A novel homoarginine-containing cyclic peptide pioamide with selective antipseudomonal activity isolated from the nematode symbiont Photorhabdus khanii
Yu Imai, Sangkeun Son, Miho Sasaki, Libang Liang, Michael F. Gates, Meghan Ghiglieri, Takeshi Shimosato, Chandrashekhar Honrao, Xiaoyu Ma, Jason J. Guo, Kim Lewis

TL;DR
A new antibiotic called pioamide, isolated from a nematode symbiont, selectively targets Pseudomonas aeruginosa and could help combat drug-resistant Gram-negative bacteria.
Contribution
Discovery of pioamide, a novel cyclic peptide antibiotic with selective antipseudomonal activity from Photorhabdus khanii.
Findings
Pioamide selectively kills Pseudomonas aeruginosa without affecting other bacteria or human cells.
Resistance to pioamide in P. aeruginosa is conferred by mutations in the pmrB gene.
Pioamide's activity is not affected by porins or efflux pumps, suggesting a unique mechanism of action.
Abstract
Gram-negative bacteria harbor an outer membrane that physically protects them from the penetration of antibiotics into the cells. This barrier makes it challenging to develop antibiotics that effectively kill Gram-negative pathogens. The entomopathogenic bacterium Photorhabdus species produces various bioactive molecules and is receiving attention as an attractive source of novel antibiotics. We identified a novel antipseudomonal antibiotic, pioamide, from the culture supernatant of Photorhabdus khanii HGB1456, a strain that produces darobactin, which selectively kills Gram-negative bacteria. Pioamide, a pentapeptide antibiotic with a molecular weight of 704, exhibits selective activity against Pseudomonas aeruginosa but does not exhibit any activity against other bacteria or human cell lines. Whole-genome sequencing of spontaneous pioamide-resistant mutants of P. aeruginosa revealed…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
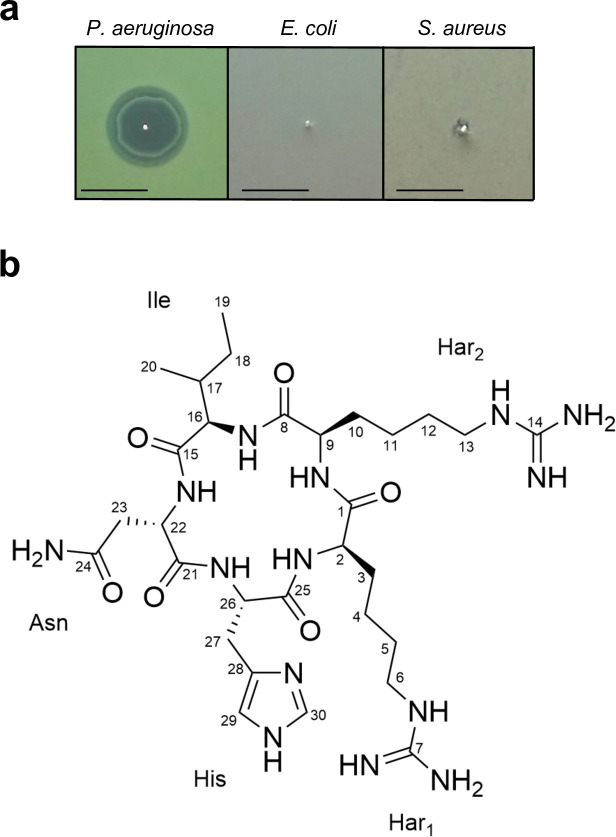 Fig 1
Fig 1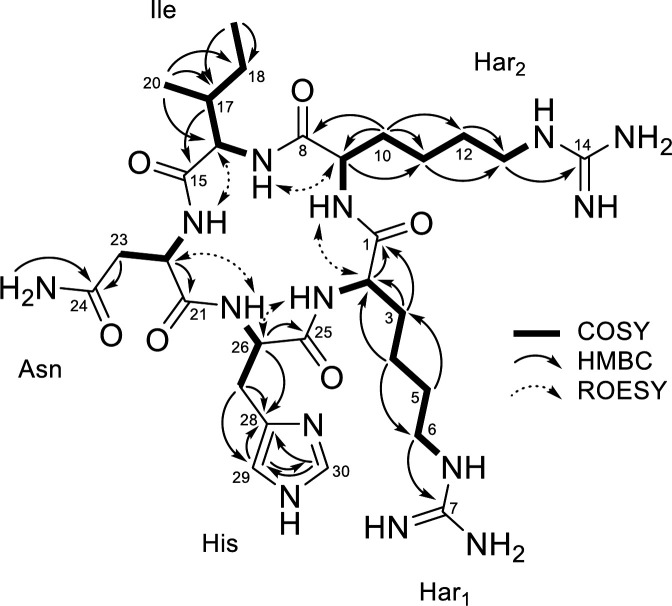 Fig 2
Fig 2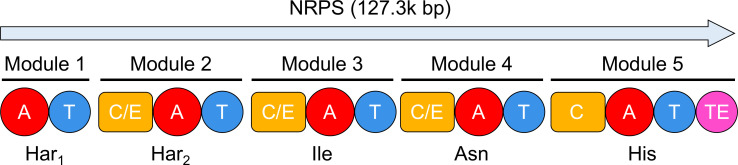 Fig 3
Fig 3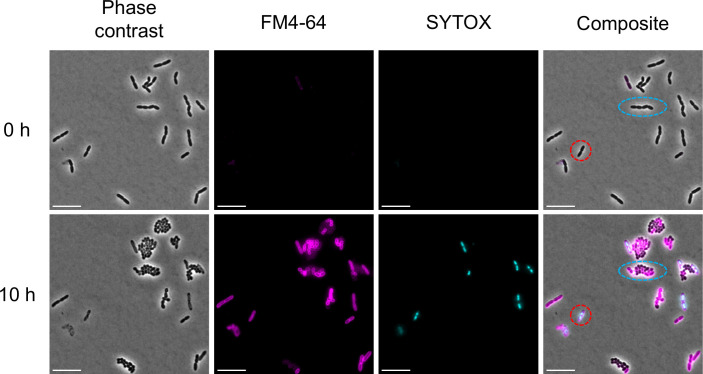 Fig 4
Fig 4| Region | Type | Nucleotide start location | Nucleotide end location | Similarity confidence | Most similar known cluster |
|---|---|---|---|---|---|
| Region 1.1 | NRPS | 1 | 46,048 | ||
| Region 1.2 | NRPS | 95,964 | 223,263 | Low | Viscosinamide A/pseudodesmin A |
| Region 1.3 | NRPS | 362,371 | 472,630 | ||
| Region 1.4 | NRPS | 596,330 | 640,721 | ||
| Region 1.5 | NRPS-like, resorcinol | 1,143,889 | 1,215,080 | High | Minimycin/indigoidine |
| Region 1.6 | Darobactin | 1,231,885 | 1,253,174 | High | Darobactin A |
| Region 1.7 | T1PKS, NRPS | 1,355,250 | 1,404,272 | ||
| Region 1.8 | NRPS, NRPS-like | 1,424,165 | 1,453,311 | ||
| Region 2.1 | T1PKS | 220,586 | 265,727 | ||
| Region 2.2 | Betalactone | 340,106 | 365,596 | ||
| Region 2.3 | Terpene-precursor | 521,928 | 542,845 | ||
| Region 2.4 | NRPS | 558,658 | 601,936 | ||
| Region 2.5 | NRPS, RRE-containing | 802,212 | 879,939 | Low | Gamexpeptide A, B, E/luminmide B, D, E, F, G |
| Region 3.1 | Azole-containing-RiPP | 179,457 | 205,555 | ||
| Region 3.2 | Phosphonate | 340,142 | 359,150 | Low | Luminmycin A/glidobactin A/cepafungin |
| Region 4.1 | PUFA, hglE-KS, NRP-metallophore, NRPS | 80,472 | 192,641 | High | Photobactin |
| Region 4.2 | NRPS | 273,698 | 348,474 | ||
| Region 5.1 | RiPP-like | 7,196 | 18,119 | ||
| Region 5.2 | NRPS | 105,837 | 183,376 | High | Pyrrolizixenamide A |
| Region 5.3 | CDPS | 259,772 | 280,479 | ||
| Region 7.1 | NRPS | 173,038 | 220,182 | ||
| Region 8.1 | NRPS | 73 | 105,947 | ||
| Region 11.1 | Leupeptin | 32,661 | 58,419 | High | Leupeptin A, B, C, D |
| Region 14.1 | Terpene | 35,136 | 60,745 | High | Carotenoid |
| Region 15.1 | NRPS | 15,781 | 84,883 | ||
| Region 19.1 | NRPS | 16,742 | 49,564 | ||
| Region 23.1 | NRPS, NRPS-like | 1 | 18,754 | High | Xenematide |
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —JSPS KAKENHI
- —The Hokuto Bio-science Promotion Foundation
- —The Nakajima Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEntomopathogenic Microorganisms in Pest Control · Insect symbiosis and bacterial influences · Insect and Pesticide Research
INTRODUCTION
We are facing an antibiotic resistance crisis (1). Pseudomonas aeruginosa, a Gram-negative opportunistic pathogen, possesses a well-developed multidrug efflux pump system and an outer membrane that serves as a barrier against large molecules. These characteristics make P. aeruginosa one of the most challenging pathogens to treat with antibiotics, especially in cases of multidrug-resistant (MDR) P. aeruginosa infections. In 2024, the World Health Organization classified P. aeruginosa as a “High Priority” pathogen that requires new antibiotics (2, 3). Moreover, according to the CDC’s Antibiotic Resistance Threats in the United States 2019, there were 32,600 cases of infection in hospitalized patients, and 2,700 cases resulted in deaths due to MDR P. aeruginosa (4).
The entomopathogenic bacterium Photorhabdus species is known to produce various bioactive compounds, including antibiotics (5). However, no Photorhabdus-derived antibiotics are currently in practical use. Narrow-spectrum antibiotics are preferable for drug development because they reduce the risk of generating drug-resistant mutants in off-target bacteria and minimize disruption of the gut microbiome, which includes beneficial bacteria, such as Lactobacillus and Bifidobacterium. P. aeruginosa is primarily associated with respiratory and systemic infections. However, systematically or orally administered antibiotics distribute throughout the body, impacting the gut microbiota. Therefore, the development of antibiotics with selective activity against P. aeruginosa is essential to mitigate collateral damage to commensal microbiota. We recently introduced the differential screening for antibiotic discovery, which compares the activity of compounds in bacterial culture supernatants against several pathogens to identify compounds with selective activity against specific pathogens (6–8). By applying differential screening with Photorhabdus strains as a screening source, we discovered several antibiotics that exhibit selective activity against Gram-negative pathogens. For instance, darobactins and dynobactins, novel classes of antibiotics discovered from Photorhabdus khanii and Photorhabdus australis, respectively, selectively kill Gram-negative pathogens by inhibiting BamA, a protein involved in inserting nascent outer membrane proteins into the outer membrane (9–11). 3′-Amino-3′-deoxyguanosine (ADG), which was discovered from Photorhabdus luminescens, acts as a prodrug that mimics GTP and selectively kills specific Gram-negative bacteria, such as Escherichia coli and Klebsiella pneumoniae, by interrupting transcription (12). More recently, we rediscovered 3,6-dihydroxy-1,2-benzisoxazole (DHB) from the culture supernatant of Photorhabdus laumondii (13). DHB is a known antibiotic with broad activity against Gram-negative bacteria that binds to 4-hydroxybenzoate octaprenyltransferase and inhibits the ubiquinone biosynthesis pathway. These discoveries emphasize the potential of Photorhabdus strains as a source of diverse anti-Gram-negative antibiotics and indicate that the combination of differential screening and Photorhabdus strains is a promising strategy for discovering novel antibiotics with pathogen-selective activity. Furthermore, most Photorhabdus strains harbor more than 20 biosynthetic gene clusters (BGCs) in their genome, with the majority of these clusters encoding unknown compounds (14). We hypothesized that Photorhabdus strains could produce additional antibiotics with selective activity against Gram-negative bacteria beyond darobactins, dynobactins, ADG, and DHB. In this study, we report the discovery of pioamide, a novel cyclic pentapeptide antibiotic with selective activity against P. aeruginosa. Pioamide exhibits narrow-spectrum activity and likely acts on the cell surface of P. aeruginosa or is taken up through a species-specific uptake mechanism.
MATERIALS AND METHODS
Isolation of pioamide
P. khanii HGB1456 was cultured in 2-L Erlenmeyer flasks containing 1 L tryptic soy broth at 28°C with shaking at 200 rpm for 10–14 days. After the culture, the bacterial cells were removed by centrifugation at 8,000 × g for 10 min, and the resulting culture supernatant was semipurified using XAD16N resin (20–60 mesh, Sigma-Aldrich), followed by cation-exchange chromatography (SP Sepharose XL, GE Healthcare), as described previously (9). The active fraction was eluted with 50 mM ammonium acetate buffer (pH 7.0). A 4-mL sample of the highly concentrated semipurified extract, corresponding to 500 mL to 1 L of the culture supernatant, was subjected to reverse-phase high-performance liquid chromatography (HPLC) using a C18 column (Luna 5 µm C18(2) 100 Å, LC column 250 × 21.2 mm, Phenomenex) under the following HPLC conditions: solvent A, Milli-Q water with 0.1% (vol/vol) formic acid (FA); solvent B, acetonitrile (ACN) with 0.1% (vol/vol) FA. The initial concentration of 2% solvent B was maintained for 7 min, followed by a linear gradient to 40% over 23 min at a flow rate of 9 mL/min. UV detection was performed at 254 nm, and fractions were collected every 20 s. The active fractions were eluted between 17 and 19 min. Active fractions from the first HPLC step were further purified by reverse-phase HPLC using a C18 column (XBridge, BEH C18 OBD column, 100 Å, 5 μm, 250 mm × 4.6 mm, Waters) under the following HPLC conditions: solvent A, 50 mM ammonium acetate (pH 8.0); solvent B, ACN. The initial concentration of 2% solvent B was maintained for 2 min, followed by a linear gradient to 95% solvent B over 20 min at a flow rate of 1 mL/min. UV detection was performed at 254 nm. The active compound was eluted at 8.5 min. The purified active fraction was subjected to a final round of purification by reverse-phase HPLC on a C18 column (XBridge, BEH C18 OBD column, 100 Å, 5 μm, 250 mm × 4.6 mm, Waters) under the following HPLC conditions: solvent A, Milli-Q water with 0.1% (vol/vol) FA; solvent B, ACN with 0.1% (vol/vol) FA. The initial concentration of 2% solvent B was maintained for 2 min, followed by a linear gradient to 95% solvent B over 20 min at a flow rate of 1 mL/min. UV detection was performed at 280 nm. The pure active compound was eluted at 7 min.
The activity of HPLC fractions and pioamide was determined by a diffusion assay on agar plates prepared as follows (15). Exponentially growing cultures of P. aeruginosa PAO1, E. coli MG1655, and Staphylococcus aureus HG003 (OD_600_ 0.1–0.9) were diluted to OD_600_ 0.03 in Mueller Hinton II Broth (MHIIB) and then used to cover Mueller Hinton II agar (MHIIA) plates. Excess culture was removed, the plates were dried on a clean bench, and the HPLC fractions or pioamide were spotted onto the MHIIA plates. The plates were incubated overnight at 37°C, and antibacterial activity was evaluated by observing the zone of inhibition around each spot.
Elucidation of structure
The 1D ^13^C nuclear magnetic resonance (NMR) spectra were acquired on a Bruker AVANCE II 400 equipped with a 5 mm BBFO probe, and all other NMR data were recorded on a Bruker AVANCE II 700 MHz NMR spectrometer equipped with a 5 mm TXI probe. Pioamide was prepared in 500 µL of DMSO-d6. The spectra were obtained at 300 K, including ^1^H (zg), ^13^C (zgpg30), COSY (cosygpmfqf), TOCSY (dipsi2etgpsi), ^1^H–^15^N HSQC (hsqcetf3gpsi), ^1^H–^13^C HSQC (hsqcedetgpsisp2.3), ^1^H–^13^C HMBC (hmbcgplpndprqf), and ROESY (roesyphpp.2). High-resolution mass spectrometry analysis was performed using a Thermo Scientific LTQ-Orbitrap mass spectrometer equipped with an electrospray ionization (ESI) source in positive ion mode (FTMS ESI positive mode, full scan MS range: m/z 150–2,000). The compound was introduced via syringe infusion. The spray solvent was a binary mixture of water and acetonitrile (80:20, vol/vol) containing 0.1% FA.
Minimum inhibitory concentration and cytotoxicity assays
The minimum inhibitory concentration (MIC) and cytotoxicity were determined using microbroth dilution as described previously (9). Overnight cultures of P. aeruginosa strains, E. coli MG1655, Acinetobacter baumannii ATCC17978, K. pneumoniae ATCC700603, and S. aureus HG003 were diluted 1:100 in MHIIB and incubated at 37°C with aeration at 220 rpm. Regarding the P. aeruginosa PΔ6-Pore mutant, the strain was cultured in the presence of 30 µg/mL gentamicin, and the expression of porin was induced by adding 100 µM IPTG in MHIIB. Exponential cultures (OD_600_ 0.1–0.9) were diluted to OD_600_ 0.001 (approximately 5 × 10^5^ CFUs/mL) in MHIIB, and 100 µL aliquots were transferred into round-bottomed 96-well plates containing pioamide, which was serially diluted twofold. After overnight incubation at 37°C, the MIC of pioamide was determined as the minimum concentration at which bacterial growth was significantly inhibited. Cytotoxicity was determined using the microplate Alamar Blue assay (MABA/resazurin). Exponentially growing FaDu pharynx squamous cell carcinoma (ATCC HTB-43) and HepG2 liver hepatocellular carcinoma (ATCC HB-8065) cell lines were cultured in Eagle’s minimum essential medium supplemented with 10% fetal bovine serum and then seeded into a 96-well, flat-bottomed, tissue culture plate (Corning), followed by incubation at 37°C with 5% CO_2_. After 24 h, the medium was aspirated and replaced with fresh medium containing the test compounds (twofold serial dilution in 100 µL of media). After 72 h of incubation at 37°C with 5% CO_2_, resazurin (Acros Organics) was added to each well to a final concentration of 0.15 mM. After 3 h, absorbance values were measured at 544 and 590 nm using a BioTek Synergy H1 microplate reader. All MIC and cytotoxicity assays were performed at least in triplicate.
Fluorescence microscopy
For time-lapse microscopy, P. aeruginosa PAO1 was cultured in MHIIB overnight to the stationary phase, and the following day was inoculated into fresh MHIIB at 1:100 and grown for 2 h at 37°C. Cells were diluted 10-fold in MHIIB, placed on top of a 1.5% low-melting-point agarose MHIIB pad containing pioamide (8× MIC), FM4-64 (10 µg/mL), and SYTOX Green (0.5 µM), and observed with a Nikon Ti2-E fluorescence microscope using a 100× oil-immersion objective. The fluorescence signals for SYTOX Green and FM4-64 were collected sequentially by excitation at 480 and 550 nm, respectively, alongside a phase-contrast image. A thermostatic chamber was used to maintain a temperature of 37°C for the duration of the experiment. Images were acquired with NIS-Elements every 30 min at a resolution of 2,048 by 2,048. The images shown in Fig. 4 were processed using an enhanced contrast process, and the HyperStackReg plugin was used to correct for the x–y drift.
Mutation analysis
P. aeruginosa PAO1 cells obtained from an exponential culture were washed with PBS and then inoculated onto MHIIA plates containing an HPLC fraction consisting of four times the MIC of pioamide at a density of 6.6 × 10^8^–7.2 × 10^8^ CFUs/plate. After 2 days of incubation at 37°C, spontaneous pioamide-resistant mutant strains were restreaked onto MHIIA plates containing four times the MIC of pioamide to evaluate the stability of resistance. Experiments were conducted with three independent cultures. Genome sequencing and variant calling were conducted at the Microbial Genome Sequencing Center (MiGS, Pittsburgh). Whole-genome sequencing was performed using paired-end reads (2× 150 bp) on Illumina NextSeq 550, and variant calling was conducted using the P. aeruginosa genome reference (NCBI: GCF_000006765.1).
RESULTS AND DISCUSSION
Discovery of pioamide from P. khanii culture
P. khanii HGB1456 is a producer of the first member of a new class of antibiotic darobactin, which is encoded by a silent BGC. Genome analysis of P. khanii HGB1456 using antiSMASH revealed that this strain harbors 27 BGCs, including the one encoding darobactin (16). Among these, 20 BGCs exhibited low or little similarity to known antibiotics (Table 1). On the basis of this observation, we hypothesized that P. khanii HGB1456 has the potential to produce additional antibiotics that selectively kill Gram-negative bacteria, which might be produced in low amounts or encoded by silent BGCs, as in the case of darobactin. To test this hypothesis, we semi-purified the culture supernatant of P. khanii HGB1456 using the synthetic resin XAD16N, followed by cation-exchange chromatography. After drying the semi-purified sample, it was resuspended in a small volume of Milli-Q water, resulting in a 250-fold concentration compared with that of the original supernatant, enabling the detection of antibiotic activity from silent BGCs or compounds produced in smaller amounts. This highly concentrated sample was subjected to HPLC, and then the eluents were fractionated. Each fraction was tested for antibacterial activity against the representative Gram-negative bacteria E. coli MG1655 and P. aeruginosa PAO1, as well as the Gram-positive bacterium S. aureus HG003 as a counter screening control. We observed that several fractions demonstrated activity against only P. aeruginosa PAO1. Further purification by HPLC combined with bioassay-guided fractionation led to the isolation of a pure active compound that exhibited a distinct zone of inhibition exclusively against P. aeruginosa (Fig. 1a; Fig. S1a). The liquid chromatography with tandem mass spectrometry analysis showed a molecular ion at m/z 705.42 [M+H]^+^ (Fig. S1b). Remarkably, this molecular mass does not match any known compounds listed in the natural product database Antibase. These data suggest that P. khanii HGB1456 produces a novel antibiotic.
Activity and structure of pioamide. (a) Antipseudomonal activity of pioamide. The zone of inhibition indicates its antibacterial activity against P. aeruginosa. Scale bar 1 cm. (b) Structure of pioamide.
Determination of pioamide structure
Pioamide was obtained as a white powder. Its structure was determined by MS spectrometric and NMR spectroscopic analysis (Fig. S2). HRESIMS (m/z 705.4208 [M+H]^+^) combined with the NMR data indicated the molecular formula of pioamide as C_30_H_52_N_14_O_6_ (calculated [M+H]^+^ = 705.4267). The analysis of ^1^H, ^13^C, and HSQC NMR spectra indicated the presence of two methyls (δH/C 0.78/14.8 and 0.82/11.4), nine methylenes (δH 1.02–2.83/25.5–31.3 and 2.41/36.7), one aliphatic methine (δH/C 1.72/37.1), and two nitrogen-bearing methylenes (δH/C 3.01/40.4 and 3.03/40.3) (Table S1). The presence of numerous exchangeable protons (δH 6.92–8.71) and downfield chemical shifts of five methine groups (δH/C 3.88/55.1, 4.08/55.7, 4.08/56.5, 4.31/53.1, and 4.36/52.2) strongly indicated the peptidic nature of the compound consisting of five amino acids. A comprehensive analysis of 2D NMR data (COSY, HMBC, and ROESY) determined the structure of each amino acid (Fig. 2). The proton spin system from the amide proton NH-16 (δH 8.02) to the methyl proton H_3_-19 (δH 0.82) through two methines H-16 and H-17, methyl H_3_-20, and methylene H_2_-18 observed in the COSY spectrum and HMBC correlations from H-17 to C-15 revealed the presence of an isoleucine residue. The asparagine residue was determined using the proton spin system (NH-22 to H_2_-23) together with HMBC correlations from H_2_-23 and NH_2_-24 to the amide carbonyl carbon C-24 (δC 170.7). The presence of two closely located carbonyls, C-7 (δC 157.3) and C-14 (δC 157.3), at characteristic chemical shifts indicated the presence of two guanidinium moieties observed in arginine. Further detailed analysis of COSY correlations revealed two spin systems (NH-2/H-2/H_2_-3/H_2_-4/H_2_-5/H_2_-6/NH-6 and NH-9/H-9/H_2_-10/H_2_-11/H_2_-12/H_2_-13/NH-13) consisting of a 4-carbon aliphatic side chain. Furthermore, the clear HMBC correlations from NH-6 and NH-13 to C-7 and C-14 determined the structure of two homoarginine residues, respectively. The remaining proton spin system (NH-26/H-26/H_2_-27) observed in the COSY spectra was extended to the imidazole unit by the key HMBC correlations from H_2_-27 to olefinic carbons C-29 (δC 117.8) and C-28 (δC 132.2). The HMBC correlation of the remaining olefinic proton H-30 (δH 7.49) to the protonated olefinic carbon C-29 and nonprotonated olefinic carbon C-28 clearly established the histidine residue. Finally, analysis of ROESY data revealed five characteristic correlations between α-methines and the amide protons of the preceding residues. Together with the molecular formula, these data demonstrate that pioamide is a cyclic pentapeptide with the sequence: homoarginine-1, homoarginine-2, isoleucine, asparagine, and histidine.
Key 2D NMR correlations of pioamide in DMSO-d6.
BGC of pioamide
P. khanii HGB1456 harbors 27 distinct BGCs identifiable by the genome accession number WHZZ00000000 using antiSMASH (Table 1). Among these, pioamide, a cyclic pentapeptide compound, is synthesized by a nonribosomal peptide synthetase (NRPS) gene cluster. Photorhabdus strains produce antibiotics that are structurally similar to pioamide, collectively referred to as GameXPeptides, which are common secondary metabolites in Photorhabdus strains and exhibit extensive activity (17, 18). On the basis of this information, we investigated the BGC responsible for pioamide.
Among the identified BGCs, 17 were NRPS-related clusters. Some of these BGCs feature multiple NRPS genes within a singular cluster, suggesting the potential to encode a variety of nonribosomal peptides. Remarkably, the analysis of adenylation (A) domains revealed two distinct NRPS clusters that encode pentapeptides. The BGC for pioamide was deduced based on both the chemical structure of the compound and the genetic composition of the cluster (Fig. 3). This specific NRPS gene cluster is distinguished by the presence of five A domains containing two adjacent arginine residues. This distinctive A domain configuration strongly implies that the gene cluster region 1.2 is responsible for the production of pioamide (Table 1). The predicted amino acid sequence, Arg-Arg-Ile-X-Tyr, aligned partially with the observed pioamide sequence, identified as Har-Har-Ile-Asn-His ([Fig. 1b and 2](#F1 F2)). The stereochemistry of the α carbons was deduced from the presence of three dual condensation/epimerization (C/E) domains in the modules 2, 3, and 4. The dual C/E domains catalyze the epimerization of the initially bound L-residue into the D-configuration and then promote the condensation of the two residues (17, 19). Thus, the stereochemistry of pioamide was deduced as d-Har, d-Har, d-Ile, l-Asn, and l-His.
Predicted biosynthetic gene cluster of pioamide. A, adenylation; T, thiolation; C/E, dual condensation/epimerization; C, condensation; TE, thioesterase.
Activity of pioamide
Pioamide exhibits selective activity against P. aeruginosa PAO1 with a relatively high MIC of 128 µg/mL (Table S2). Although several antibacterials with activity against P. aeruginosa have been reported, only the synthetic compound murepavadin exhibits selective activity against this pathogen (Table S3). Moreover, pioamide exhibited no activity against other bacterial species or human cell lines. Interestingly, we found that the MIC of pioamide against the wild-type strain PAO1 was similar to that against the PΔ6-pore mutant strain, which lacks six efflux pumps (mexAB-oprM, mexCD-oprJ, mexXY, mexJKL, mexEF-oprN, and triABC) and produces porins in its outer membrane, making it highly susceptible to multiple antibiotics (Table S2) (20, 21).
Furthermore, fluorescence microscopy analysis revealed distinct morphological changes in P. aeruginosa cells treated with pioamide. We observed that P. aeruginosa cells treated with pioamide lost their membrane integrity (Fig. 4). Interestingly, several cells underwent multiple divisions, transitioning from the typical rod shape to a spherical form, after which their growth was arrested (Fig. 4). The formation of spherical cells is commonly observed in P. aeruginosa treated with meropenem, an antibiotic that inhibits the synthesis of bacterial cell wall (22, 23). These spherical cells, induced by meropenem treatment, display defective cell walls and outer membranes as a survival strategy (24). In this condition, the target of the antibiotic becomes less accessible or even absent, allowing the cells to survive without acquiring resistance mutations. Our results suggest that P. aeruginosa transitions from the rod shape to spherical form in response to pioamide using a survival mechanism similar to that observed in meropenem-treated P. aeruginosa. These findings support the idea that pioamide activity is not significantly influenced by efflux pumps or porin-mediated permeability. While this may suggest that pioamide acts without entering the cell, it is also possible that it is taken up via specific uptake pathways, such as the self-promoted uptake mechanism previously described for polycationic antibiotics like colistin and aminoglycosides (25). Additionally, P. aeruginosa possesses species-specific outer membrane proteins like OprD, which facilitate carbapenem uptake (26). Other Gram-negative bacteria, such as A. baumannii, have homologs of OprD; however, deletion of A. baumannii oprD does not affect carbapenem resistance (27). Such structural differences in outer membrane proteins may underlie the selectivity of pioamide.
Fluorescence microscopy analysis of P. aeruginosa treated with pioamide. P. aeruginosa PAO1 cells were placed on an agarose pad containing 8× MIC of pioamide and the fluorescent dye FM4-64 (magenta) and SYTOX (light blue) to stain the membrane and demonstrate membrane permeability, respectively. P. aeruginosa PAO1 cells were incubated at 37°C, and growth was monitored by microscopy. The red circle indicates the loss of membrane integrity of cells; light blue indicates spherical cells. Scale bar 10 µm.
To explore the mechanism of resistance to pioamide in P. aeruginosa, we generated spontaneous pioamide-resistant mutant strains from the P. aeruginosa wild-type strain PAO1 by plating cells onto MHIIA plates containing four times the MIC of pioamide. As a result, spontaneous pioamide-resistant mutant strains were obtained with a frequency of 1.6 × 10^−7^–3.7 × 10^−7^. Whole-genome sequencing of the three mutants revealed mutations in pmrB (pmrB L18P, pmrB D47E, and pmrB c.450_451ins CAGATCTGGATCAGCGAA), which encodes a two-component regulatory system signal sensor kinase (Table S4). It is well established that specific mutations in pmrB confer resistance to polymyxin B and colistin, which target lipopolysaccharide (LPS) in Gram-negative bacteria and cause cell lysis. The resistance arises through the modification of lipid A by the addition of L-Ara4N, which reduces the affinity of lipid A for these antibiotics (28). Fluorescence microscopy analysis of P. aeruginosa treated with colistin revealed immediate cell lysis, with no visible cells in the microscopic field when an agarose pad was applied (data not shown). This finding suggests that pmrB mutations confer P. aeruginosa with resistance to both pioamide and colistin, although the mechanisms of action of these antibiotics are distinct. A recent study demonstrated that pmrB mutations confer cross-resistance between colistin and murepavadin (POL7080) in P. aeruginosa (29, 30). Murepavadin is an outer membrane-targeting peptide that specifically interacts with the LPS transporter protein LptD located on the cell surface and kills P. aeruginosa through a nonlytic mechanism. It has been suggested that modified LPS causes reduced murepavadin binding to LptD. It is more likely that pioamide resistance in P. aeruginosa is due to pmrB mutations inhibiting the interaction between pioamide and its target, as observed in the case of murepavadin, rather than PmrB itself being the target.
Although our study revealed important insights into the activity of pioamide and its resistance mechanisms, several questions remain unanswered due to the limited amount of pioamide currently available. Given that the resistance mechanism involves pmrB, which regulates LPS modifications in response to environmental stress, it is possible that the MIC of pioamide varies depending on the culture medium conditions (31). In a related context, it remains unclear whether the activity of pioamide is influenced by divalent cations, such as Ca^2+^ and Mg^2+^, which are known to affect the outer membrane integrity and antibiotic susceptibility of Gram-negative bacteria (32, 33). Therefore, MIC assays under different ionic conditions, such as in the presence or absence of divalent cations, or in minimal media like M9-MOPS, may provide further insight into the activity of pioamide. Further studies addressing these points will deepen our understanding of pioamide and contribute to the development of novel selective antibiotics against P. aeruginosa. Although pioamide exhibits limited potency, considering its peptidic nature, chemical derivatization is anticipated to enhance its activity (34). Remarkably, pioamide exhibited highly selective activity against P. aeruginosa, warranting further research.
To date, murepavadin is the only P. aeruginosa-selective antibacterial agent. In addition, the most effective, approved antibiotics active against P. aeruginosa, including recently reported compounds, are broad-spectrum agents, emphasizing the rarity of P. aeruginosa-selective antibiotics (Table S3) (9, 11, 29, 35–37). Considering the intrinsic resistance of P. aeruginosa to multiple antibiotics, such selective activity is both unusual and puzzling. Further research into the mechanism of action of pioamide may expand our understanding of potential targets for P. aeruginosa-selective therapeutics. Our study findings reinforce that Photorhabdus species are prolific producers of antibiotics with novel modes of action. Therefore, these bacterial species may remain a rich source for the discovery of new antibiotics effective against Gram-negative pathogens.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Naghavi M, Vollset SE, Ikuta KS, Swetschinski LR, Gray AP, Wool EE, Robles Aguilar G, Mestrovic T, Smith G, Han C, et al.. 2024. Global burden of bacterial antimicrobial resistance 1990–2021: a systematic analysis with forecasts to 2050. Lancet 404:1199–1226. doi:10.1016/S 0140-6736(24)01867-139299261 PMC 11718157 · doi ↗ · pubmed ↗
- 2Sati H, Carrara E, Savoldi A, Hansen P, Garlasco J, Campagnaro E, Boccia S, Castillo-Polo JA, Magrini E, Garcia-Vello P, Wool E, Gigante V, Duffy E, Cassini A, Huttner B, Pardo PR, Naghavi M, Mirzayev F, Zignol M, Cameron A, Tacconelli E, WHO Bacterial Priority Pathogens List Advisory Group. 2025. The WHO Bacterial Priority Pathogens List 2024: a prioritisation study to guide research, development, and public health strategies against antimicrobial resistance. Lancet Infect Dis 25:1033–1043. doi · doi ↗ · pubmed ↗
- 3WHO. 2025. WHO Bacterial Priority Pathogens List, 2024: bacterial pathogens of public health importance to guide research, development and strategies to prevent and control antimicrobial resistance. Available from: https://www.who.int/publications/i/item/978924009346110.1016/S 1473-3099(25)00118-5PMC 1236759340245910 · doi ↗ · pubmed ↗
- 4Centers for Disease Control and Prevention (CDC). 2019. Antibiotic Resistance Threats in the United States, 2019. Available from: https://www.cdc.gov/antimicrobial-resistance/data-research/threats/index.html
- 5Tobias NJ, Wolff H, Djahanschiri B, Grundmann F, Kronenwerth M, Shi Y-M, Simonyi S, Grün P, Shapiro-Ilan D, Pidot SJ, Stinear TP, Ebersberger I, Bode HB. 2017. Natural product diversity associated with the nematode symbionts Photorhabdus and Xenorhabdus. Nat Microbiol 2:1676–1685. doi:10.1038/s 41564-017-0039-928993611 · doi ↗ · pubmed ↗
- 6Lewis K. 2020. The science of antibiotic discovery. Cell 181:29–45. doi:10.1016/j.cell.2020.02.05632197064 · doi ↗ · pubmed ↗
- 7Leimer N, Wu X, Imai Y, Morrissette M, Pitt N, Favre-Godal Q, Iinishi A, Jain S, Caboni M, Leus IV, et al.. 2021. A selective antibiotic for Lyme disease. Cell 184:5405–5418. doi:10.1016/j.cell.2021.09.01134619078 PMC 8526400 · doi ↗ · pubmed ↗
- 8Imai Y, Hauk G, Quigley J, Liang LB, Son S, Ghiglieri M, Gates MF, Morrissette M, Shahsavari N, Niles S, Baldisseri D, Honrao C, Ma XY, Guo JJ, Berger JM, Lewis K. 2022. Evybactin is a DNA gyrase inhibitor that selectively kills Mycobacterium tuberculosis. Nat Chem Biol 18:1236–1244. doi:10.1038/s 41589-022-01102-735996001 PMC 9844538 · doi ↗ · pubmed ↗