Pathogenicity and transmission of Morganella morganii in honey bees
Yijun Chen, Qiang Huang, Jeffrey Dvorin, Jeffrey Dvorin, Jeffrey Dvorin

TL;DR
A deadly bacterium, Morganella morganii, is transmitted by Varroa mites to honey bees, causing high mortality and contributing to colony collapse.
Contribution
The study isolates and characterizes Morganella morganii as a novel pathogen transmitted by Varroa mites to honey bees.
Findings
Morganella morganii causes over 30% mortality in honey bees at low doses.
The bacterium is transmitted from mites to bees and vice versa, but not through social bee interactions.
Transmission rates are 92.1% from bees to mites and 68.49% from mites to bees.
Abstract
Honey bees provide essential pollination services in the ecosystem. The high annual loss of honey bees has raised concerns about global food security and the agricultural economy. As a primary stressor causing colony failure, the mite Varroa destructor feeds on the hemolymph and the bee’s fat body tissue. The Varroa mite-associated deformed wing virus has been extensively studied because it can be found in each individual mite and causes bee mortality. A recent study shows that the Varroa mite can transmit pathogenic bacteria, while the transmission route remains unclear. In this study, we isolated and assembled a previously uncultured bacterium, Morganella morganii, from the mites Varroa destructor. This pathogenic bacterium exhibited a high case fatality rate, as evidenced by 215 cells causing over 30% mortality in pupae and adult bees. Using a fluorescent protein-tagged strain, we…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
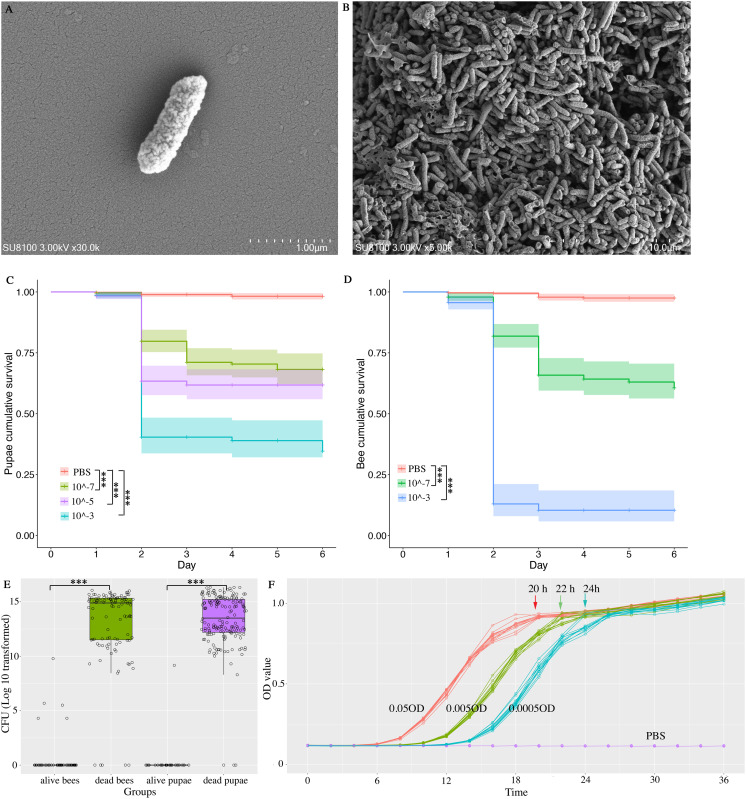 Fig 1
Fig 1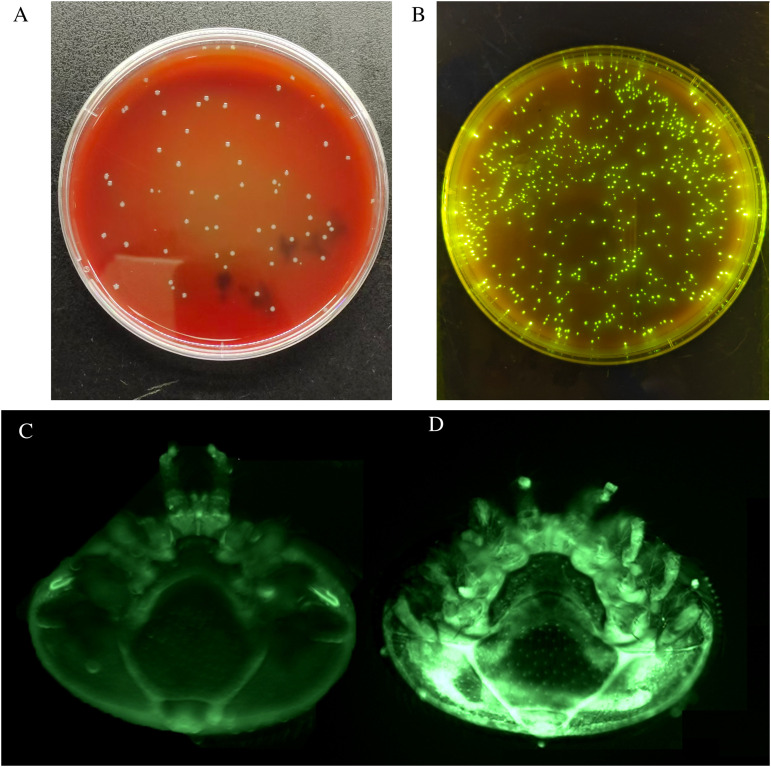 Fig 2
Fig 2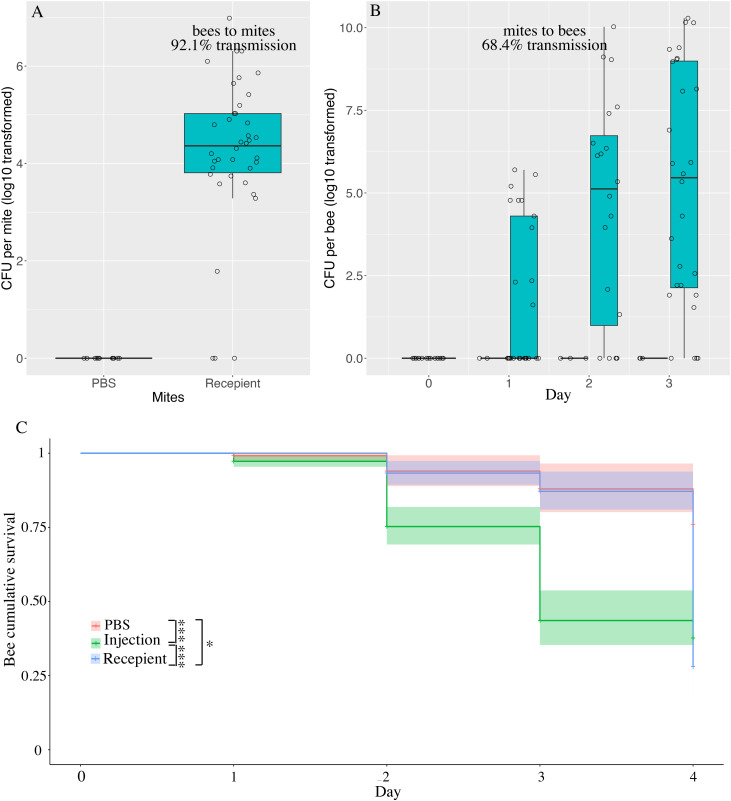 Fig 3
Fig 3- —Jiangxi Double Thousand Plan
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect and Pesticide Research · Plant and animal studies · Insect and Arachnid Ecology and Behavior
Introduction
The honey bee, Apis mellifera, is a crucial domesticated pollinator that substantially improves fruit set for flowering plants. Recently, the annual loss of honey bee colonies has approached 60% in North America [1], and the mite Varroa destructor is the primary stressor causing colony failure [2–4]. V. destructor is an ectoparasitic mite that feeds on the hemolymph and bee’s fat body tissue [5]. The Varroa mites parasitize larvae, pupae, and adult bees [6]. Once parasitized by the Varroa mites, honey bees exhibit immature aging [7], impaired flying and cognition [8,9], and shortened lifespan [10,11]. Upon feeding, the mites can inject a variety of viruses into the hemolymph of bees: among them, the deformed wing virus (DWV) is one of the most prevalent, causing developmental disorders in infected individuals [12,13]. The naïve Varroa mites can also acquire the viruses when feeding on infected bees [14].
DWV has been extensively investigated because of its ubiquity in V. destructor [12]. DWV can be horizontally transmitted from bee to bee, as validated by fluorescent tagging, and infected individuals serve as a reservoir for naive mites to acquire and subsequently transmit the virus [14]. The Varroa mites also harbor several bacteria: a previous 16S sequencing study identified a few bacteria in mites and honey bees, including Morganella, Spiroplasma, and Enterococcus, suggesting these bacteria may transmit between them [15,16]. Given the difficulties in cultivating these pathogens and the complexity of the bacterial community found in Varroa mites, this field of study remains largely unexplored. Until recently, Enterococcus faecalis was isolated from dead bees, which is vectored by Varroa mites. E. faecalis caused substantial mortality in the honey bee A. mellifera [17]. Thus, the mite also serves as a vector for pathogenic bacteria of honey bees. However, the transmission routes of E. faecalis are not fully understood.
In a previous study, a few bacterial pathogens were inferred in both mites and honey bees [16]. We hypothesize that the Varroa mite transfers and acquires bacterial pathogens upon feeding the bees. To investigate the virulence and transmission routes of the bacteria vectored by V. destructor, we first isolated and cultured those bacteria. Then, we injected the bacterial inoculum into bees to observe the bacterial proliferation and bee mortality. Specifically, we isolated the first strain of Morganella morganii in Varroa mites, which is highly virulent in honey bees. Unlike the DWV, M. morganii can’t be transmitted from bees to bees. Instead, transmission of M. morganii depends on Varroa mites. Honey bees and Varroa mites both serve as a reservoir of M. morganii, facilitating the mites in acquiring and transmitting it. Our data revealed a high lethality induced by M. morganii through mite bite: this might partially explain bee colony losses during late autumn and winter, when intraspecific contact among bees is high.
Materials and methods
Morganella morganii isolation, morphology, and growth
We collected V. destructor mites from adult honey bees A. mellifera, using the sugar-shaking method from multiple colonies in the experimental apiary in Jiangxi Agricultural University [18]. We collected and rinsed 50 mites under distilled water, pooled each of 5 mites, and homogenized them in 500 μL Phosphate Buffered Saline (PBS). We further diluted the homogenates to 10^-3^ and plated 50 μL on Lysogeny Broth (LB), Columbia Blood Agar (CBA), Trypticase soy agar (TSA), Potato Dextrose Agar (PDA), Brain Heart Infusion Agar (BHI). We prepared two plates for each culturing medium and incubated one plate in a CO_2_ incubator and the other in a regular incubator. Single colonies were collected and used for 16S sequencing to infer the bacterial species. The morphology of single cells was photographed using scanning electron microscopy. To investigate the rate of M. morganii proliferation, we performed a time-series recording of OD_600_ in vitro. We recorded the Optical Density (OD) at 2-hour intervals until 36 hours post-plating to observe the growth rate in LB liquid medium (N = 12 replicate samples for each concentration).
M. morganii genome assembly and gene annotation
The DNA of a single colony of M. morganii was extracted using Sodium Dodecyl Sulfate (SDS) and sheared to 30 Kbp to be sequenced in PacBio Sequel II according to the Illumina DNA preparation instructions. The highly accurate long reads (HIFI) were assembled using Flye assembler (Version 0.3-3) with default parameters [19], and the gene features were annotated using the NCBI Prokaryotic Genome Annotation Pipeline [20]. To assess the completeness of the genome, we aligned the protein sequence to the conserved single-copy orthologs of the bacterial kingdom (BUSCO Bacteria_odb10 database) [21]. The genomes’ average nucleotide identity (ANI) was pairwise calculated using FastANI (version 1.34) with default parameters [22].
Colonization of M. morganii and mortality in honey bees
To simulate the Varroa mites’ feeding behavior, we injected the inoculum into the abdomen of pupae and gnotobiotic honey bees A. mellifera. Specifically, we prepared M. morganii solution at OD_600_ = 1 and diluted with the PBS solution to 10^-3^, 10^-5^, and 10^-7^ of inoculum. To estimate the number of CFUs in each dilution, we first counted the number of cells in 1µL of 10^-7^ inoculum (N = 11 replicates); then, we derived the titer of the 10^-5^ and 10^-3^ dilutions, as a proportion of the average number of CFU counts in the 10^-7^ dilution. We injected 1μL inoculum of 10^-3^ (N = 127 pupae), 10^-5^ (N = 126 pupae), and 10^-7^ (N = 128 pupae) into the pupae. Additionally, we injected 1 μL PBS into 174 pupae as the control group. Dead pupae were recorded and homogenized to manually count CFU (Colony Forming Unit) using a dilution series. The remaining pupae were collected to count CFU at 7 dpi (Days Post Inoculation). Additionally, we injected 1μL inoculum of 10^-3^ (N = 102 gnotobiotic bees), and 10^-7^ (N = 116 gnotobiotic bees) into gnotobiotic adult honey bees. Again, we injected 1 μL PBS into 102 gnotobiotic bees as the control group. Dead bees were recorded and homogenized to count CFU manually. All live bees were collected to count CFU at 7 dpi.
Oral transmission of M. morganii
We engineered M. morganii to differentiate it from the wild type that the mites might carry. The engineered M. morganii:pBTK520 expresses green fluorescent protein with Spectinomycin resistance. First, we investigated whether fluorescent M. morganii can be transmitted among bees through co-housing them. To this aim, we injected 1 μL of 10^-7^ inoculum into 15 gnotobiotic bees, marking them with a color patch placed on the thorax for identification. We co-housed these infected bees with 15 naive bees in a cup at 1:1 ratio with 5 replicates (N = 75 gnotobiotic infected bees and 75 naïve bees). As a control group, we injected 1 μL PBS into 15 gnotobiotic bees as the PBS group with 5 replicates (N = 75 bees). Dead bees were recorded daily and homogenized for CFU counting. At day 7, the remaining bees were homogenized for CFU counting. M. morganii is engineered using the Bee Microbiome Toolkit [23]. Detailed procedures are provided in S1 Appendix.
Mite vectored transmission of M. morganii
We investigated the naïve Varroa mites’ acquisition of M. morganii from infected bees, as well as the transmission of M. morganii from infected mites to naïve bees. We selected the concentration of 10^-7^ to investigate the transmission routes of M. morganii among bees and mites because this concentration allows bees to survive the experiment before being killed by the infection. First, we injected 25 gnotobiotic bees with the fluorescent M. morganii, transferring one mite to each bee’s body surface for 24 hours, providing 5 replicates of this experiment (N = 125 infected bees and 125 mites): we refer to this as the injection group. At 24 hours, we homogenized the mite for CFU counting to infer the proportion of mites that acquired the fluorescent M. morganii (N = 38 mites) from the injected bees. After 24 hours, we randomly selected 15 of those mites, transferring each one to a naïve bees’ body surface for 3 days (5 replicates, N = 75 naive bees and 75 infected mites) as the recipient group. Furthermore, we injected PBS into 15 gnotobiotic bees and transferred mites to the bees’ body surface at a 1:1 ratio (5 replicates, N = 75 gnotobiotic bees and 75 mites) as the control group. Dead bees were recorded daily and homogenized for CFU counting. At day 7, the remaining bees were homogenized for CFU counting.
Statistics
The statistics were performed in R (Version 4.2.2) [24]. Colonization and persistence of M. morganii were analyzed using the Kruskal-Wallis test. Survival was analyzed using the Kaplan-Meier estimate in the survival package [25], and multiple comparisons were adjusted for False Discovery Rate (FDR). M. morganii titers within bees and mites were analyzed using the Wilcoxon rank-sum test, and multiple comparisons were adjusted for FDR. The figures were plotted using the ggplot2 package in R [26].
Results
Strain isolation, colonization, and genome assembly
We isolated 33 bacterial species (Table A in S1 Appendix) and successfully cultivated M. morganii in LB, CBA, and BHI in both regular and CO_2_ incubators. We sequenced a single colony of M. morganii, assembled and annotated its genome under the NCBI genome GCA_044772875.1 (Fig 1A and 1B).
Morganella morganii in mites.We isolated M. morganii from the mite Varroa destructor. It appears rod-shaped under a scanning electron microscope (A), with no clear indication of a biofilm (B). Injection of M. morganii caused high mortality in pupae (C) and adult bees (D). High bacterial titer was identified in dead pupae and dead bees, which is rarely found in those that survived the M. morganii injection (E). The proliferation of M. morganii reached a stationary phase in approximately 24 hours in a series of diluted homogenates (F).
Its genome is 3.8 Mbp in a single contig, with 3,531 protein-coding genes (Table B in S1 Appendix), including 122 complete and 2 fragmented BUSCOs. We compared the isolated M. morganii genome with 27 previously published M. morganii genomes from various hosts. M. morganii exhibits minor genome variance between vertebrate and invertebrate hosts (Fig A and Table C in S1 Appendix). The Average Nucleotide Identity (ANI) is 92.0% with a 95% confidence interval from 91.4% to 92.6% (Table C in S1 Appendix).
M. morganii induced mortality in pupae and adult bees
To investigate the impact of M. morganii on honey bee mortality, we injected the bacterial inoculum into the pupae and adult bees, simulating the feeding behavior of Varroa mites. The pupae injected with PBS showed a high survival (98.2%, 95% CI: 96.9% ~ 99.5%), while the injection of M. morganii resulted in a significantly high mortality at all concentrations. Specifically, the pupae survival was the lowest in the 10^-3^ group (34.6%, 95% CI: 25.6% ~ 46.8%, Log-rank test, adjusted P < 0.01) compared with the 10^-5^ (61.8%, 95% CI of 55.9% ~ 62.8%; Log-rank test, adjusted P < 0.01) and the 10^-7^ groups (68.2%, 95% CI of 66.2% ~ 74.8.0%; Log-rank test, adjusted P < 0.05, Fig 1C). The injection of M. morganii also resulted in low survival in adult bees. The bee survival was again the lowest in the 10^-3^ group (10.4%, 95% CI of 18.5% ~ 58.3%) compared with the 10^-7^ (60.6%, 95% CI of 52.9% ~ 69.5%; Log-rank test, adjusted P < 0.001) and the PBS groups (95% CI of 96.0% ~ 99.1%; Log-rank test, adjusted P < 0.05, Fig 1D).
To investigate whether the bee mortality was caused by the proliferation of the injected M. morganii, we collected dead and live individuals. We plated their homogenate of to count the CFU. We found that the CFU of M. morganii is 10^13.5^ (95% CI of 10^13.2^ ~ 10^15.4^) in dead pupae, while this pathogenic bacterium is rarely found in live pupae (Wilcoxon rank test P < 0.0001). In adult bees, M. morganii again showed a higher CFU in dead bees of 10^13.7^ (95% CI of 10^13.3^ ~ 10^14.1^) than in live bees of 10^5.9^ (95% CI of 10^3.9^ ~ 10^7.8^, Wilcoxon rank test P < 0.0001). Comparatively, the dead adult bees and dead pupae showed a similar number of CFU (Wilcoxon rank test P = 0.35, Fig 1E). M. morganii exhibited a rapid proliferation and reached the stationary phase in approximately 24 hours (Fig 1F), which may explain the observed high bacterial titer in dead bees at 2 dpi.
The transmission of M. morganii
By plating the inoculum of 10^-7^ concentration, 1 μL of the inoculum is approximately 215 CFU (95% CI: 168 ~ 261). First, we investigated whether M. morganii (Fig 2A and 2B) can be transmitted from infected bees to naïve bees through co-housing. We injected M. morganii into gnotobiotic bees. Then, we co-housed the inoculated bees with naïve ones for 7 days. At the end of the experiment, M. morganii titer was high in the injection group (10^12.6^ cells, 95%CI: 10^11.9^ ~ 10^13.2^), while it was absent in the co-housed naïve bees and PBS group (Fig B in S1 Appendix). All co-housed naïve bees survived, except for one dead bee in which M. morganii was not detected. Thus, we conclude that M. morganii can’t be transmitted from infected bees to naïve bees through co-housing.
Varroa vectored transmission of M. morganii.We engineered M. morganii (A) to express green fluorescent protein using the Bee Microbiome Toolkit as M. morganii:pBTK520 (B). We inject gnotobiotic bees with M. morganii:pBTK520 or PBS, and then transfer a mite to the body surface of the inoculated bees. The green fluorescence is not detected in mites transferred to bees injected with PBS (C). Comparatively, the mite transferred to bees injected with M. morganii:pBTK520, exhibits clear green fluorescence (D).
Secondly, we investigated the transmission from infected bees to naïve Varroa mites. After introducing the mites to the PBS-injected bees, no fluorescence is observed in the mites (Fig 2C). Comparatively, after introducing the mites to the fluorescent M. morganii infected bees, we observed a clear fluorescence in the mites (Fig 2D). By plating the homogenates of the mites, we found the titer of M. morganii approached 10^4.5^ per mite (95% CI of 10^4.2^ ~ 10^4.9^, Fig 3A), after parasitizing M. morganii injected bees, while 3 dead mites were uninfected. In total, 35 out of 38 mites harbored M. morganii once they parasitized bees that had been injected with M. morganii. Thus, 92.1% of mites acquired M. morganii from infected bees through parasitism.
Acquisition of M. morganii from bees and mites.We found 35 out of 38 mites that were transferred to bees injected with M. morganii, harbored M. morganii (A). Thus, the cumulative incidence of Varroa mites’ acquiring M. morganii from bees is 92.1% in 24 hours through parasitism. The mites that carry M. morganii vector the pathogenic bacteria to other naïve bees. The cumulative incidence of bees’ acquiring M. morganii from mites is 68.4% (B). This pathogenic bacterium resulted in a 72% mortality rate among recipient bees, which is comparable to the 62.4% mortality rate in the injection group (C).
Thirdly, we investigated the transmission of M. morganii from infected Varroa mites to naïve bees. We collected the mites that had parasitized the injected bees and transferred those mites to the recipient gnotobiotic bees. We found that M. morganii can colonize and persist in most bees in the recipient group, as reflected by the CFU (10^5.6^, 95% CI of 10^4.8 ^~ 10^6.3^, Fig 3B). Since all alive mites acquired M. morganii from the infected bees, the cumulative incidence of naïve bees acquiring M. morganii from the infected mites is 68.49%.
To infer whether the mortality of injected and parasitically transmitted M. morganii is similar, we compared the survival of bees infected through injection, to that of recipient bees. At 4 dpi, we found that the bees in the injection group showed a low survival (37.6%, 95% CI of 28.8% ~ 49.2%), which was even lower in the recipient group (28%, 95% CI of 16.3% ~ 48.2%). The injection group showed a high mortality as early as 2 dpi (24.7%, 95% CI of 18.1% ~ 30.8%). Comparatively, the bees in the recipient group experienced a delay of mortality to 4 dpi, reflecting the accumulation of the bacteria. The injection may cause physical damage to the bees, reflected by the mortality in the PBS group. The bees in the PBS group showed a 76.0% survival (95% CI: 62.8% ~ 91.9%), which is higher than the injection (Log rank test, adjusted P < 0.001) and recipient groups (Log rank test, adjusted P < 0.001, Fig 3C).
Discussion
In this study, we isolated the first M. morganii in Varroa mites and highlighted that this pathogenic bacterium is highly virulent in honey bees. M. morganii is an opportunistic pathogen that infects a broad range of invertebrates [27,28], and vertebrates [29,30], including humans [31]. However, the virulence of M. morganii is rarely investigated. A previous study found that 10^5^ CFU of M. morganii resulted in 100% mortality in flies, and a lower dosage of 10^2^ CFU caused 44% mortality [32]. M. morganii also reduced the flight ability and pupal weight of the flies [33]. In nematodes, 10^6^ CFU of M. morganii leads to 100% mortality of infected nematodes in 24 hours, and 10^2^ CFU leads to 20% mortality [27]. In our data, M. morganii caused over 30% mortality in honey bees at a low dosage of 10^2^ CFU. However, the titer of M. morganii increases to 10^10^ CFU within 24 hours, thereby enhancing the case fatality rate, leading to the observed acute mortality.
The transmission route could determine the outcomes of the pathogen infection. For example, another opportunistic pathogen of honey bees, Serratia marcescens, caused approximately 20% mortality when infected through feeding, while injecting it leads to 100% mortality [34]. In our study, M. morganii is transmitted through the feeding of Varroa mites. At the individual level, the salivary secretions of Varroa mites can prevent the healing of the wound, through which the mites suck the hemolymph and fat body [35]. During feeding, the Varroa mites inject several pathogens into the bees, as is found in other blood-feeding arthropods [36,37]. The hosts then serve as a reservoir for the vectors to pick up and transmit the pathogens [38]. For example, Deformed wing virus (DWV) is ubiquitously harbored and vectored by the Varroa mite [39]. The titer of DWV is relatively low in honey bees when the Varroa mites are absent, which increases in orders of magnitude when the vector arrives [12,40]. In our study, we found that M. morganii couldn’t be transmitted from infected bees to naïve bees during the cohousing, while it exhibited a 92.1% transmission from infected bees to mites; this is much higher than the reported 29.3% DWV transmission from bees to mites [14]. Once the Varroa mites acquire the pathogenic bacteria, they disperse the bacteria to other bees. Thus, the population of mites determines the prevalence and titer of M. morganii.
In winter, the honey bee queen stops laying eggs, and the bees cluster to keep warm [41]. The Varroa mite can easily switch from one bee to another. Thus, viruses and bacteria are dispersed among nestmates in a short period of time. Hence, the mite population determines whether a honey bee colony can survive the winter [11]. In this study, we isolated a pathogenic bacterium M. morganii, which is vectored by mites and causes substantial mortality in honey bees. This study is limited to the germfree bees, and it is interesting to investigate the impacts of gut microbes on M. morganii for future studies.
Supporting information
S1 AppendixFig A. Heatmap of Average Nucleotide Identity of Morganella strains. The one isolated from the Varroa mite shows an average identity of 92%. The genome assembled in this study is highlighted. Fig B. The transmission of M. morganii through social contacts. We injected the engineered M. morganii:pBTK520 into bees and co-housed these bees with naïve ones. The engineered M. morganii:pBTK520 developed high titers in the injection group, which is absent in bees in the co-housing group and PBS group. The data suggest that the symbiont can’t be transmitted through social contacts. Table A. The bacterial species isolated from mites. All mites were rinsed to remove microbes from the body surface. The mites were then homogenized to isolate single colonies under various conditions. In total, 33 bacterial species were isolated. Morganella is particularly interesting because it has been reported in both bees and mites in a previous metagenome study, suggesting a mutual transmission. Table B. The genome assembly statistics of M. morganii CYJ1 genome. The genome is assembled into a single contig, with 3.8 Mbp. Out of 124 conserved single-copy genes in the bacterial kingdom, 122 were identified, and the remaining 2 were fragmented. Table C. Genomes of Morganella morganii for ANI (Average Nucleotide Identity) analysis. We randomly selected 20 complete genomes to perform ANI and AAI comparisons with the strain CYJ1.(DOCX)
S1 DataData deposit.(XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lamas ZS, Rinkevich F, Garavito A, Shaulis A, Boncristiani D, Hill E, et al. Viruses and vectors tied to honey bee colony losses. bio Rxiv. 2025; 2025.05.28.656706. 10.1101/2025.05.28.656706 · doi ↗
- 2Requier F, Leyton MS, Morales CL, Garibaldi LA, Giacobino A, Porrini MP, et al. First large-scale study reveals important losses of managed honey bee and stingless bee colonies in Latin America. Sci Rep. 2024;14(1):10079. doi: 10.1038/s 41598-024-59513-6 38698037 PMC 11066017 · doi ↗ · pubmed ↗
- 3Johannesen J, Wöhl S, Berg S, Otten C. Annual fluctuations in winter colony losses of Apis mellifera L. are predicted by honey flow dynamics of the preceding year. Insects. 2022;13(9):829. doi: 10.3390/insects 13090829 36135530 PMC 9501173 · doi ↗ · pubmed ↗
- 4Warner S, Pokhrel LR, Akula SM, Ubah CS, Richards SL, Jensen H, et al. A scoping review on the effects of Varroa mite (Varroa destructor) on global honey bee decline. Science of The Total Environment. 2024;906:167492. doi: 10.1016/j.scitotenv.2023.16749237778563 · doi ↗ · pubmed ↗
- 5Ramsey SD, Ochoa R, Bauchan G, Gulbronson C, Mowery JD, Cohen A, et al. Varroa destructor feeds primarily on honey bee fat body tissue and not hemolymph. Proc Natl Acad Sci U S A. 2019;116(5):1792–801. doi: 10.1073/pnas.1818371116 30647116 PMC 6358713 · doi ↗ · pubmed ↗
- 6Rosenkranz P, Aumeier P, Ziegelmann B. Biology and control of Varroa destructor. J Invertebr Pathol. 2010;103 Suppl 1:S 96-119. doi: 10.1016/j.jip.2009.07.016 19909970 · doi ↗ · pubmed ↗
- 7Ellis JT, Rangel J. Stress drives premature hive exiting behavior that leads to death in young honey bee (Apis mellifera) workers. Biol Res. 2024;57: 92. doi: 10.1186/s 40659-024-00569-z 39593109 PMC 11600856 · doi ↗ · pubmed ↗
- 8Muijres FT, van Dooremalen C, Lankheet M, Lugt H, de Vries LJ, Van Langevelde F. Varroa destructor infestation impairs the improvement of landing performance in foraging honeybees. R Soc Open Sci. 2020;7(9):201222. doi: 10.1098/rsos.201222 33047066 PMC 7540786 · doi ↗ · pubmed ↗