Acinetobacter zhairhuonensis sp. nov., isolated from sediments of the East China Sea
Qian Jiang, Yike Wang, Kaiwen Zheng, Yuangui Tang, Yunlu Cui, Qunjian Yin, Daoqiong Zheng, Jinzhong Xu, Daxiong Ji, Dongdong Zhang, Sanjit Chandra Debnath, Pinmei Wang

TL;DR
A new species of bacteria, Acinetobacter zhairhuonensis, was discovered in the East China Sea and characterized through genomic and phenotypic analysis.
Contribution
The discovery and formal classification of a new Acinetobacter species based on comprehensive genomic and biochemical data.
Findings
The new species is distinct from other Acinetobacter species based on genomic thresholds like ANI, AAI, and dDDH.
The genome includes a 3.57 Mb chromosome and a plasmid with 41.3 mol% GC content.
Phenotypic traits and lipid profiles further support the classification of this novel species.
Abstract
A novel aerobic bacterial strain, designated as Acinetobacter zhairhuonensis sp. nov. A7.4T, was isolated from East China Sea sediments near Zhairuo island. Cells were Gram-stain-negative, non-motile short rods, forming milky, moist colonies. Growth occurred at 10–35 °C, pH 6.0–9.0 and 0–5% NaCl concentrations (w/v). Strain A7.4T was catalase-positive and oxidase-negative. Major fatty acids (>10%) included summed feature 3 (C16:1 ω7c/C16 :1 ω6c), C18:1 ω9c and C16:0. The polar lipid profile comprised diphosphatidylglycerol, phosphatidylglycerol, phosphatidylethanolamine, two phospholipids and two aminolipids. Ubiquinone Q-9 and Q-8 predominated. The genome (3.57 Mb, 41.3 mol% GC) included one chromosome and a plasmid. The core-genome phylogeny placed A7.4T within Acinetobacter, forming a distinct clade with Acinetobacter kanungonis PS-1T, Acinetobacter tibetensis Y-23T and Acinetobacter…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
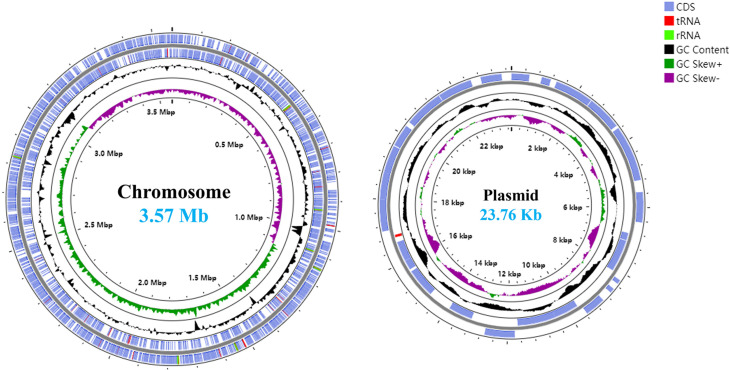 Fig. 1
Fig. 1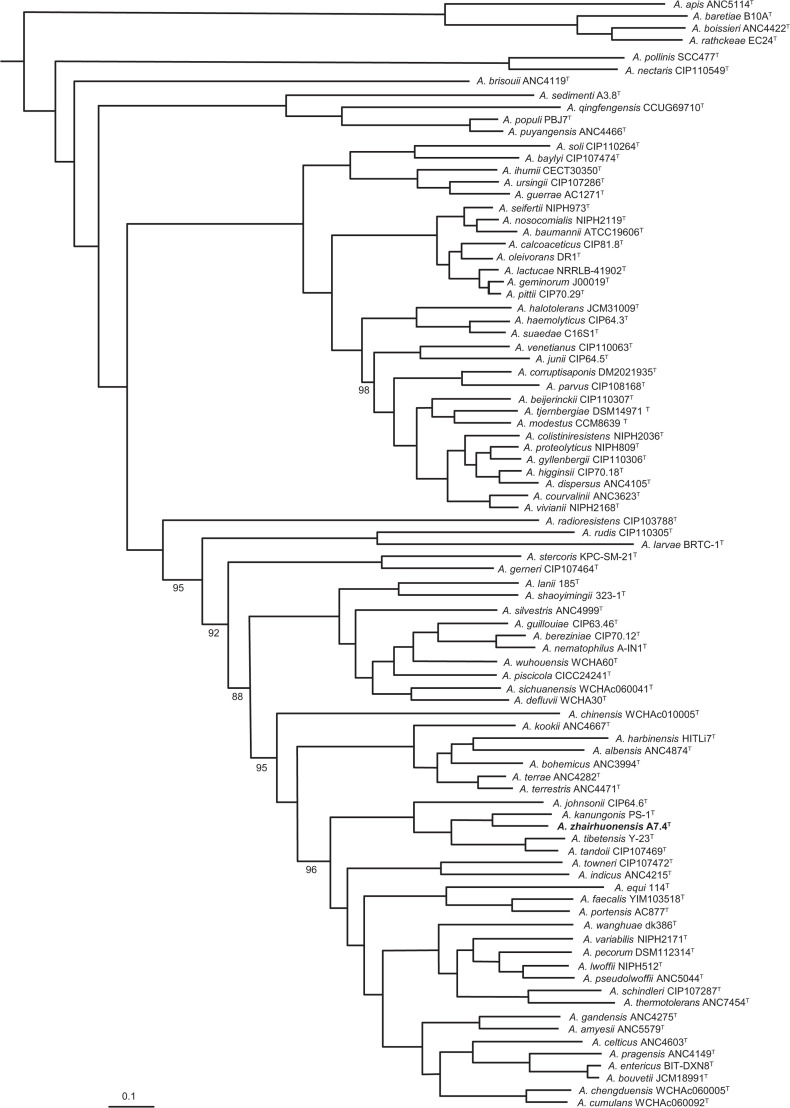 Fig. 2
Fig. 2| Characteristic* | 1† | 2 | 3 | 4 |
|---|---|---|---|---|
| Growth at 37 (℃) | − | + | + | + |
| Growth at 35 (℃) | + | + | + | + |
| Growth at 30 (℃) | + | + | + | + |
| Acidification of | − | − | − | − |
| Haemolysis of sheep blood | − | − | − | − |
| Liquefaction of gelatin | − | − | − | − |
| H2S production | + | − | + | − |
| API 20NE | ||||
| Esculin hydrolysis | + | + | − | − |
| Caprate assimilation | + | + | + | + |
| Malate assimilation | + | + | + | + |
| Citrate assimilation | + | − | + | + |
| Phenylactate assimilation | + | + | + | + |
| Assimilation (growth on)‡ | ||||
| | + | − | + | + |
| | − | − | − | + |
| 2,3-Butanediol | + | + | + | − |
| Glutarate |
| − | − | + |
| | − | − | + | + |
| Putrescine | + | − | + | + |
| | − | − | + | + |
| Tricarballylate | + | − | + | + |
| Tryptamine | + | + | − | + |
- —the Key Laboratory of Tropical Marine Ecosystem and Bioresource, MNR (CN)
- —National Key Research and Development Program of China
- —National Key Research and Development Program of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Identification and Quantification in Food · Microbial Community Ecology and Physiology
The genus Acinetobacter was initially proposed by Brisou and Prevot [1], with the earliest recorded species dating to 1911, when Beijerinck isolated a soil-derived bacterium subsequently reclassified from Micrococcus calcoaceticus [2]. The genus Acinetobacter is taxonomically classified within the family Moraxellaceae (order Pseudomonadales, class Gammaproteobacteria). Acinetobacter species exhibit remarkable ecological adaptability, colonizing diverse habitats including soil [34], aquatic systems [56], organisms [78] and food products [9]. According to the List of Prokaryotic names with Standing in Nomenclature (July 2025 release), the genus currently recognizes 118 species, with 87 maintaining a validly published name under the International Code of Nomenclature of Prokaryotes. These Gram-stain-negative, non-motile bacteria exhibit oxidase-negative and catalase-positive characteristics, with certain strains displaying haemolytic activity and gelatin hydrolysis capabilities. The genome typically comprises a single circular chromosome (2.6–4.7 Mb) and a strain-dependent set of plasmid replicons [10]. During a marine microbial diversity investigation in the East China Sea, strain A7.4^T^ was isolated from marine sediments near Zhairuo Island, Zhejiang province. This isolate formed milky, circular colonies on mineral salt medium (MM) agar. Phenotypic and biochemical characterization, together with genomic analysis and comparisons, confirmed its membership in the genus Acinetobacter; nonetheless, it was shown to be distinct from all other Acinetobacter species. Based on the polyphasic taxonomic study, we propose Acinetobacter zhairhuonensis sp. nov. (type strain A7.4^T^=MCCC 1K07162^T^=LMG 32567^T^).
Strain isolation
Strain A7.4^T^ was isolated from sediments of Zhairuo island (29° 56′ N, 121° 04′ E), Zhoushan city, Zhejiang province, China, in March 2019. One gram of beach sediments from the island was collected, suspended in artificial seawater, gradient diluted and spread on MM agar (1.0 g K_2_HPO_4_, 1.0 g KH_2_PO_4_, 2.0 g NaNO_3_, 0.5 g MgSO_4_.7H_2_O, 0.5 g (NH_4_)2_SO_4, 0.2 mg Na_2_MoO_4_ and 20.0 g agar per litre ddH_2_O, pH 7.0) [11]. A milky white colony with a diameter of 2 mm was picked and purified by the streak-plate technique on a fresh MM agar medium and designated as A7.4^T^. Strain A7.4^T^ has been deposited in the Marine Culture Collection of China (MCCC) and the Belgian Co-ordinated Collections of Micro-organisms (BCCM/LMG).
Genomic characteristics
The genomic DNA of A7.4^T^ was extracted using the MiniBEST Universal Genomic DNA Extraction Kit (Takara, Japan). The complete genome of strain A7.4^T^ was obtained using Illumina short-read sequencing and Nanopore long-read sequencing. Illumina paired-end sequencing (2×150 bp) was performed on a NovaSeq platform. Cutadapt was used to remove the adapters and bad quality reads [12]. Nanopore sequencing was conducted according to previously described methods [1314]. Primary genome assembly was conducted with NECAT using Nanopore reads [15], followed by error correction with Pilon (v.1.13) using the high-quality Illumina reads [16]. The assembled genome was annotated using National Center for Biotechnology Information (NCBI) Prokaryotic Genome Annotation Pipeline (version 6.9) with default parameters [17]. Using nanopore sequencing and Illumina technology, an intact genome of A7.4^T^ was assembled, comprising a single circular chromosome and one plasmid (Fig. 1). The genome size of strain A7.4^T^ is 3.57 Mb. The genomic DNA G+C content of strain A7.4^T^ is 41.3 mol%, which is within the range (34.9–49.6 mol%) reported for members of Acinetobacter [1018]. A total of 3,401 genes were predicted in the genome of strain A7.4^T^, including 3,245 protein-coding genes, 7 stretches of 5 S-16S-23S rRNA genes, 81 tRNAs, 4 ncRNAs and 50 pseudogenes.
A. zhairhuonensis A7.4T genome circular maps illustrated with the CGViewer pipeline, including one chromosome and one plasmid: CDS (blue), tRNA (red), rRNA (light green), G+C content (black), GC skew+ (dark green), GC skew- (violet).
Genome-based taxonomy and phylogeny
Current 16S rRNA gene sequencing analyses demonstrate limited resolution for distinguishing closely related genomic species within Acinetobacter due to the gene’s low polymorphism [19]. Genus-wide core-genome phylogenetic analysis represents the current gold standard for determining phylogenetic positions of novel bacterial genomes, surpassing single-gene marker approaches [682021]. Protein sequences of strain A7.4^T^ and 87 validly published Acinetobacter species were retrieved from NCBI (https://www.ncbi.nlm.nih.gov/), with OrthoFinder [22] identifying 717 single-copy orthologous genes (each showing >40% amino acid identity). Phylogenetic reconstruction was performed using FastTree [23] with visualization in Evolview v3 [24]. The core-genome phylogeny (Fig. 2) revealed A7.4^T^ forming a distinct subclade with Acinetobacter kanungonis PS-1^T^, which clustered with a sister branch containing Acinetobacter tibetensis Y-23^T^ and Acinetobacter tandoli CIP107469^T^. These three phylogenetically proximate strains were consequently selected as references for polyphasic characterization of A7.4^T^.
Phylogenetic tree reconstructed from 717 single-copy core genes shared between A. zhairhuonensis A7.4T and 87 validly published species of the genus Acinetobacter. Bootstrap support values (percentage of 1,000 replicates) are indicated at nodes only when<100%. The position of strain A7.4T is highlighted in bold.
The complete 16S rRNA gene sequences of strain A7.4^T^ and other validly published species were aligned by the clustalw software [25]. Phylogenetic trees were constructed using neighbour-joining and maximum evolution methods of the mega-x program [26] with 1,000 replicates to yield the bootstrap values. Phylogenetic analysis based on 16S rDNA (Figs S1 and S2, available in the online Supplementary Material) confirmed A7.4^T^ within the Acinetobacter genus, forming a separate evolutionary branch with A. kanungonis PS-1^T^, but failed to cluster with A. tibetensis Y-23^T^ and A. tandoli CIP107469^T^, diverging from the core-genome phylogeny results.
Genomic comparisons using average nucleotide identity (ANI), digital DNA–DNA hybridization (dDDH) and average amino acid identity (AAI) consistently supported the novel species status of strain A7.4^T^ (Table S1). Comparative analyses between strain A7.4^T^ and all 87 validly published Acinetobacter species demonstrated ANI values (calculated using JSpeciesWS [27]) ranged from 78.3% to 95.1%, dDDH values (Genome-to-Genome Distance Calculator (GGDC) v.3.0 formula 2 [28]) varied between 21.4% and 68.9% and AAI values (EzAAI [29]) spanned 84.5% to 94.98% – all below respective species delineation thresholds (ANI/AAI<96% [830]; dDDH <70% [2830]). Among these, the three phylogenetically closest strains (A. kanungonis, * A. tibetensis* and Acinetobacter tandoii) exhibited the highest genomic similarity values (ANI, 84.17–89.8%; dDDH, 28.3–40.0%; AAI, 90.78–94.98%), yet still showed divergence exceeding species delineation criteria. These results confirm strain A7.4^T^ as a novel species and are consistent with the core-genome phylogeny, further validating the superior accuracy of core-genome phylogenetic analysis compared to 16S rRNA gene-based phylogenetic approaches.
Screening of publicly available metagenomic datasets and NCBI databases
To further evaluate the occurrence and abundance of strain A7.4^T^ in natural environments, particularly marine habitats, blast analyses were performed targeting four genetic markers of strain A7.4^T^. Given that 16S rRNA V4 and V3–V4 hypervariable regions are widely used in microbiota studies due to their taxonomic accuracy [31], these regions were screened across 4,663 metagenomic samples (439 finished; 4,224 draft) from marine environments (ocean, 230; marine, 2,544; sea, 1,450) in the Integrated Microbial Genomes and Microbiomes [32]. High-coverage, high-identity (>97%) matches were further assessed using full-length 16S rRNA sequences and the rpoB gene region (RNA polymerase β-subunit; positions 2,916–3,773) [21]. The 16S rRNA V4 region yielded high-coverage, high-identity (>99%) matches, whereas the V3–V4 region, full-length 16S rRNA and rpoB gene exhibited lower coverage and identity, with rpoB showing no significant hits or identities below 92% (Excel File Dataset S1).
The four genetic markers were additionally screened against NCBI databases (Excel File Dataset S1). blast searches of the 16S rRNA V4 region revealed numerous 100% coverage/100% identity matches, including seawater metagenomes (KDVZ00000000.1) and uncultured clones. The V3–V4 region similarly showed multiple hits with 100% coverage and >99% identity from uncultured clones, suggesting widespread occurrence of A.7.4^T^-related sequences. For full-length 16S rRNA, high matches included Acinetobacter sp. ANC 4910 (SJNT01000006.1; 100% coverage, 99.0% identity) and an uncultured Acinetobacter sp. clone (HM159980.1; 100% coverage, 99.13% identity), in addition to A.7.4^T^ itself and the close relative A. tandoii (99.2% identity). However, genome-based comparisons (ANI, dDDH and AAI) between Acinetobacter sp. ANC 4910 and A.7.4^T^ indicated distinct species status, with all values below established thresholds (Table S1). No significant rpoB matches were observed (all identities<80%).
In summary, while the 16S rRNA V4 region confirms the environmental prevalence of strain A.7.4^T^, its identification based on a longer sequence remains limited. Obtaining additional isolates or high-quality metagenomes for this taxon requires further extensive investigation.
Comparative genomics and functional profiling
The complete genome sequences of strain A7.4^T^ and 87 validly published Acinetobacter species were subjected to comprehensive in silico functional analysis, revealing both conserved metabolic features and strain-specific functional characteristics.
Metabolic pathway annotation using KofamKOALA [33] and KEGG Decoder [21] identified 116 functional pathways classified into 20 categories (Fig. S3). Consistent with other Acinetobacter species [21], strain A7.4^T^ is capable of synthesizing most essential amino acids and possesses PGA synthesis proteins for poly-β-1,6-N-acetylglucosamine production, a polymer critical for biofilm formation [34]. Respiratory energy metabolism is supported by the presence of F-type ATPase and cytochrome bd complexes, correlating with the aerobic growth requirements of strain A7.4^T^. Genomic evidence of non-motility, indicated by absent flagellar genes, was confirmed through transmission electron microscopy (Fig. S4) and motility assays. Notably, while metabolic pathway profiles showed high conservation between strain A7.4^T^ and its phylogenetically proximate relatives (A. kanungonis, A. tibetensis and A. tandoii), significant genomic divergence was evident through ANI (<96%) and dDDH (<70%) values. This conservation of core metabolic functions despite genomic differentiation parallels observations in other Acinetobacter species descriptions [21].
Secondary metabolite gene clusters were investigated using the antiSMASH bacterial version website [35] with default parameters. The heatmap was created using an online platform (https://www.bioinformatics.com.cn) [36]. Comparative analysis of secondary metabolite biosynthetic gene cluster (BGC) profiles reveals distinct features in strain A7.4^T^ relative to other members of the genus (Fig. S5). While strain A7.4^T^ exhibits an intermediate overall BGC count (total=8), specific enrichments highlight its biosynthetic specialization. Notably, it encodes two non-ribosomal peptide synthetase (NRPS) clusters compared to phylogenetically close relatives such as A. tibetensis, A. tandoii and A. kanungonis, and more β-lactone BGCs than most Acinetobacter species. Conversely, strain A7.4^T^ lacks BGCs associated with hybrid (e.g. Polyketide Synthase-Nonribosomal Peptide Synthetase (PKS-NRPS)) and other undefined categories. Overall, strain A7.4^T^ combines moderate BGC diversity with specialized enrichments, distinguishing it from closely related species.
Phenotypic traits
Cell morphology and size of strain A7.4^T^ were observed under light microscopy (E100; Nikon) and transmission electron microscopy (JEM1230, Hitachi). Gram reaction was examined with a commercial Gram staining kit (BD) following the manufacturer’s protocol. The phenotypic and physicochemical features of A7.4^T^ and its reference strains were tested under the same conditions. Gliding motility was tested with freshly cultured cells according to the hanging drop method, and motility was investigated by semi-solid Luria-Bertani (LB) agar () [37]. Anaerobic growth was tested on LB agar after incubating in an anaerobic box (AnaeroPack^TM^-Anaero; Mitsubishi Gas Chemical Co., Inc.) at 30 ℃ for 7 days according to the manufacturer’s instructions. After 24 h of incubation on LB agar medium at 30 ℃, colonies of strain A7.4^T^ were milky, round and moist with a diameter of ~2 mm. Transmission electron microscopy showed a short rod cell of ~1.0–1.3 µm×1.0–1.6 µm (Fig. S4). Strain A7.4^T^ was Gram-stain-negative, aerobic and non-motile.
OD_600_ was used to monitor the growth of strain A7.4^T^ in LB medium at different temperatures (10, 30, 35, 37 and 40 ℃), different NaCl concentrations (w/v, 0–10% at increments of 1%) and different pH (pH 4–10, at intervals of 1 pH unit). The bacterial growth was evaluated using the following media: LB agar, nutrient agar (NA), trypticase soy agar (TSA) and Reasoner’s 2A agar (R2A) (all from Oxoid), with incubation at 30 °C for 3 days. Strain A7.4^T^ grew on all of the tested media, with the fastest growth on LB agar medium. Strain A7.4^T^ grew at 10–35 ℃ and pH 6.0–9.0 with 0–5% (w/v) NaCl. The optimal growth was observed at 30 ℃, pH 7.0, and in the presence of 2.5% (w/v) NaCl.
Oxidase and catalase activities were investigated by colour change in 1% (w/v) tetramethyl-p-phenylenediamine and generation of bubbles in 3.0% (v/v) H_2_O_2_, respectively. Methyl red and Voges–Proskauer reaction, H_2_S production, starch hydrolysis, gelatin hydrolysis, lipase (Tweens 20/40/60/80), phenylalanine deaminase and DNA hydrolysis were determined according to previously described methods [18]. Tests for the assimilation of the 36 carbon sources were performed according to previously described methods [38]. Other biochemical properties were tested using API 20NE and API 50CH tests (bioMérieux) according to the manufacturer’s instructions.
H_2_S production, catalase, Tweens 20, 40, 60, 80 and tyrosine hydrolysis tests of strain A7.4^T^ are positive. Hydrolysis of starch, casein, acidification of d-glucose, haemolysis of sheep blood, liquefaction of gelatin, methyl red, Voges–Proskauer, phenylalanine deaminase and DNA hydrolysis tests are negative. In the API 20NE kit, a positive reaction for aesculin hydrolysis and assimilation of malate, citrate and phenylacetate is observed. The key and differential characteristics between strain A7.4^T^ and three reference strains are given in Table 1. The negative reaction of strain A7.4^T^ on API kits is shown in Table S2. The shared carbon source assimilation patterns between strain A7.4^T^ and reference strains are listed in Table S3.
Chemotaxonomic traits
The fatty acid [39] composition of the cells grown on the TSA medium for 2 days was determined according to the protocol of the Sherlock Microbial Identification System (MIDI) [40]. The predominant fatty acids of strain A7.4^T^ (> 5.0% of the total amounts) were summed feature 3 (C_16:1_ ω7c/C_16:1_ ω6c) (31.2%), C_18:1_ ω9c (22.2%), C_16:0_ (16.7%), C_12:0_ (7.5%) and C_12:0_ 3-OH (5.1%). The fatty acid composition of A7.4^T^ is consistent with previous results for recognized species of the genus Acinetobacter, justifying its placement in this genus [48]. The detailed fatty acid profile of strain A7.4^T^ is summarized in Table S4.
Polar lipids [41] were separated and analysed with TLC Silica gel 60 F_254_ (Merck 1.05554.0001) and a freshly prepared developing agent according to the method described by Minnikin et al. [42]. The polar lipids of strain A7.4^T^ contained diphosphatidylglycerol, phosphatidylglycerol, phosphatidylethanolamine, two phospholipids (PL1–2) and two aminolipids (Fig. S6). To determine respiratory quinones, strain A7.4^T^ was incubated in 500 ml LB medium for 2 days at 30 °C, harvested and lyophilized. Then respiratory quinones of strain A7.4^T^ were extracted and analysed by HPLC according to the process described previously [43]. The main quinones of strain A7.4^T^ are Q-8 and Q-9 (Fig. S7). The chemotaxonomic characteristics of strain A7.4^T^ compared with the closely related type strains A. tibetensis Y-23^T^ [11], A. tandoii CIP107469^T^ [44] and A. kanungonis PS-1^T^ [8] further support that strain A7.4^T^ represents a member of the genus Acinetobacter.
Description of Acinetobacter zhairhuonensis sp. nov.
Acinetobacter zhairhuonensis (zhai.rhu.o.nen’sis. N.L. masc. adj. zhairhuonensis, pertaining to Zhairuo, an island of the East China Sea, from where the type strain was isolated).
Cells are Gram-stain-negative, aerobic, non-motile, oxidase-negative, catalase-positive, short rods, 1.0–1.3 µm wide and 1.0–1.6 µm long. It can grow in LB, NA, TSA and R2A medium, with LB yielding the fastest and best growth. After culturing on LB agar plates for 3 days, the colonies are round, smooth, milky, opaque and moist with a diameter of about 2 mm. Growth occurs at 10–35 ℃ (optimum 30 ℃), pH 6.0–9.0 (optimum pH 7.0) and 0–5% (w/v) NaCl (optimum 2.5%). Positive for H_2_S production, catalase, Tweens 20, 40, 60, 80 and tyrosine hydrolysis. Negative for hydrolysis of starch, casein, acidification of d-glucose, haemolysis of sheep blood, liquefaction of gelatine, methyl red, Voges–Proskauer, phenylalanine deaminase and DNA hydrolysis. Of 36 tested carbon sources, assimilation was positive for 22 substrates: acetate, trans-aconitate, 4-aminobutyrate, l-arginine, l-aspartate, benzoate, 2,3-butanediol, citrate, ethanol, l-glutamate, glutarate, l-histidine, 4-hydroxybenzoate, dl-lactate, d-malate, malonate, l-ornithine, phenylacetate, l-phenylalanine, putrescine, tricarballylate, tryptamine, whereas negative results were obtained for adipate, β-alanine, l-arabinose, azelate, citraconate, gentisate, d-gluconate, d-glucose, histamine, l-leucine, levulinate, d-ribose, l-tartrate and trigonelline. In the API 20NE kit, a positive reaction for aesculin hydrolysis and assimilation of caprate, malate, citrate and phenylacetate. All the other tests are negative in the API 50CH kit.
The predominant fatty acids include Summed feature 3 (C_16:1_ ω7c/C_16:1_ ω6c), C_18:1_ ω9c, C_16:0_, C_12:0_ and C_12:0_ 3-OH. Polar lipids include diphosphatidylglycerol, phosphatidylglycerol, phosphatidylethanolamine, two phospholipids (PL1–2) and two aminolipids. The main respiratory quinones are Q-8 and Q-9. The genome comprises a single circular chromosome and one plasmid. The genome size and genomic DNA G+C content are 3.57 Mb and 41.3 mol%, respectively.
The type strain, A7.4^T^ (=MCCC 1K07162^T^=LMG 32567^T^), was isolated from sediments of Zhairuo Island in Zhejiang, China. The GenBank accession numbers for the genome and 16S rRNA gene sequence of A. zhairhuonensis A7.4^T^ are GCA_047300875 and MW287273, respectively.
Supplementary material
10.1099/ijsem.0.006936Uncited Supplementary Material 1.
10.1099/ijsem.0.006936Uncited Supplementary Material 2.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Brisou J Prevot AR Studies on bacterial taxonomy. X. The revision of species under Acromobacter group Ann Inst Pasteur 19548672272813197842 · pubmed ↗
- 2Beijerinck M Pigments as products of oxidation by bacterial action Koninklijke Nederlandse Akademie van Wetenschappen Proceedings Series B Physical Sciencesvol 13191010661077
- 3Choi JY Ko G Jheong W Huys G Seifert H et al Acinetobacter kookii sp. nov., isolated from soil Int J Syst Evol Microbiol 2013634402440610.1099/ijs.0.047969-023950148 · doi ↗ · pubmed ↗
- 4Pan H Li J Liu H-H Lu X-Y Zhang Y-F et al Acinetobacter tibetensis sp. nov., isolated from a soil under a greenhouse in Tibet Curr Microbiol 2023805110.1007/s 00284-022-03158-z 36547704 · doi ↗ · pubmed ↗
- 5Li W Zhang D Huang X Qin W Acinetobacter harbinensis sp. nov., isolated from river water Int J Syst Evol Micr 2014641507151310.1099/ijs.0.055251-024478215 · doi ↗ · pubmed ↗
- 6Nemec A Radolfová-KřížováL MaixnerováM Nemec M ŠpanělováP et al Delineation of a novel environmental phylogroup of the genus Acinetobacter encompassing Acinetobacter terrae sp. nov., Acinetobacter terrestris sp. nov. and three other tentative species Syst Appl Microbiol 20214412621710.1016/j.syapm.2021.12621734107439 · doi ↗ · pubmed ↗
- 7Wolf S Barth-Jakschic E Birkle K Bader B Marschal M et al Acinetobacter geminorum sp. nov., isolated from human throat swabs Int J Syst Evol Microbiol 20217100501810.1099/ijsem.0.00501834633923 PMC 8604166 · doi ↗ · pubmed ↗
- 8Das L Deb S Das SK Description of Acinetobacter kanungonis sp. nov., based on phylogenomic analysis Int J Syst Evol Microbiol 20217100483310.1099/ijsem.0.00483334128781 · doi ↗ · pubmed ↗