Breaking the silence—role of MucR as a virulence determinant in Brucella
Ian S. Barton, Connor B. Cribb, Beatriz Tartilán-Choya, Graham J. Bitzer, Jodi Ogle, Neil Garza Hernandez, Daniel W. Martin, Nieves Vizcaíno, Xindan Wang, Ilaria Baglivo, R. Martin Roop

TL;DR
This paper explores how the protein MucR controls virulence in Brucella bacteria by regulating gene expression during infection.
Contribution
The study identifies MucR as a key regulator of virulence genes in Brucella, linking its function to bacterial fitness during infection.
Findings
MucR regulates genes like those for the Type IV secretion system and adhesins BtaE and BmaC.
MucR ensures virulence gene expression aligns with bacterial fitness during infection.
MucR's role as a gene silencer is central to its function as a virulence determinant.
Abstract
The Zn finger protein MucR is an H-NS-like protein that serves as a gene silencer and nucleoid-structuring protein in the α-proteobacteria. MucR is also an essential virulence determinant in Brucella, where it directly and indirectly controls the expression of many genes required for the virulence of these bacteria in their mammalian hosts, including those encoding the Type IV secretion system and its effectors, the autotransporter adhesins BtaE and BmaC, and the quorum sensing regulators VjbR and BabR. Experimental evidence suggests that one of the primary functions of the Brucella MucR is to ensure that virulence genes are only expressed when their corresponding gene products provide fitness benefits to these bacteria during their infectious lifecycle, and we propose that this function is central to the well-established role of MucR as a virulence determinant.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
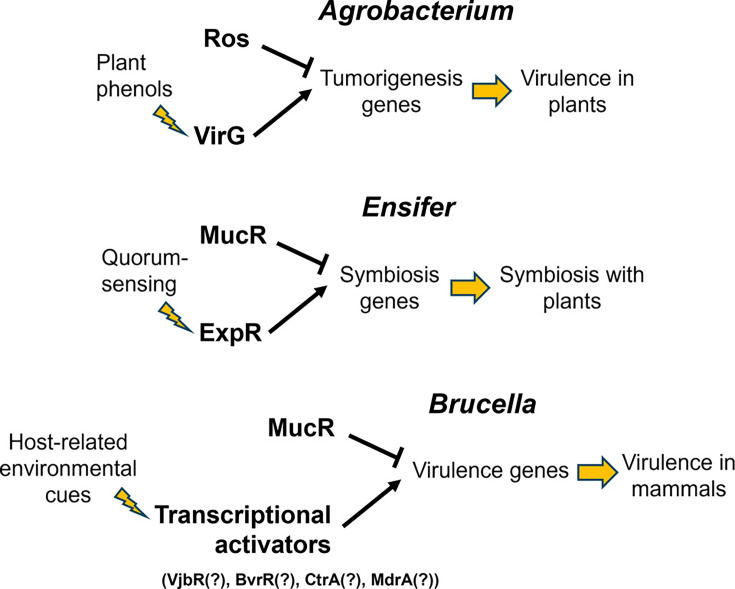 Fig 1
Fig 1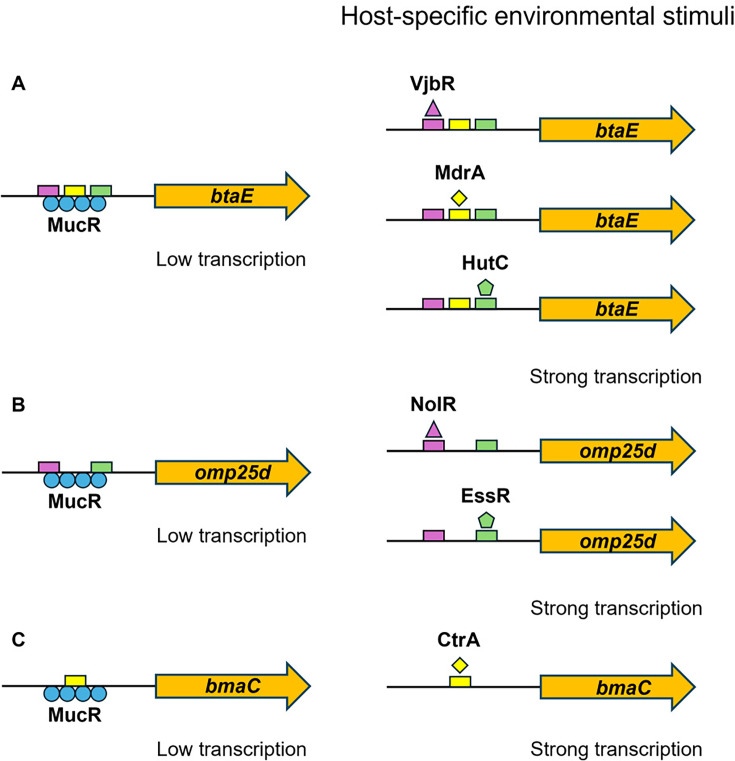 Fig 2
Fig 2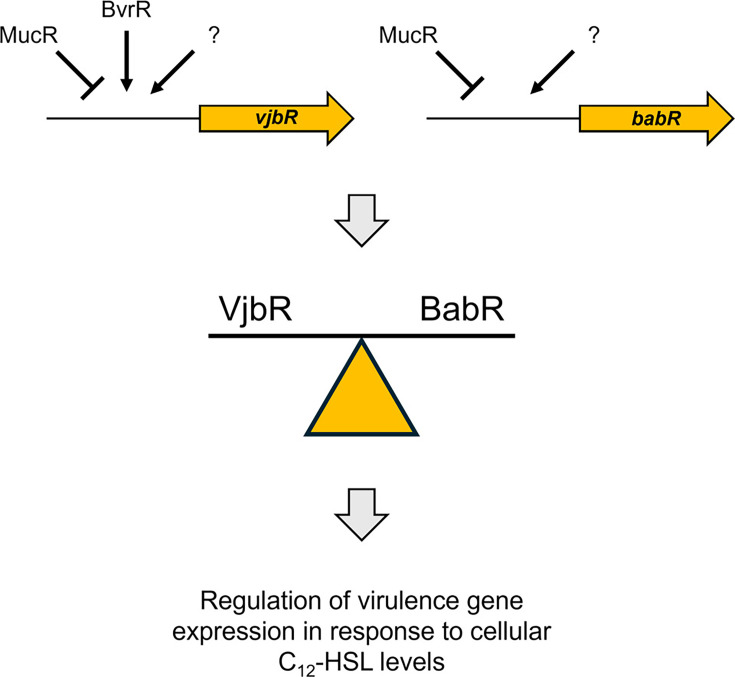 Fig 3
Fig 3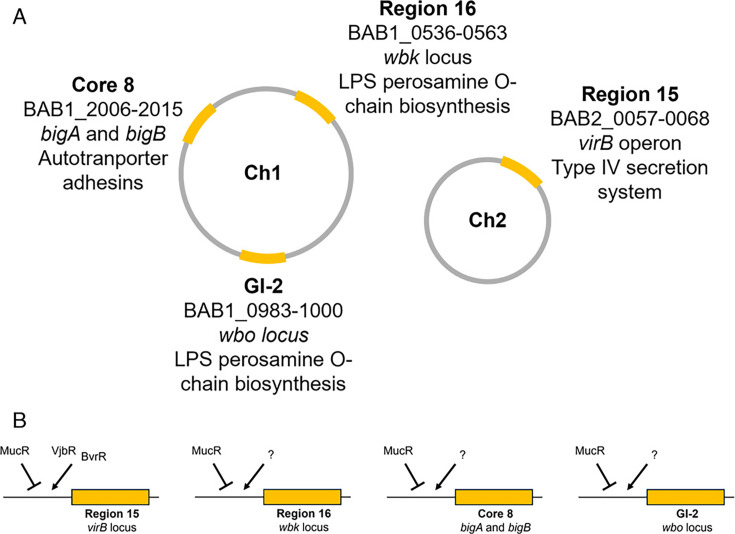 Fig 4
Fig 4| Gene | Possible counter-silencer(s) | Proposed function | References |
|---|---|---|---|
|
| VjbR, MdrA, HutC | Adhesin | ( |
|
| CtrA | Adhesin | ( |
|
| NolR, EssR | Adhesin | ( |
|
| VjbR, BvrR | T4SS | ( |
|
| VjbR | T4SS | ( |
|
| VjbR | T4SS effector | ( |
|
| VjbR, BvrR | T4SS effector | ( |
|
| VjbR | Acid shock protein | ( |
|
| CtrA | Cu/Zn SOD | ( |
|
| VjbR, CtrA | Unknown | ( |
|
| CtrA | Flagellin | ( |
|
| BvrR | Transcriptional regulator | ( |
|
| VjbR, CtrA | Transcriptional regulator | ( |
|
| VjbR, CtrA | Transcriptional regulator | ( |
- —National Institute of General Medical Scienceshttp://dx.doi.org/10.13039/100000057
- —National Science Foundationhttp://dx.doi.org/10.13039/501100008982
- —MCIN/AEI/10.13039/501100011033
- —National Institute of Allergy and Infectious Diseaseshttp://dx.doi.org/10.13039/100000060
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBrucella: diagnosis, epidemiology, treatment · Bacteriophages and microbial interactions · Escherichia coli research studies
MucR IS AN H-NS-LIKE GLOBAL REGULATOR IN THE α-PROTEOBACTERIA
MucR, also known as Ros or RosR, is a C_2_H_2_-type Zn finger protein that serves as a global regulator in the α-proteobacteria (1, 2). The gene encoding MucR was originally discovered during genetic studies aimed at identifying genes linked to the regulation of virulence traits in Agrobacterium (3) and exopolysaccharide biosynthesis in Rhizobium (now Ensifer) (4). The “ros” and “mucR” designations are based on the “rough outer surface” and “mucoid” colony phenotypes displayed by the Agrobacterium tumefaciens ros and Ensifer meliloti mucR mutants, respectively. MucR homologs control the expression of extensive regulons often encompassing hundreds of genes in many of the α-proteobacteria (5–10). Notably, some of these genes play critical roles in the symbiotic and pathogenic interactions of these bacteria with plants and animals. Despite its well-established role as a global regulator, the precise molecular mechanisms by which MucR exerts its regulatory functions remained unclear for many years. But recent studies have shown that MucR is a nucleoid-structuring protein that functions in a manner analogous to the Histone-like Nucleoid Structuring (H-NS) and H-NS-like proteins described in other bacteria (11). Specifically, MucR binds to low-consensus AT-rich regions across the bacterial genome, and its capacity to form higher-order oligomers folding in an unusual circular structure allows it to compact and structure the nucleoid and serve as a global transcriptional repressor or “gene silencer” (2, 11, 12). The gene silencing function of MucR has been proposed to work in concert with antagonistic “counter-silencers” to coordinate the proper temporal expression of cell cycle, symbiosis, and virulence genes in the α-proteobacteria (5, 8, 13) (Fig. 1) and protect these bacteria from the potentially toxic effects of uncontrolled expression of foreign genes acquired by horizontal gene transfer (HGT) (14).
Proposed roles of MucR homologs in coordinating the proper temporal expression of host-interaction genes in Agrobacterium tumefaciens (13), Ensifer (formerly Sinorhizobium) meliloti (5), and Brucella (11).
MucR IS AN ESSENTIAL VIRULENCE DETERMINANT IN BRUCELLA
Brucella strains are significant causes of abortion and infertility in food animals and sources of zoonotic infections worldwide (15). A link between MucR and Brucella virulence was first discovered during a screen of transposon mutants derived from Brucella melitensis 16M selected for attenuation in the J774.A1 murine macrophage-like cell line (16). Subsequent studies in several laboratories confirmed that B. melitensis mucR mutants are highly attenuated in cultured mammalian cells and mice (7, 16–18) and showed that Brucella abortus, Brucella canis, and Brucella ovis mucR mutants are likewise attenuated in these experimental models (6, 19, 20).
The nucleoid-structuring function of MucR allows this protein to have both direct and indirect impacts on gene expression by altering global DNA topology (11) and directly influencing the interactions of RNA polymerase and transcriptional regulators with genes. Transcriptomics and proteomics indicate that over 400 Brucella genes are directly or indirectly regulated by MucR (6, 7, 18–20), and ChIP-seq analysis in B. abortus 2308 suggests that this number may be even greater depending upon the environmental conditions under which gene expression is being evaluated (11). Many MucR-regulated genes are involved in basic physiology and metabolism, and Brucella mucR mutants typically display pleiotropic phenotypes when they are cultivated in vitro, including altered outer membrane properties, growth defects and a reduced capacity to respond to multiple environmental stresses including oxidative stress, acidic pH, and Fe deprivation. These generalized fitness defects almost certainly contribute to the attenuation exhibited in Brucella mucR mutants by compromising their ability to overcome the harsh environmental conditions within the intracellular environment of their mammalian hosts (21). However, as pointed out below, evidence suggests that MucR also plays very specialized roles in Brucella virulence.
MucR AND “COUNTER-SILENCERS” CONTROL VIRULENCE GENE EXPRESSION IN BRUCELLA
Some virulence determinants are only beneficial to bacterial pathogens during specific stages of their infectious life cycle. Adhesins, for example, allow bacteria to attach to and/or enter mammalian cells and often impart specificity for certain host tissues (22). Effectors secreted by Type IV secretion systems, on the other hand, typically exert their direct impacts during the intracellular replication of bacteria in host cells (23). Thus, it is not surprising that uncontrolled expression of certain virulence genes in bacterial pathogens has been shown to have a detrimental effect on the infectious process and lead to attenuation (24).
One of the major functions of the H-NS and H-NS-like gene silencers in bacterial pathogens is to prevent the uncontrolled and gratuitous expression of virulence genes (25). Transcriptional activators known as counter-silencers play an equally important role in ensuring the proper temporal expression of these genes by responding to host-specific environmental cues and overcoming this gene silencing. The concerted activities of these antagonistic gene silencer/counter-silencer pairs allow pathogens to ensure that genes encoding specific virulence determinants are selectively expressed at stages of the infectious process when they confer a fitness benefit (26, 27).
As noted in a previous section, MucR has been proposed to function as an H-NS-like gene silencer and work in concert with antagonistic transcriptional activators to coordinate the proper temporal expression of virulence genes in A. tumefaciens (13) and symbiosis genes in E. meliloti (5). Recent evidence indicates that MucR plays the same regulatory role in Brucella (Fig. 1). Table 1 lists the Brucella virulence genes for which there is evidence suggesting co-regulation by MucR and specific transcriptional activators. Notably, two of these transcriptional activators, VjbR and BvrR, control gene expression in response to host-specific environmental stimuli. VjbR is the predominant quorum-sensing regulator in Brucella (28–30) and BvrR and the corresponding sensor BvrS regulate gene expression in response to the acidic pH and nutrient deprivation encountered in the intracellular environment of mammalian cells (31). CtrA is also notable because it is the master regulator of the cell cycle in Brucella (32) and Brucella cells arrested in phase G1 of the cell cycle are the most infectious for host cells (33). Similarly, NolR, MdrA, and HutC have also been linked to the activation of important Brucella virulence genes (34–37).
ADHESINS
The best experimental evidence for the existence of MucR/counter-silencer pairs coordinating virulence gene expression in Brucella comes from ongoing studies of MucR regulation of the genes encoding BtaE and Omp25d. BtaE is a polar autotransporter adhesin that plays an important role in the attachment of B. suis and B. abortus strains to epithelial cells (22, 38). Omp25d is an outer membrane protein that is required for full virulence of B. suis in mice and may be involved in membrane homeostasis (37), but also affects the capacity of B. ovis to attach to and invade epithelial cells and macrophages (43), suggesting that Omp25d may play multiple roles in virulence across Brucella spp. Both proteins are produced at very low levels in Brucella cells during routine in vitro cultivation, suggesting that the corresponding genes are tightly regulated (20, 22). MucR binds directly to the btaE and omp25d promoters and strongly represses the expression of these genes (6, 11, 20). VjbR-, MdrA-, and HutC-binding sites overlap the MucR binding site in the btaE promoter in B. abortus 2308 (11, 38, 39), and MdrA can displace MucR from this promoter in an electrophoretic mobility shift assay (11). Similarly, a predicted NolR-binding site overlaps the MucR binding site in the omp25d promoter in B. abortus (11, 37). These experimental findings suggest that MucR and antagonistic counter-silencers can work together to coordinate the proper temporal expression of adhesins in Brucella in response to host-specific environmental stimuli (Fig. 2A and B) in much the same fashion that H-NS/counter-silencer pairs coordinate the temporal expression of adhesin genes in other pathogenic bacteria such as Yersinia enterocolitica (44) and Escherichia coli O157:H7 (27). It is also interesting to note that the gene encoding the unipolar autotransporter adhesin BmaC, which like BtaE is important for attachment to epithelial cells and is produced at low levels during routine in vitro cultivation of Brucella cells (45), is strongly repressed by MucR in B. abortus and has binding sites for the cell cycle regulator CtrA that overlap MucR binding sites in its promoter (11, 32). This suggests that MucR and CtrA may work in concert to coordinate bmaC expression in response to cell cycle signals (Fig. 2C), which would be consistent with the observation that Brucella cells in G1 of the cell cycle are more invasive for mammalian cells than those in other phases of the cell cycle (33).
Proposed roles of MucR and counter-silencers in regulating the proper temporal expression of the genes encoding BtaE (A), Omp25d (B), and BmaC (C) in Brucella. Under non-permissive conditions, MucR represses transcription of these genes (left) until host-specific environmental stimuli activate counter-silencers that displace MucR and induce expression (right).
QUORUM SENSING
Two LuxR-type regulators, VjbR and BabR (aka BlxR), control virulence gene expression in Brucella in response to cellular C_12_-HSL levels (28–30, 46, 47). These two regulators control the expression of many of the same genes, but experimental infections in mice and cultured mammalian cells have shown that VjbR has a more dramatic effect on virulence than BabR. MucR binds directly to the babR promoter in B. abortus 2308 and babR expression is elevated >20-fold in an isogenic mucR mutant compared to 2308 (6, 11). Elevated babR expression has also been reported in B. melitensis, B. ovis, and B. canis mucR mutants (7, 19, 20). As shown in Table 1, there is also evidence suggesting that vjbR expression may be coordinately regulated by MucR and BvrR (11). This raises the possibility that MucR regulation of babR and vjbR expression may play an important role in balancing the cellular levels of these two quorum-sensing regulators and influencing their differential impacts on virulence gene expression (Fig. 3).
Proposed role of MucR in balancing the cellular levels of the Brucella quorum-sensing regulators VjbR and BabR. MucR and transcriptional regulators (known and unknown) regulate the expression of vjbR and babR (top), which affects the relative cellular levels of VjbR and BabR (middle) that ultimately modulate virulence gene expression in response to cellular C12-HSL levels (bottom).
TYPE IV SECRETION
Multiple lines of evidence suggest that MucR regulates the genes encoding the Type IV secretion system in Brucella (7, 19, 20, 48). ChIP-seq and EMSA analysis have also demonstrated a strong binding affinity for MucR in the intergenic space between virB1 and virB2 in B. abortus (11) suggesting that MucR directly regulates these genes. However, as discussed in the previous section, MucR has a strong repressive effect on babR expression, and BabR regulates virB expression in B. melitensis (28, 47). Moreover, as shown in Table 1, evidence suggests that MucR and the transcriptional activators VjbR and BvrR may work in concert to coordinate the expression of the genes encoding the T4SS and its effectors in response to host-specific stimuli (11). This complex regulatory network could be envisioned to provide the brucellae with a mechanism for maximizing the expression of the T4SS genes at specific stages of infection when these genes provide their fitness benefits in much the same way that H-NS and H-NS-like proteins work in concert with antagonistic counter-silencers to coordinate the expression of the genes encoding the Type III secretion systems and their effectors in Salmonella (49) and Pseudomonas (50). But whether the MucR regulatory links to virB are essential for the wild-type virulence of Brucella strains remains to be experimentally determined.
MucR AND THE EVOLUTION OF BRUCELLA VIRULENCE
Like its H-NS and H-NS-like counterparts, MucR has been proposed to function as a xenogeneic silencer in the α-proteobacteria preventing the potentially toxic expression of genes acquired by HGT (51). Based on their intracellular niche and close association with their mammalian hosts, it is thought that the “classical” Brucella strains such as B. melitensis, B. suis, B. abortus, B. canis, and B. ovis have little opportunity to acquire novel genes by HGT (52). But it has been proposed that the acquisition of specific virulence genes such as those encoding the perosamine O-chain of the LPS, the T4SS, and the adhesins BigA and BigB from other bacteria by HGT played key roles in the evolution of these Brucella strains to become highly host-adapted mammalian pathogens (52–54), though recent examination of atypical Brucella spp. suggests that the acquisition, distribution, and role of HGT loci in the evolution of Brucella may be more complex (55). Consequently, it is interesting that the multi-gene genomic islands that encode the T4SS, LPS O-chain, and BigA and BigB adhesins in B. abortus 2308 are covered by broad MucR ChIP signatures (Fig. 4A) (11). Identifying the transcriptional activators that regulate the expression of these genes, defining the environmental stimuli that these regulators respond to, and determining how MucR and these regulators interact to coordinate the expression of these horizontally-acquired virulence genes (Fig. 4B) will give us a better picture of how MucR’s proposed role as a xenogeneic silencer has influenced the evolution of Brucella strains as mammalian pathogens.
(A) B. abortus 2308 genomic islands encoding known virulence determinants that are bound by MucR (11) including Region 16 (52), GI-2 (53), and Core 8 (52, 54) on Chromosome 1 that encode the wbk locus, wbo locus, and bigA/bigB, respectively, as well as Region 15 (52) on Chromosome 2 that encodes the virB operon (B) Model for regulation of known virulence determinants encoded on genomic islands by MucR and other transcriptional activators.
REGULATION OF MucR ACTIVITY
One aspect of the role of MucR in Brucella and related members of the α-proteobacteria that remains elusive is how the activity of MucR is regulated outside of interactions with functional transcriptional counter-silencers. While the transcription of mucR has been shown to be autoregulated in Ensifer (56), Agrobacterium (57), and Brucella (6), the effects of this regulation on MucR protein levels have not been thoroughly examined. In systems encoding H-NS and other H-NS-like proteins, post-translational mechanisms—including covalent modifications (e.g., phosphorylation and acetylation), targeted proteolysis, and interactions with binding partners and small molecules—regulate protein function and stability (58, 59). However, it is unclear whether similar regulatory mechanisms exist for MucR. In E. loti, one study failed to detect post-translational modifications of native MucR (60), but this was only characterized under a single growth condition and to our knowledge, post-translational modifications of MucR have not been examined in other members of the α-proteobacteria, including Brucella spp. Future experiments aimed at understanding the regulation of MucR activity will be important for understanding the complete role of MucR in the α-proteobacteria and Brucella virulence.
CONCLUSIONS
** **MucR’s function as a nucleoid-structuring protein, its impact on the expression of a broad array of genes, and the pleiotropic fitness defects displayed by Brucella mucR mutants undoubtedly play important roles in the dramatic attenuation displayed by mucR mutant strains (6, 7, 11, 19, 20). However, as detailed in previous sections of this manuscript, evidence also suggests that the capacity of MucR to function as a gene silencer allows this protein to work in concert with antagonistic counter-silencers to ensure that specific virulence-associated genes are only highly expressed when their gene products provide fitness benefits during the infectious life cycle (11). Preventing the gratuitous expression of these genes would minimize wasteful energy expenditure and, in theory, prevent unproductive interactions of the brucellae with specific host tissues. This proposed function is consistent with the role that H-NS and H-NS-like proteins play in coordinating the proper temporal expression of virulence genes in other bacterial pathogens (25, 61, 62). Moreover, from an evolutionary perspective, MucR has been proposed to play a similar role in coordinating the proper temporal expression of “host-interaction” genes in plant pathogens and symbionts that are close phylogenetic relatives of the brucellae (5, 13). Future studies will tell us just how important these functions are in MucR’s role as an essential virulence determinant in Brucella. Additionally, further elucidation of how MucR oligomerization influences its interaction with DNA will be critical, as emerging structural data suggest that these parameters may modulate MucR function in ways distinct from H-NS and other H-NS-like proteins (12).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Janczarek M. 2022. The Ros/Muc R zinc-finger protein family in bacteria: structure and functions. Int J Mol Sci 23:15536. doi:10.3390/ijms 23241553636555178 PMC 9779718 · doi ↗ · pubmed ↗
- 2Baglivo I, Malgieri G, Roop RM 2nd, Barton IS, Wang X, Russo V, Pirone L, Pedone EM, Pedone PV. 2025. Muc R protein: three decades of studies have led to the identification of a new H-NS-like protein. Mol Microbiol 123:154–167. doi:10.1111/mmi.1526138619026 PMC 11473720 · doi ↗ · pubmed ↗
- 3Close TJ, Tait RC, Kado CI. 1985. Regulation of Ti plasmid virulence genes by a chromosomal locus of Agrobacterium tumefaciens. J Bacteriol 164:774–781. doi:10.1128/jb.164.2.774-781.19854055699 PMC 214319 · doi ↗ · pubmed ↗
- 4Zhan HJ, Levery SB, Lee CC, Leigh JA. 1989. A second exopolysaccharide of Rhizobium meliloti strain SU 47 that can function in root nodule invasion. Proc Natl Acad Sci USA 86:3055–3059. doi:10.1073/pnas.86.9.30552717610 PMC 287063 · doi ↗ · pubmed ↗
- 5Mueller K, González JE. 2011. Complex regulation of symbiotic functions is coordinated by Muc R and quorum sensing in Sinorhizobium meliloti. J Bacteriol 193:485–496. doi:10.1128/JB.01129-1021057009 PMC 3019836 · doi ↗ · pubmed ↗
- 6Caswell CC, Elhassanny AEM, Planchin EE, Roux CM, Weeks-Gorospe JN, Ficht TA, Dunman PM, Roop RM 2nd. 2013. Diverse genetic regulon of the virulence-associated transcriptional regulator Muc R in Brucella abortus 2308. Infect Immun 81:1040–1051. doi:10.1128/IAI.01097-1223319565 PMC 3639602 · doi ↗ · pubmed ↗
- 7Dong H, Liu W, Peng X, Jing Z, Wu Q. 2013. The effects of Muc R on expression of type IV secretion system, quorum sensing system and stress responses in Brucella melitensis. Vet Microbiol 166:535–542. doi:10.1016/j.vetmic.2013.06.02323932078 · doi ↗ · pubmed ↗
- 8Fumeaux C, Radhakrishnan SK, Ardissone S, Théraulaz L, Frandi A, Martins D, Nesper J, Abel S, Jenal U, Viollier PH. 2014. Cell cycle transition from S-phase to G 1 in Caulobacter is mediated by ancestral virulence regulators. Nat Commun 5:4081. doi:10.1038/ncomms 508124939058 PMC 4083442 · doi ↗ · pubmed ↗