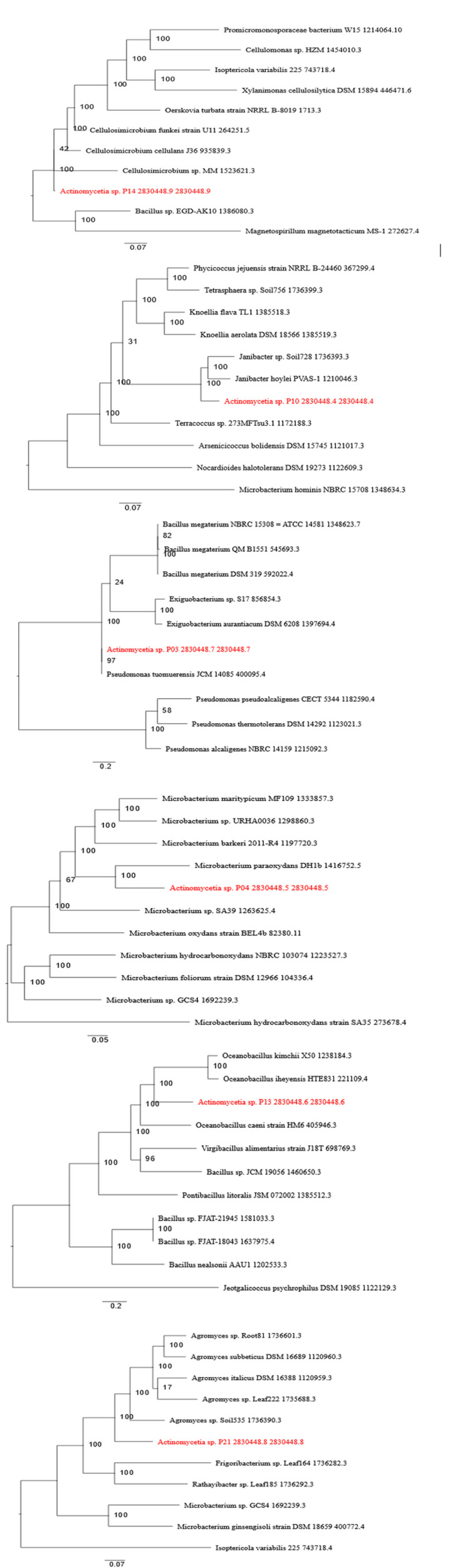 Figure 4
Figure 4| Sample | Source of Isolate | Color of Colonies | Mycelium Production |
| HKOO9-2 | Kabale Hot Spring | White | Yes |
| HB022 | Kitagata Hot Spring – Bushenyi | White | Yes |
| HB010 | Kitagata Hot Spring – Bushenyi | Yellow | Yes |
| HB030 | Kitagata Hot Spring – Bushenyi | White | No |
| HB013 | Kitagata Hot Spring – Bushenyi | White | Yes |
| HB012-2 | Kitagata Hot Spring – Bushenyi | Yellow | No |
| HB003 | Kitagata Hot Spring – Bushenyi | White | No |
| HB004 | Kitagata Hot Spring – Bushenyi | Orange | No |
| HB028-2 | Kitagata Hot Spring – Bushenyi | Yellow | No |
| HB028-1 | Kitagata Hot Spring – Bushenyi | Yellow | No |
| HK012 | Kabale Hot Spring | White | No |
| HB032 | Kitagata Hot Spring – Bushenyi | Orange | No |
| HB005 | Kitagata Hot Spring – Bushenyi | Orange | No |
| HB021 | Kitagata Hot Spring – Bushenyi | White | Yes |
| HK009-1 | Kabale Hot Spring | White | No |
| HB014 | Kitagata Hot Spring – Bushenyi | White | Yes |
| HKO10 | Kabale Hot Spring | Cream | No |
| HKOO8 | Kabale Hot Spring | White | Yes |
| HB016 | Kitagata Hot Spring – Bushenyi | Orange | No |
| HB009 | Kitagata Hot Spring – Bushenyi | Yellow | No |
| HB033 | Kitagata Hot Spring – Bushenyi | Orange | No |
| HB029-1 | Kitagata Hot Spring – Bushenyi | Cream | No |
| HB027 | Kitagata Hot Spring – Bushenyi | Orange | No |
| HB025 | Kitagata Hot Spring – Bushenyi | White | No |
| HB015 | Kitagata Hot Spring – Bushenyi | Orange | No |
| HB031 | Kitagata Hot Spring – Bushenyi | Yellow | No |
| HB024 | Kitagata Hot Spring – Bushenyi | White | No |
| HB007-1 | Kitagata Hot Spring – Bushenyi | Yellow | No |
| HB026 | Kitagata Hot Spring – Bushenyi | White | No |
| Total | - | 30 | 30 |
| Genome Features | Isolate Identification | |||
| P03 | P04 | P13 | P21 | |
| Contigs | 677 | 641 | 87 | 36 |
| GC content (%) | 53.94 | 57.37 | 37.6 | 72.28 |
| Contig L50 | 40 | 81 | 9 | 8 |
| Contig N50 | 114,697 | 28,400 | 173,000 | 147,808 |
| Genome length | 15,704,711 bp | 7,722,833 bp | 4,336,595 bp | 3,218,242 bp |
| Genome annotation | Actinomycetia sp. | Actinomycetia sp. | Actinomycetia sp. | Actinomycetia sp. |
| Unique genome identity | 2830448.7 | 2830448.5 | 2830448.6 | 2830448.8 |
| AMR Mechanism | Genes in Selected Isolates | |||||
| P10 | P14 | P03 | P04 | P13 | P21 | |
| Antibiotic activation enzyme | - | - | KatG | KatG | - | - |
| Antibiotic inactivation enzyme | AAC(3)-I | - | CatA family, FosB, Vgb(A) | APH(6)-Ia/APH(6)-Ib | FosB | - |
| Antibiotic target in susceptible species | Alr, Ddl, dxr, EF-G, EF-Tu, folA, Dfr, folP, gyrA, gyrB, inhA, fabI, Iso-tRNA, kasA, rho, rpoB, rpoC, S10p, S12p | Alr, Ddl, dxr, EF-G, EF-Tu, folA, Dfr, folP, gyrA, gyrB, inhA, fabI, Iso-tRNA, kasA, MurA, rho, rpoB, rpoC, S10p, S12p | Alr, Ddl, dxr, EF-G, EF-Tu, folA, Dfr, folP, gyrA, gyrB, inhA, fabI, Iso-tRNA, kasA, MurA, rho, rpoB, rpoC, S10p, S12p | Alr, Ddl, dxr, EF-G, EF-Tu, folA, Dfr, folP, gyrA, gyrB, inhA, fabI, Iso-tRNA, kasA, MurA, rho, rpoB, rpoC, S10p, S12p | Alr, Ddl, EF-G, EF-Tu, folA, Dfr, folP, gyrA, gyrB, inhA, fabI, Iso-tRNA, kasA, MurA, rho, rpoB, rpoC, S10p, S12p | Alr, Ddl, dxr, EF-G, EF-Tu, folA, Dfr, folP, gyrA, gyrB, Iso-tRNA, kasA, MurA, rho, rpoB, rpoC, S10p, S12p |
| Antibiotic target modifying enzyme | - | - | - | Erm(X) | RlmA(II) | - |
| Antibiotic target protection protein | - | - | BcrC | - | - | - |
| Efflux pump conferring antibiotic resistance | - | - | BceA, BceB, MdtABC-OMF, MdtABC-TolC, MexJK-OprM/OpmH, MexVW-OprM, TolC/OpmH | BceA, BceB, Cmx family, Tet(42) | BceA, BceB, YkkCD | - |
| Antibiotic target replacement protein | FabG, HtdX | FabG, HtdX | FabG, fabL, HtdX | fabL, FabL-like | - | FabL-like |
| Gene conferring resistance via absence | gidB | gidB | gidB | gidB | gidB | gidB |
| Protein altering cell wall charge conferring antibiotic resistance | GdpD, PgsA | GdpD, PgsA | GdpD, PgsA | GdpD, PgsA | GdpD, PgsA | GdpD, PgsA |
| Protein modulating permeability to antibiotic | - | - | OprD family, OprF | - | - | - |
| Regulator modulating expression of antibiotic resistance genes | MtrA, MtrB | MtrA, MtrB | BceR, BceS, LiaF, LiaR, LiaS, MtrA, MtrB, OxyR | BceR, BceS, LiaF, LiaR, LiaS, MtrA, MtrB, VanO-type | BceR, BceS, LiaF, LiaR, LiaS | LpqB, MtrA, MtrB |
| Isolate Identification Number |
| Extended-Spectrum Beta Lactamase-Producing |
| Methicillin-Resistant |
| Azole-Resistant |
| HK012 | + | + | + | + | + | + |
| HK009-2 | - | - | + | + | + | + |
| HBO28-2 | + | + | + | + | + | + |
| HB030 | + | + | + | + | + | + |
| HB013 | - | - | + | + | + | + |
| HB021-1 | - | - | - | - | + | + |
| HKOO9-1 | + | + | + | + | + | + |
| HB022 | - | - | - | - | + | + |
| HB028-1 | + | + | + | + | + | + |
| HK010 | + | + | + | + | + | + |
| HB003 | - | - | - | - | - | - |
| HB004 | + | + | + | + | + | + |
| HB005 | + | + | - | - | + | + |
| HB010 | - | - | + | + | + | + |
| HB032 | + | + | + | + | + | + |
| HB012-2 | + | + | + | + | + | + |
| HK008 | + | + | + | + | + | + |
| HBO21 | - | - | - | - | + | + |
| HB014 | - | - | + | + | + | + |
| HB026 | - | - | + | + | + | + |
| Identification Number | Isolate | Zone of Clearance (mm) | |||||
|
| Extended-spectrum beta-lactamase-producing |
| Methicillin-resistant |
| Azole-resistant | ||
| 1 | HK012 | 0 | 0 | 12 | 12 | 0 | 0 |
| 2 | HK009-2 | 12 | 10 | 0 | 0 | 12 | 12 |
| 3 | HBO28-2 | 15 | 15 | 12 | 12 | 12 | 12 |
| 4 | HB030 | 15 | 15 | 0 | 0 | 10 | 10 |
| 5 | HB021-1 | 0 | 0 | 19 | 19 | 0 | 0 |
| 6 | HB022 | 17 | 16 | 15 | 14 | 0 | 0 |
| 7 | HK010 | 10 | 10 | 0 | 0 | 0 | 0 |
| 8 | HB003 | 18 | 16 | 15 | 15 | 0 | 0 |
| 9 | HB004 | 0 | 0 | 12 | 12 | 0 | 0 |
| 10 | HB005 | 10 | 10 | 14 | 14 | 12 | 12 |
| 11 | HB032 | 0 | 0 | 18 | 14 | 0 | 0 |
| 12 | HB012-2 | 0 | 0 | 16 | 12 | 0 | 0 |
| 13 | HK008 | 12 | 10 | 0 | 0 | 0 | 0 |
| 14 | HB014 | 10 | 10 | 0 | 0 | 10 | 10 |
| 15 | HB026 | 0 | 0 | 18 | 16 | 14 | 14 |
| Ciprofloxacin | - | 30 mm | 8 mm | 30 mm | 8 mm | - | - |
| Fluconazole | - | - | - | - | - | 26 mm | 8 mm |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Natural Products and Biosynthesis · Antimicrobial agents and applications · Actinomycetales infections and treatment
Introduction
Actinomycetes is a genus of Gram-positive filamentous bacteria known for their filamentous growth pattern that belongs to the phylum Actinobacteria [1]. These bacteria are widely known for having the ability to release a variety of secondary metabolites, particularly antibiotics known to combat bacterial, fungal, and parasitic infections [2,3].
Antimicrobial resistance (AMR) is one of the most critical public health challenges of the 21st century, threatening effective treatment of infectious diseases globally [4]. The emergence of drug resistance and evolving multidrug-resistant (MDR) pathogenic bacteria has rendered the currently available antibiotics increasingly less effective against available drugs [3]. According to the World Health Organization (WHO), AMR is responsible for approximately 1.27 million deaths annually, with infections caused by resistant bacteria contributing to nearly five million deaths globally each year (4). The emergence of MDR and extensively drug-resistant (XDR) pathogens, such as Escherichia coli, Klebsiella pneumoniae, and methicillin-resistant *Staphylococcus aureus *(MRSA), has significantly reduced the efficacy of existing antibiotics, leading to prolonged hospital stays, increased healthcare costs, and higher mortality rates. Addressing this crisis requires exploration of novel environments and sources of bioactive compounds, particularly from microorganisms with remarkable metabolic capabilities such as Actinomycetes, which are prolific producers of antibiotics and other therapeutics [5,6].
Geothermal ecosystems, defined by high temperatures, severe pH levels, and mineral-rich conditions, provide a habitat for thermophilic and extremophilic microbes. This unique environment is thought to promote microbial adaptation and synthesis of metabolites with potential antimicrobial properties [7]. Actinomycetes have been discovered in various hot spring environments across the world, including those in Africa [8]. In Africa, hot springs are widely distributed across several countries, including Uganda, Kenya, Tanzania, and Ethiopia, as well as in all other regions with geothermal activity [9]. Hot springs throughout the world, such as in Iceland and China, have been studied and found to be repositories of microbial diversity and biosynthetic potential [10].
Despite growing global awareness of geothermal habitats as potential sources of antimicrobial compounds, Uganda's hot springs remain largely unexplored due to challenges such as logistical constraints in accessing remote geothermal sites, inadequate funding for microbiological and genomic studies, and a lack of advanced screening technologies for bioactive compounds. Therefore, the intention of this study was to isolate Actinomycetes from geothermal environments to evaluate antimicrobial activity against clinically relevant pathogens and explore the genes responsible for the production of antimicrobial compounds.
Materials and methods
Study location
This was an experimental, laboratory-based study conducted at the Department of Medical Microbiology of Mbarara University of Science and Technology (MUST), located in Southwestern Uganda, East Africa.
Laboratory methods
Sample Collection
A total of 30 *Actinomycetes *isolates, previously obtained from soil samples collected around the geothermal environments of Kitagata hot springs in Bushenyi District and Kabale hot springs in Southwestern Uganda, were stored at -70°C in the Microbiology Laboratory at MUST for six months and later retrieved following standard operating procedures. During primary isolation, the soil samples were pretreated following the method of Hayakawa et al. (1991) to suppress fast-growing bacteria and fungi [27]. Briefly, 1 g of soil was suspended in 10 mL of sterile saline, vortexed, and left to settle for 30 minutes. The suspension was then serially diluted, and 100 µL aliquots were plated onto starch casein agar. Plates were incubated at 37°C for five days, after which colonies with the characteristic chalky and filamentous morphology of Actinomycetes were picked, purified, and preserved at -70°C until further analysis.
Identification of Actinomycetes
Morphology Characterization of Actinomycetes
These isolates were thawed and sub-cultured on Starch M protein agar (HiMedia, Thane, India), a selective medium optimized for Actinomycetes growth and promotes colony formation [11], for seven days at 55°C. Morphological characterization of Actinomycetes isolates was done according to the ISP (International Streptomyces Project) recommendations [12]. Microscopic identification was done by Gram staining and observing under the 100× magnification of a light microscope.
Molecular Identification of Actinomycetes Isolates
DNA extraction: Genomic DNA was extracted using the cetyltrimethylammonium bromide (CTAB) method. DNA was purified using chloroform-isoamyl alcohol and precipitated with ethanol and stored at −20°C for further analysis.
Polymerase chain reaction (PCR) conditions: PCR amplification targeted the 16S rRNA gene using primers 63F (5′-CAG GCC TAA CAC ATG CAA GTC-3′) and 1378R (5′-GGG CGG WGT GTA CAA GGC-3′), yielding a 1300 bp product. PCR was performed in a BIORAD CFX 96 thermocycler under the following conditions: (i) initial denaturation at 95°C for five minutes, followed by cycling by 30 cycles at 95°C for one minute; (ii) annealing at 58°C for 40 seconds; and (iii) extension at 72°C for one minute and final extension at 72°C for 10 minutes. PCR products were visualized via gel electrophoresis (1% agarose gel) with a molecular ladder (1500 bp) to confirm the expected amplicon size.
Sequencing protocols: Amplicons were purified and sequenced using the Sanger method. The resulting sequences were compared against the NCBI GenBank database using BLAST to identify the Actinomycetes species.
Determination of antibacterial and antifungal activity of Actinomycetes
Primary Screening of Actinomycetes
Primary screening for the determination of antimicrobial activity was performed using the cross-streak method [13]. This technique was used to aid in identifying the Actinomycetes antimicrobial activity against standard strains of Escherichia coli ATCC 25922,* E. coli* resistant to extended-spectrum beta-lactamase (ESBL) enzyme harboring CTX, TEM, and SHV genes, Staphylococcus aureus ATCC 25904, MRSA strain, Candida albicans ATCC 10231, and azole-resistant Candida famata with the Erg 11 gene mutation. Starch M agar plates were prepared and inoculated with the pure culture of Actinomycetes by a single streak in the center of the Petri dish and incubated at 37°C for four days. The plates were inoculated with the test organisms by a perpendicular streak to the line of Actinomycetes growth. The plates were then incubated for 24 hours to determine zones of inhibition [14].
Secondary Screening of Actinomycetes
Secondary screening of Actinomycetes aided in analyzing the byproducts with antimicrobial products. Isolates demonstrating significant inhibition in primary screening were selected for secondary screening. Selection criteria included the presence and size of inhibition zones (≥10 mm in diameter). A pure culture of Actinomycetes grown on starch M agar plates for four days was inoculated into Soya Bean Casein Digest Broth, 15 mL in a 50 mL Falcon tube, using a sterile wire loop, and incubated for 24 hours. Fermentation was conducted in Soya Bean Casein Digest Broth (150 mL) at 28°C in a shaking incubator (300 rpm) for five days. Metabolites were extracted using ethyl acetate and dried to obtain crude extracts. Antimicrobial activity was evaluated using the agar well diffusion method [15].
Media Preparation and Incubation
Selective media such as Starch M protein agar and Soya Bean Casein Digest agar were prepared per the manufacturer's instructions. Starch M protein agar plates were used for initial culturing (5-10 days, 55°C), while fermentation utilized liquid broth in Erlenmeyer flasks incubated at 28°C with agitation. Indicator organisms for antimicrobial assays were cultured on Mueller-Hinton Agar for bacteria [16] and Sabouraud Dextrose Agar for the yeasts [17] at 37°C.
Quality Control
Positive controls included ciprofloxacin and fluconazole for bacterial and fungal assays, respectively. The results were interpreted according to the Clinical and Laboratory Standards Institute (CLSI) 2024 guidelines. Laboratory guidelines and protocols were used while performing all laboratory procedures.
Data management
All results obtained were double-checked for verification. Data were coded and entered into a pre-designed template in Microsoft Excel (Microsoft Corporation, Redmond, Washington) and then exported to STATA software (StataCorp. 2023. Stata Statistical Software: Release 18. College Station, TX: StataCorp LLC). The outcomes were presented in terms of percentages and discrete numbers.
Ethical consideration
The study proposal was submitted to the MUST Research Ethics Committee (REC) for approval. Permission to use the stored Streptomyces isolates was obtained from the principal investigator of the MUST-PES study.
Results
Isolation and identification of the possible Actinomycetes from Ugandan hot springs
Out of 30 isolates, 13 (43%) showed white colonies, eight (27%) yellow, seven (23%) orange, and two (7%) cream. The majority of the isolates (23, 77%) lacked mycelium production, while only seven (23%) showed mycelium production on selected culture media (Table 1, Figure 1).
Growth of isolate HB013 from Kitagata hot spring in Bushenyi
Molecular identification of Actinomycetes isolates
Genotypic Identification of the Isolates
PCR amplification of the 16S rRNA gene produced clear ~1300 bp bands in all isolates, as shown in Figure 2.
Electrophoresis gel image of Actinomycetes
Whole-Genome Sequencing
Whole-genome sequencing of selected isolates revealed high GC content (53.94%-74.55%) and identified various AMR genes. Isolate P03 harbored the highest number of AMR genes (145), including genes encoding efflux pumps and antibiotic-modifying enzymes. Phylogenetic analysis confirmed that all isolates were distinct members of the Actinomycetia class (Tables 2, 3) (Figures 3, 4). This enabled the identification of biosynthetic gene clusters (BGCs) associated with antimicrobial compound production, confirming the metabolic potential of the isolated Actinomycetes.
Graphical display of the distribution of genome annotations of sequenced strainsFrom outer to inner rings, contigs, coding sequences (CDS) on the forward strand, CDS on the reverse strand, RNA genes, CDS with antimicrobial resistance genes, CDS with virulence factors, GC content, and GC skew. The colors on CDS on the forward and reverse strands indicate the subsystem where the functionality proteins belong.Image created by the authors.
Evolutionary distance tree showing phylogenetic placement of sequenced isolatesAll the isolates exhibited >99% sequence homology with Acticetianomycetia sp.Image created by the authors.
Antimicrobial activity of the Actinomycetes isolated from Ugandan hot springs
Primary Screening
Primary screening using the cross-streak method revealed that 15/30 (50%) of the isolates showed antimicrobial activity. Four isolates demonstrated broad-spectrum activity against both Gram-positive and Gram-negative bacteria. Six isolates were active only against Gram-positive bacteria, while five isolates were active only against Gram-negative bacteria. Notably, isolate HB003 showed the highest activity against Gram-negative bacteria (zone of clearance: 18.0 mm), while HB026 exhibited the highest activity against Gram-positive bacteria (zone of clearance: 18.0 mm), suggesting these isolates as potential candidates for further microbiology studies (Table 4).
Secondary Screening
Antimicrobial activity was confirmed for all 15 isolates, with significant inhibition zones observed (p < 0.05) (Table 5). Activity against Candida species was observed in six isolates, with the largest zone of clearance (14.0 mm) recorded for isolate HB026.
Discussion
This study demonstrates the potential of Actinomycetes isolated from Ugandan hot springs as a source of novel antimicrobial compounds. Morphological, molecular, and genomic analyses confirmed that the isolates belong to the genus Actinomycetes, with some isolates exhibiting significant antibacterial and antifungal activities against clinically relevant pathogens. In reference to the standard positive controls, ciprofloxacin for antibacterial assays and fluconazole for antifungal assays, 15 out of 30 (50%) of the Actinomycetes isolates exhibited antimicrobial activity, suggesting a strong antimicrobial potential. This broad-spectrum activity highlights their value as promising compounds for further purification and compound identification. These findings contribute to the evidence supporting geothermal environments as reservoirs of bioactive microorganisms, aligning with similar findings in different geothermal sites worldwide [5,18-24].
The results revealed a diverse spectrum of microbial activity where 20% of isolates inhibited Gram-negative bacteria, another 13.3% inhibited Gram-positive bacteria, and 16.7% showed antifungal activity against Candida species. Particularly, isolates HB003 and HB026 exhibited strong antibacterial activity, with inhibition zones of 18.0 mm in diameter, comparable to and even exceeding those of resistant strains to standard antibiotics, such as ciprofloxacin. HB026 also demonstrated antifungal activity (14.0 mm) against Candida species, suggesting that some hot spring *Actinomycetes *may produce broad-spectrum antimicrobial compounds.
The GC content and abundance of AMR genes observed in our isolates are comparable to those reported in Actinomycetes from geothermal sites globally [25,26]. This genomic analysis of six selected isolates further strengthened our observations. High GC content (ranging from 53.94% to 74.55%) and the detection of BGCs and AMR genes confirm their metabolic versatility and potential to produce novel antibiotics. The presence of genes for efflux pumps, antibiotic-modifying enzymes, and target alteration suggests intrinsic defense mechanisms, possibly co-evolved in response to microbial competition in the geothermal environment, highlighting the complex genomic adaptation to extreme niches.
Hot springs are extreme environments with unique selective pressures that may drive the evolution of novel bioactive compounds not found in mesophilic microbes. Thus, isolates from such settings may be sources of new chemical compounds with activity against resistant microbes, including MRSA, ESBL-producing Enterobacteriaceae, and fluconazole-resistant *Candida *species, as used in this study.
However, unlike some studies employing species-level identification, our analysis focused on genus-level characterization, which limits direct strain comparisons.
Future studies should incorporate advanced molecular techniques such as metagenomics sequencing for comprehensive microbial profiling. Scaling up fermentation processes and isolating specific antimicrobial compounds through chromatographic techniques and mass spectrometry are critical next steps for harnessing the therapeutic potential of these isolates. Additionally, testing the efficacy of these compounds against a broader range of multidrug-resistant pathogens could validate their clinical relevance.
Conclusions
The geothermal hot springs of Uganda harbor thermophilic Actinomycetes with significant antimicrobial potential and novel drug sources to combat pathogenic microorganisms, addressing the global challenge of antimicrobial resistance. This observation indicates the untapped microbial resources with immense potential for drug discovery. Furthermore, the unique adaptations of these microorganisms to extreme conditions provide insights into novel biochemical pathways and secondary metabolites with antimicrobial properties.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1An update about plant growth promoting Streptomyces species Asian J Pharmacogn Al‑Tammar FK Khalifa AYZ 3240112023
- 2Streptomyces inside-out: a new perspective on the bacteria that provide us with antibiotics Philos Trans R Soc Lond B Biol Sci Chater KF 76176836120061662729310.1098/rstb.2005.1758 PMC 1609407 · doi ↗ · pubmed ↗
- 3How many antibiotics are produced by the genus Streptomyces?Arch Microbiol Watve MG Tickoo R Jog MM Bhole BD 38639017620011170208210.1007/s 002030100345 · doi ↗ · pubmed ↗
- 4Antimicrobial resistance Gastrointest Endosc Clin N Am Morrison L Zembower TR 6196353020203289122110.1016/j.giec.2020.06.004 · doi ↗ · pubmed ↗
- 5Investigation of antibacterial potential of Actinomycetales isolated from hot springs in India through a combined analysis involving in vitro and computational methods Chem Pap Kumari A Stany B Mishra S 115782024
- 6Genomic and metabolite profiling reveal a novel Streptomyces strain, QHH-9511, from the Qinghai-Tibet plateau Microbiol Spectr Feng XL Zhang RQ Wang DC 011202310.1128/spectrum.02764-22PMC 992749236622153 · doi ↗ · pubmed ↗
- 7Characterization and applications of Thermomonas hydrothermalis isolated from Jordan's hot springs for biotechnological and medical purposes Process Biochem Al-Daghistani HI Mohammad BT Kurniawan TA 1711811042021
- 8Going to extremes: progress in exploring new environments for novel antibiotics NPJ Antimicrob Resist Quinn GA Dyson PJ 8220243984350810.1038/s 44259-024-00025-8PMC 11721673 · doi ↗ · pubmed ↗