Methylation differences between assisted reproductive technology-conceived and naturally conceived children near BRCA1 and NBR2
Yunsung Lee, Håkon Kristian Gjessing, Christian Magnus Page, Jon Bohlin, Robert Lyle, Per Magnus, Siri Eldevik Håberg

TL;DR
Children conceived via assisted reproductive technology show subtle DNA methylation differences near BRCA1/NBR2 at birth, but these differences fade over time.
Contribution
This study provides longitudinal evidence on the persistence of ART-associated DNA methylation differences in children over time.
Findings
ART-conceived children showed subtle hypermethylation at the BRCA1/NBR2 promoter at birth.
The methylation differences diminished over time and were not statistically significant after correction.
The findings suggest a potential ART-associated epigenetic signature that may fade postnatally.
Abstract
Recent studies have shown that newborns conceived using assisted reproductive technology (ART) exhibited significantly different DNA methylation (DNAm) profiles at birth compared to those conceived naturally. Of note was the observation of increased DNAm at the promoter region of BRCA1/NBR2 in ART-conceived newborns. However, it remains unclear if these DNAm differences persist after birth. Using the Norwegian Mother, Father, and Child Cohort Study (MoBa), a large-scale population-based pregnancy cohort with extensive longitudinal data collected through biological samples and questionnaires, we generated longitudinal DNAm data for 105 ART-conceived and 250 naturally conceived children at birth and at ages 3–22 years. DNAm differences in the BRCA1/NBR2 promoter between ART- and naturally conceived children, at birth and postnatally, were tested using linear mixed model with adjustment…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
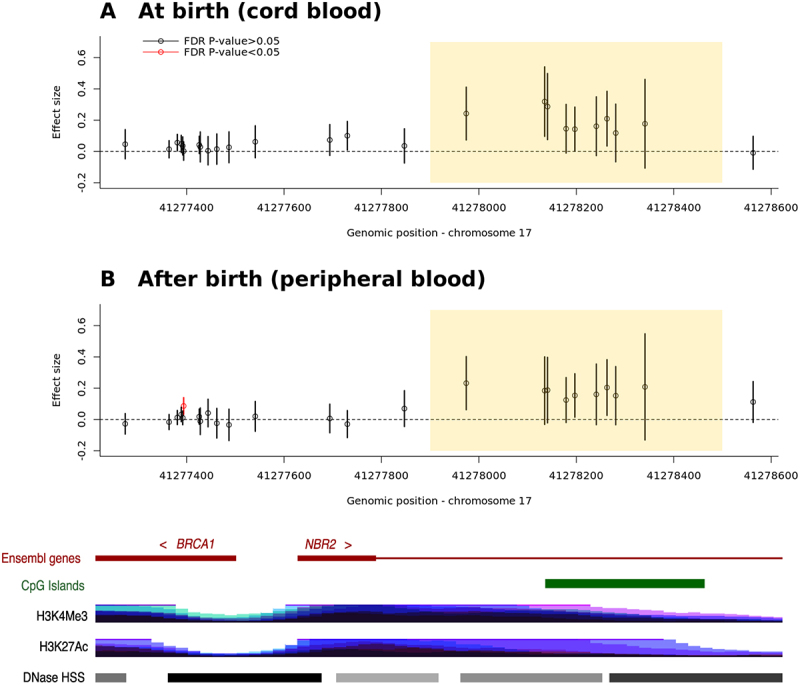 Figure 1
Figure 1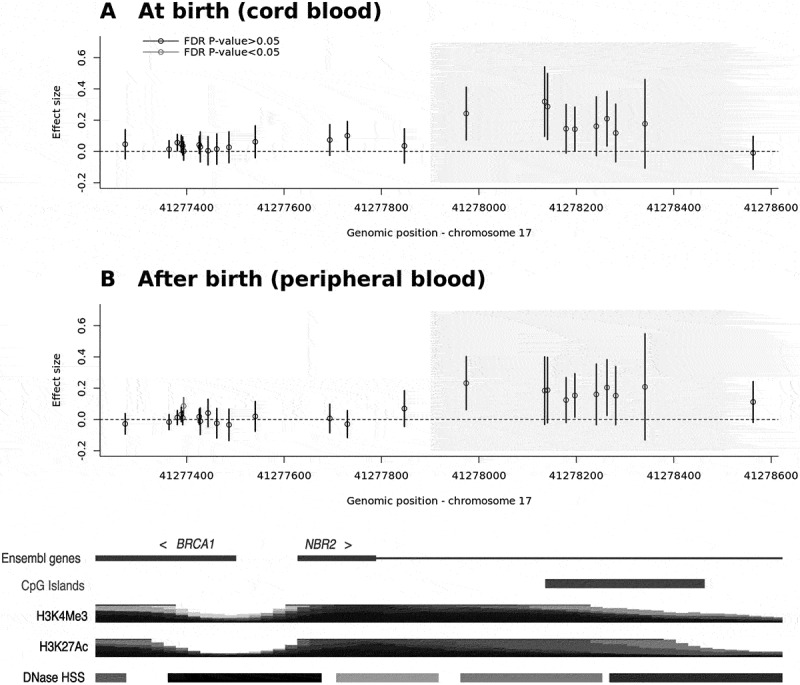 Figure 2
Figure 2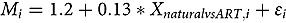 Figure 3
Figure 3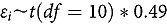 Figure 4
Figure 4- —Norwegian Cancer Society10.13039/100008730
- —Research council of Norway10.13039/501100005416
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBRCA gene mutations in cancer · Prenatal Screening and Diagnostics · Assisted Reproductive Technology and Twin Pregnancy
Introduction
A recent publication by Håberg, Page [1] presented the largest epigenome-wide association study of assisted reproductive technology (ART) to date, identifying 607 CpG sites that were differentially methylated in cord blood between newborns conceived using assisted reproductive technology (ART) and those conceived naturally. Among these differentially methylated loci, multiple CpGs in the bidirectional promoter region of BRCA1/NBR2 were found to be hypermethylated in ART-conceived newborns. Since BRCA1 is a well-established susceptibility gene for early-onset breast cancer and ovarian tumors [2,3], this finding may have important health implications in later life. Aberrant DNA methylation (DNAm) in the promoter region of this gene may alter gene expression patterns [4,5], and promoter methylation of this gene in peripheral blood has been associated with an increased risk of sporadic breast cancer [6,7] and ovarian cancer [8].
Despite these findings, it remains unknown whether the observed hypermethylation of BRCA1/NBR2 in ART-conceived children persists beyond birth or undergoes temporal changes during childhood as studies with longitudinal follow-up of DNAm changes are scarce. Yeung, Mendola [9] reported one differentially methylated locus (cg04061372 hg19 17:40,805,777) in ART-conceived (n = 23) and naturally conceived children (n = 93) at age 8–10 years, while reporting a null finding for BRCA1 promoter methylation. Similarly, Novakovic, Lewis [10] conducted an epigenome-wide association study of the mode of conception (ART vs natural) in adults aged 22–35 years (n = 233) but did not report any differentially methylated CpGs at BRCA1/NBR2.
To examine whether DNAm differences in the BRCA1 and NBR2 bidirectional promoter persisted beyond birth, we used a longitudinal design with repeated DNAm measurements, with one time point at birth and an additional time point between the age of 3 and 22 years, in the same children.
Methods
Study population
MoBa is a nationwide pregnancy cohort in which approximately 95,000 mothers, 75,000 fathers, and 114,000 children were recruited from 1999 to 2008 across Norway [11]. The participation rate of invited pregnant women was 41%. The participants completed a series of questionnaires during pregnancy and after childbirth and are still followed up. Peripheral whole-blood samples were collected from mothers and fathers around the 17^th^ week of gestation and from the mothers (whole-blood) and newborn children (cord blood) at delivery [12,13]. Various sub-studies in MoBa have collected peripheral blood samples, from different subsets of children between the age of 3 and 22 years [14,15].
For this study, we selected 119 ART-conceived and 250 naturally conceived control children matched on age, sex, and on the sub-study providing follow-up samples, with available cord blood at birth and peripheral blood at later age in the MoBa biobank. Among the ART-conceived children, there were 14 twin pairs. In contrast, among the naturally conceived children, there were no twin pairs. To ensure independence among children, we randomly selected one child from each twin pair in the ART group, resulting in 105 ART-conceived and 250 naturally conceived children for analyses (Table 1).Table 1.Descriptive statistics. Naturally-conceived (n = 250)ART-conceived (n = 105)Repeated measure of DNAm^1^ 2 time points243(97.2%)80(76.2%)1 time point7(2.8%)25(23.8%)NA0(0%)0(0%)Child’s sex Male102(40.8%)46(43.8%)Female148(59.2%)59(56.2%)NA0(0%)0(0%)Type of births^2^ Singleton248(99.2%)81(77.1%)Multiple (Twin)2(0.8%)24(22.9%)NA0(0%)0(0%)Maternal smoking during pregnancy No191(76.4%)91(86.7%)Sometimes47(18.8%)9(8.6%)Daily12(4.8%)5(4.8%)NA0(0%)0(0%)Maternal BMI before pregnancy Underweight7(2.8%)4(3.8%)Normal172(68.8%)69(65.7%)Overweight41(16.4%)22(21%)Obese16(6.4%)7(6.7%)NA14(5.6%)3(2.9%)Maternal parity 0125(50%)71(67.6%)182(32.8%)27(25.7%)236(14.4%)7(6.7%)35(2%)0(0%)42(0.8%)0(0%)NA0(0%)0(0%)Maternal age (in years) < 201(0.4%)0(0%)[20, 25)15(6%)0(0%)[25, 30)75(30%)14(13.3%)[30, 35)95(38%)42(40%)≥3564(25.6%)49(46.7%)NA0(0%)0(0%)^a^We measured DNAm levels at birth using cord blood samples and after birth (at age 3–22 years) using peripheral blood samples. Two time points were referred to as both at and after birth, while one time point was referred to as either of them.^b^This item shows the number of children from singleton or multiple births. Prior to generating this table, we randomly selected one child from each twin pair. As a result, in this table, the counts do not include both members of any twin ‘pair’ – each pair contributes only one child.
ART and covariates
ART procedures, including in vitro fertilization, intracytoplasmic sperm injection, and fresh or frozen-thawed embryo transfer, are reported by fertility clinics to the Medical Birth Registry of Norway (MBRN) which records all births nationwide. Using personal identification numbers, we could link the relevant information on ART procedures from MBRN to the MoBa children. Maternal smoking during pregnancy and body mass index (BMI) before pregnancy were obtained from the MoBa questionnaires, while parity, child’s sex, and child’s age were sourced from MBRN.
Measurement of DNA methylation
We retrieved DNA extracts from the selected children, collected at birth (cord blood samples) and after birth (at age 3–22 years, peripheral blood samples) that were stored at the MoBa biobank [12]. These samples were shipped to the Life & Brain Centre of the University of Bonn (Bonn, Germany) for generation of DNAm data using the Illumina Infinium MethylationEPIC v2.0 array.
Bisulphite conversion was performed using the EZ-96 DNA Methylation-Lightning MagPrep kit (Zymo Research, Irvine, California, USA). Raw iDat files were processed with the RnBeads v.2.21.3 R package in two batches, with random allocation to each batch. Sample-level quality control (QC) followed the RnBeads defaults with explicit thresholds: we visually inspected the fluorescence intensities from the control probes for all samples and predicted sex from X/Y probe intensities was compared with sex registered on MBRN; no mismatches were observed. To remove unreliable measurements, we applied the ‘greedycut’ algorithm with a detection P of 0.01 at both the probe and sample levels. Additionally, at the probe level, we excluded probes with low bead count (minimum coverage set to 5). The normal-exponential using out-of-band probes by SeSAMe [16] was used for background correction, and the beta-mixture quantile normalization [17] was used for normalization of Type I and II probe intensities. The QCed DNAm data included 869,783 autosomal CpGs, 22,618 X-chromosome CpGs, and 131 Y-chromosome CpGs.
From the QCed DNAm data, the cell-type composition was estimated using the UniLIFE reference panel [18] and EpiDISH R package [19], which is well suitable for longitudinal design including cord and peripheral blood samples. For longitudinal consistency, we applied the cord-blood reference (including nRBC) at birth and the adult peripheral-blood reference postnatally. Statistical analyses were performed on M-values [20].
Statistical analyses
First, we conducted association tests between the use of ART and DNAm in BRCA1/NBR2 at birth using DNAm data from the cord blood samples. We then performed the same association tests after birth using the data from the peripheral blood samples. For outlier removal, in each dataset, we identified and excluded children whose M-values fell outside the range: median (M-value) ± 5 × median absolute deviation. We applied a linear mixed model to regress each CpG on fixed effects for the use of ART, maternal age, smoking, BMI, parity, child’s sex, child’s age (only applicable to DNAm after birth), multiple pregnancy, and a random effect for the 96-well plate. As a sensitivity analysis, we included cell-type composition as additional covariates in the model. We also performed another sensitivity analysis by excluding children from multiple births.
Second, we modelled the within-individual change in DNAm according to the use of ART. To account for sex, age (only applicable to DNAm data after birth), and plate/batch effect, we applied a linear mixed model to regress each CpG on these variables separately in DNAm data at birth and those after birth. The residuals from these regressions were then standardized within each dataset to ensure comparability. Next, we computed the difference in standardized residuals between birth and later age for each CpG site, reflecting changes in DNAm over time. This difference was then used as the outcome variable in an ordinary linear model, where we examined its association with ART exposure while adjusting for maternal age, smoking status, BMI, and parity.
All the analyses were performed in R 4.1.2., and the linear mixed model was implemented using the lme function from the nlme R package. The two-sided Wald test was used to assess statistical significance. Relevant R code can be downloaded from https://github.com/yunsunglee-dev/BRCA-NBR2-methylation-in-ART-conceived-children.git.
Results
This study included a total of 250 naturally conceived children and 105 ART-conceived children (Table 1). For the 250 naturally conceived ones, we measured 247 individuals’ DNAm levels at birth in cord blood samples and 246 individuals’ DNAm after birth (at age 3–22 years) in peripheral blood samples (Figure S1, Supplemental File 1). For the 105 ART-conceived children, 81 individuals had available DNAm at birth, and 104 individuals had available DNAm after birth.
The distributions of the after-birth samples’ age were as follows: 81 individuals at age 0–5 years (naturally conceived n = 53 and ART n = 28), 21 individuals at 5–10 years (naturally conceived n = 19 and ART n = 2), 198 individuals at 10–15 years (naturally conceived n = 137 and ART n = 61), 43 individuals at 15–20 years (naturally conceived n = 33 and ART n = 10), and 7 individuals at 20–25 years (naturally conceived n = 4 and ART n = 3). These are visualized in Figure S1, Supplemental File 1.
The mothers of the ART-conceived children (33.8 years old) were, on average, older than those of the naturally conceived children (31.3 years old). The difference in maternal age was statistically significant (P-value = 8.5E-07).
We first examined the DNAm-ART association at birth, focusing on 25 CpGs located within the BRCA1/NBR2 promoter (hg19 17:41,277,059–41,278,712) previously reported by Håberg, Page [1]. As expected, a group of CpGs at the BRCA1/NBR2 promoter showed moderate hypermethylation in ART-conceived newborns as compared to naturally conceived ones (highlighted in yellow in Figure 1(A)). Although these DNAm-ART associations did not reach statistically significance after multiple testing correction, e.g., false discovery rate (FDR) <5%, the overall trend was similar to the previous findings (Figure 7(C) in Håberg, Page [1]). Figure 1.Dnam differences at BRCA1 and NBR2 in ART- and naturally conceived children at and after birth.The DNAm differences between ART- and naturally conceived children are presented as effect sizes on the y-axis. The effect sizes are shown (A) at birth (cord blood) and (B) after birth (peripheral blood, at age 3–22 years). Each dot represents the effect size of a DNAm-ART association at a CpG site, with black indicating FDR-adjusted p-value >0.05 and red indicating FDR-adjusted p-value <0.05. The yellow-highlighted region marks the genomic region where Håberg, Page [1] found hypermethylation in ART-conceived children compared to naturally conceived ones.
Next, we investigated whether these DNAm-ART associations identified above, persisted after birth by analyzing DNAm data from the same individuals at the age of 3–22 years. Again, we observed a similar trend with slight hypermethylation at the same CpG sites in ART-conceived children, although it did not reach statistical significance (Figure 1(B) and Table S1).
As a sensitivity analysis, we adjusted for estimated cell-type composition, which did not alter the results (Figure S2, Supplemental File 1). We also excluded the children from multiple births and performed the same analysis. The results remained unchanged (Figure S3, Supplemental File 1).
To assess whether DNAm-ART associations at birth differed from those after birth, we leveraged the longitudinal design of this study with repeated DNAm measurement in same individuals. Specifically, we calculated within-individual differences in DNAm level at each CpG between birth and later age and examined their associations with ART. We found no statistically significant associations between ART and the change in DNAm levels over time, indicating that there is no clear evidence that hypermethylation in ART-conceived children disappears after birth (Table 2).Table 2.Association between ART and within-child changes in DNAm at BRCA1 and NBR2 CpG sites.CpG siteCHR^1^Position^1^RegulatoryFeature Group^2^Relation to UCSCCpG Island^2^UCSC RefGeneName^2^ Beta coefficient^3^(95% confidence interval)RawP-valueFDR-correctedP-value^4^cg083868861741,277,274Promoter AssociatedN ShoreBRCA1;NBR2−0.067 (−0.184, 0.049)0.2580.785cg248069531741,277,364Promoter AssociatedN ShoreBRCA1;NBR2−0.016 (−0.085, 0.054)0.6610.787cg201872501741,277,381Promoter AssociatedN ShoreBRCA1;NBR2−0.038 (−0.108, 0.032)0.2870.785cg154192951741,277,389Promoter AssociatedN ShoreBRCA1;NBR2−0.02 (−0.094, 0.053)0.5870.787cg169630621741,277,392Promoter AssociatedN ShoreBRCA1;NBR2−0.019 (−0.092, 0.054)0.6090.787cg166309821741,277,394Promoter AssociatedN ShoreBRCA1;NBR20.088 (0.012, 0.165)0.0250.488cg212539661741,277,426Promoter AssociatedN ShoreBRCA1;NBR2−0.023 (−0.096, 0.051)0.5470.787cg041104211741,277,428Promoter AssociatedN ShoreBRCA1;NBR2−0.056 (−0.187, 0.076)0.4080.787cg046583541741,277,444Promoter AssociatedN ShoreBRCA1;NBR20.065 (−0.064, 0.194)0.3260.785cg173012891741,277,462Promoter AssociatedN ShoreBRCA1;NBR20.002 (−0.136, 0.14)0.9760.976cg094419661741,277,487Promoter AssociatedN ShoreBRCA1;NBR2−0.045 (−0.188, 0.099)0.5440.787cg268915761741,277,541Promoter AssociatedN ShoreBRCA1;NBR2−0.031 (−0.171, 0.108)0.6600.787cg108930071741,277,694Promoter AssociatedN ShoreBRCA1;NBR2−0.079 (−0.207, 0.049)0.2290.785cg121824521741,277,730Promoter AssociatedN ShoreBRCA1;NBR2−0.088 (−0.213, 0.037)0.1700.785cg098310101741,277,847Promoter AssociatedN ShoreBRCA10.073 (−0.063, 0.209)0.2940.785cg250671621741,277,974Promoter AssociatedN ShoreBRCA1−0.02 (−0.105, 0.064)0.6360.787cg262762331741,278,135 IslandBRCA1−0.119 (−0.231, −0.006)0.0390.488cg060017161741,278,141 IslandBRCA1−0.114 (−0.241, 0.013)0.0800.664cg022865331741,278,179 IslandBRCA1−0.034 (−0.114, 0.047)0.4150.787cg149472181741,278,197 IslandBRCA10.009 (−0.06, 0.079)0.7950.903cg160060041741,278,241 IslandBRCA1−0.007 (−0.113, 0.099)0.8990.976cg183722081741,278,263 IslandBRCA1−0.052 (−0.161, 0.056)0.3450.785cg146874741741,278,281 IslandBRCA10.003 (−0.092, 0.098)0.9500.976cg252881401741,278,341 IslandBRCA10.043 (−0.138, 0.223)0.6430.787cg275817621741,278,563 S ShoreBRCA10.1 (−0.041, 0.241)0.1650.785^a^Based on Genome Reference Consortium Human Build 37.^b^Retrieved from the Illumina Infinium MethylationEPIC v2.0 manifest file.^c^Derived from the regression of within-child difference in DNAm between at and after birth on ART and covariates.^d^The Benjamini-Hochberg procedure was applied.
We visually inspected scatter plots of the DNAm levels at CpGs against child’s age (Figure S4, Supplemental File 1). Among the CpGs located in the genomic region where Håberg, Page [1] found hypermethylation in ART-conceived children, we observed a tendency of differences in M-values between ART-conceived and naturally conceived children diminishing as child’s age increased. However, this interpretation was based on visual inspection, not based on statistical test such as the within-child difference model mentioned above.
Discussion
We found subtle hypermethylation at the BRCA1/NBR2 promoter in ART-conceived children compared to naturally conceived children both at birth and at later ages up to 22 years. Hypermethylation in the promoter region was observed at the same CpGs at birth and later ages. Though hypermethylation at later ages was slightly reduced compared to that at birth, the difference in the intensity of hypermethylation between at birth and at later ages was not statistically significant.
Several previous studies explored epigenetic differences between ART-conceived and naturally conceived children using different types of tissues, e.g., cord blood, placenta, and peripheral blood [21]. Among these studies, only one study by Håberg, Page [1] identified hypermethylation in ART-conceived children at BRCA1/NBR2. Although this finding has not yet been replicated, we consider it unlikely to be false positive, given the study’s large sample size (approximately 1,000 ART and 1,000 naturally conceived), control for parental DNAm, stringent multiple testing threshold (FDR < 1%), and the observation of increased DNAm at 10 consecutive CpGs. Two other studies that analyzed peripheral blood samples from children or adolescents, Yeung, Mendola [9] and Penova-Veselinovic, Melton [22], did not report statistically significant hypermethylation of BRCA1/NBR2 in ART-conceived individuals.
The BRCA1 promoter is normally unmethylated in peripheral blood [23], resulting in an unsilenced promoter and intact tumor-suppressor activity. Rarely, however, constitutional BRCA1 promoter methylation (occurring in non-tumor tissues such as peripheral blood) has been linked to increased risk of early onset breast cancer and ovarian cancer [7,24,25]. Although we found no strong evidence of the BRCA1/NBR2 promoter hypermethylation in ART-conceived children at the ages studied, it is still plausible that ART-conceived children show hypermethylation in their later life. McCartney, Zhang [26] reported two promoter CpGs, i.e., cg08386886 and cg26891576, showing modest correlation with chronological age, and peripheral blood methylation at these two CpGs together with their neighboring CpGs have been associated with triple-negative breast cancers and high-grade serous ovarian cancers (see Lonning, Nikolaienko [24]; CpG17 and CpG32 listed in eTable 1).
Furthermore, a recent study by Ruiz-Arenas, Hernandez-Ferrer [4] reported that increased methylation at cg25067162, located ~150 bp proximal to the CpG island, significantly reduced the expression levels of NBR2 (TC17000547.hg.1, fold change (FC) = 0.93, P-value = 3.61E-42) and TC17002214.hg.1 (FC = 0.92, P-value = 2.93E-43) in children’s peripheral blood. NBR2, a long non-coding RNA, has been suggested to be a regulatory element in cancer biology [27]. Taken together, increased DNAm in this region may have functional consequences.
We excluded outliers to secure the stability of linear mixed models. In such models, coefficient estimates – here, the differences in methylation between naturally-conceived and ART-conceived children – can be largely influenced by a handful of ‘technical’ outliers. Nevertheless, we do not rule out the possibility that outliers were ‘biologically’ driven [28,29]. For example, Ghosh, Mainigi [30] reported that children with low birthweight showed more methylation outliers than those with high birthweight, and among the children with low birthweight, those conceived in-vitro had more outliers in methylation than those conceived in-vivo. However, we did not observe a similar pattern in our data. In the peripheral blood (after-birth) methylation data, cg04110421 had the highest number of outliers (n = 11, all positive), with seven from naturally conceived children and four from ART-conceived children. Given the sample sizes (246 naturally conceived and 104 ART-conceived), this distribution of outliers was within expectation. Similar trends were observed across other CpGs, suggesting that outliers were unlikely to influence our findings.
This study has several limitations. First, the sample size limited our ability to detect small DNAm differences with statistical significance. According to our post-hoc power analysis of the cord blood DNAm data, 4.5-folds increase in sample size (1,112 naturally conceived and 365 ART-conceived) would be required to achieve 80% power in detecting the methylation difference at cg02286533. This estimate was based on a Monte-Carlo simulation with 10,000 iterations that assumed a true methylation difference of 0.13 in M-values (derived from Håberg, Page [1]), a standard deviation of 0.49 in both groups’ M-values (also derived from Håberg, Page [1]). To simulate slight heavy tails, a student t distribution with 10 degrees of freedom was used. In mathematical terms, the simulation model was , where . The statistical significance was defined conservatively as p < 0.05/25 = 0.002 (Bonferroni correction). For the peripheral blood DNAm data, we would have the confidence intervals and p-values included in Figure 1(B) reflect the statistical power as no prior study exists to inform us of key parameters for power calculation. Second, our study population consisted of individuals of Northern European ancestry, which may limit the generalizability of our findings to more diverse populations. Future studies with larger and more diverse cohorts are needed to validate and expand upon our results. Finally, this study did not expand its scope to the genome-wide level but focused on the BRCA1/NBR2 region. This decision was made a priori, as the study was conceived as a hypothesis-driven evaluation of this locus based on its established clinical relevance to cancer susceptibility and prior ART-related methylation findings by Håberg, Page [1]. In addition, the limited availability of postnatal peripheral blood samples in MoBa would have substantially reduced the statistical power for a discovery-scale epigenome-wide analysis, increasing the risk of generating underpowered or misleading results.
In conclusion, our findings suggest that subtle and persistent hypermethylation at the BRCA1/NBR2 promoter in ART-conceived children may reflect an epigenetic alteration with potential functional relevance, warranting further investigation in larger and more diverse populations.
Supplementary Material
Supplemental_File1.docx
S_Table1.xlsx
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Håberg SE, Page CM, Lee Y, et al. Dna methylation in newborns conceived by assisted reproductive technology. Nat Commun. 2022;13(1):1896. doi: 10.1038/s 41467-022-29540-w 35393427 PMC 8989983 · doi ↗ · pubmed ↗
- 2Miki Y, Swensen J, Shattuck-Eidens D, et al. A strong candidate for the breast and ovarian cancer susceptibility gene BRCA 1. Science. 1994;266(5182):66–9. doi: 10.1126/science.75459547545954 · doi ↗ · pubmed ↗
- 3Esteller M, Silva JM, Dominguez G, et al. Promoter hypermethylation and BRCA 1 inactivation in sporadic breast and ovarian tumors. J Natl Cancer Inst. 2000;92(7):564–569.10749912 10.1093/jnci/92.7.564 · doi ↗ · pubmed ↗
- 4Ruiz-Arenas C, Hernandez-Ferrer C, Vives-Usano M, et al. Identification of autosomal cis expression quantitative trait methylation (cis e QT Ms) in children’s blood. Elife. 2022;11:11. doi: 10.7554/e Life.65310 PMC 893300435302492 · doi ↗ · pubmed ↗
- 5Kumar M, Sahu RK, Goyal A, et al. Brca 1 promoter methylation and expression - associations with ER+, PR+ and HER 2+ subtypes of breast carcinoma. Asian Pac J Cancer Prev. 2017;18(12):3293–3299. doi: 10.22034/APJCP.2017.18.12.329329286222 PMC 5980886 · doi ↗ · pubmed ↗
- 6Bosviel R, Garcia S, Lavediaux G, et al. Brca 1 promoter methylation in peripheral blood DNA was identified in sporadic breast cancer and controls. Cancer Epidemiol. 2012;36(3):e 177–82. doi: 10.1016/j.canep.2012.02.00122402307 · doi ↗ · pubmed ↗
- 7Iwamoto T, Yamamoto N, Taguchi T, et al. Brca 1 promoter methylation in peripheral blood cells is associated with increased risk of breast cancer with BRCA 1 promoter methylation. Breast Cancer Res Treat. 2011;129(1):69–77. doi: 10.1007/s 10549-010-1188-120882403 · doi ↗ · pubmed ↗
- 8Lonning PE, Berge EO, Bjornslett M, et al. White blood cell BRCA 1 promoter methylation status and ovarian cancer risk. Ann Intern Med. 2018;168(5):326–334. doi: 10.7326/M 17-010129335712 · doi ↗ · pubmed ↗