Primary production by the purple nonsulfur bacterium Rhodopila globiformis in an acidic, moderately sulfidic warm spring
Kristopher M. Fecteau, Katelyn M. Weeks, R. Vincent Debes, Tanner J. Barnes, Kirtland J. Robinson, Joshua J. Nye, Melody R. Lindsay, Eric S. Boyd, Everett L. Shock

TL;DR
This study shows that Rhodopila globiformis, a purple nonsulfur bacterium, performs significant primary production in acidic hot springs by assimilating dissolved CO2 through photoautotrophy.
Contribution
The paper reports the first observation of light-driven CO2 assimilation in the most acidophilic anaerobic anoxygenic phototroph known.
Findings
R. globiformis assimilated dissolved inorganic carbon at rates comparable to acidophilic algae.
Acetate assimilation was much lower and unaffected by light, suggesting no photoheterotrophic use of acetate.
The spring conditions are rare, resulting from shallow mixing of groundwater and CO2-rich volcanic gases.
Abstract
Populations of the acidophilic purple nonsulfur bacterium Rhodopila globiformis were identified in two geographically distinct thermal areas in Yellowstone National Park (Wyoming, USA), as confirmed by 16S rRNA gene sequencing and detection of characteristic methoxylated ketocarotenoids. Microcosm-based carbon uptake assays where oxygenic photosynthesis was excluded via addition of 3-(3,4-dichlorophenyl)-1,1-dimethylurea yielded a light-driven dissolved inorganic carbon (DIC) assimilation rate (7 ± 2 mg C g C−1 h−1) comparable to those of highly productive algal mats in acidic hot springs, suggesting that R. globiformis may be performing photoautotrophy at the time of the assay. Rates of acetate assimilation were more than two orders of magnitude lower than DIC assimilation and did not differ between light and dark treatments, indicating photoheterotrophic use of acetate was not…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
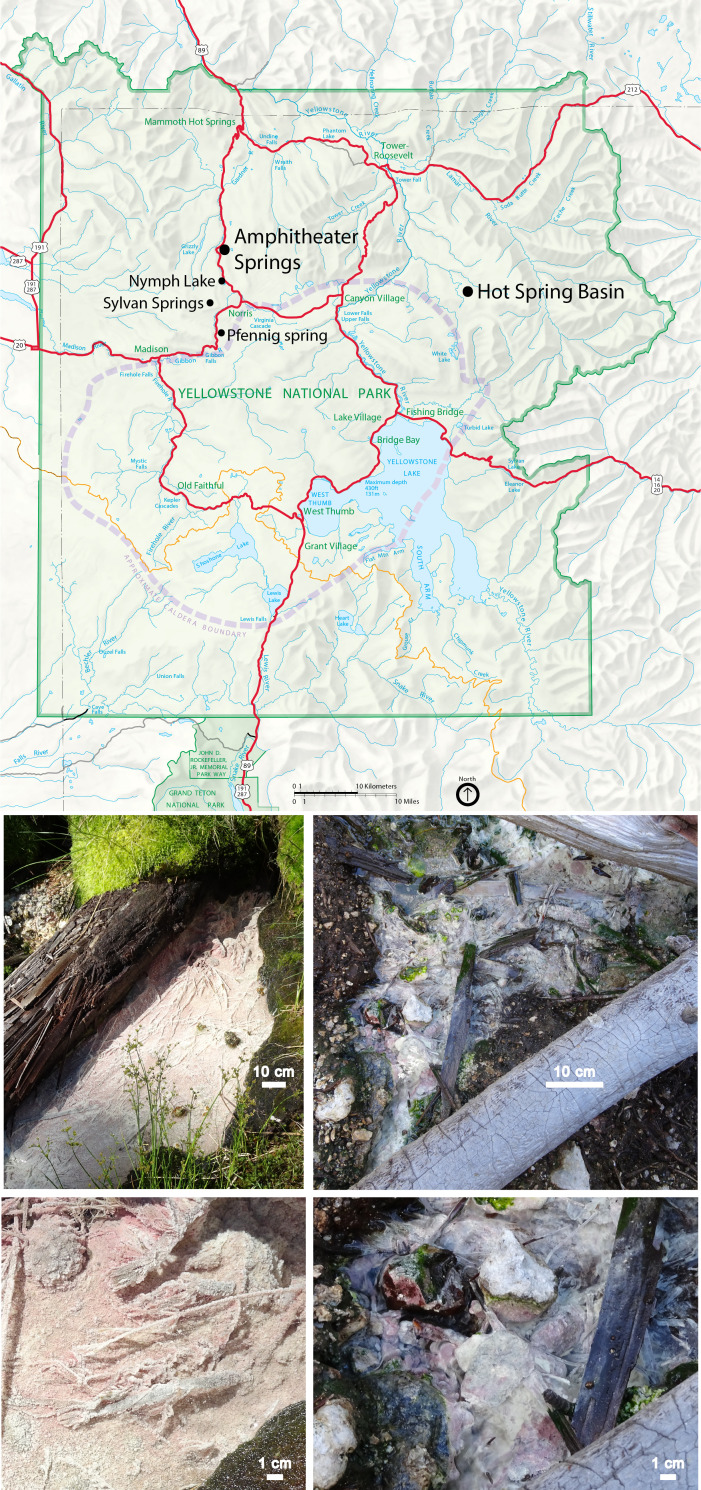 Fig 1
Fig 1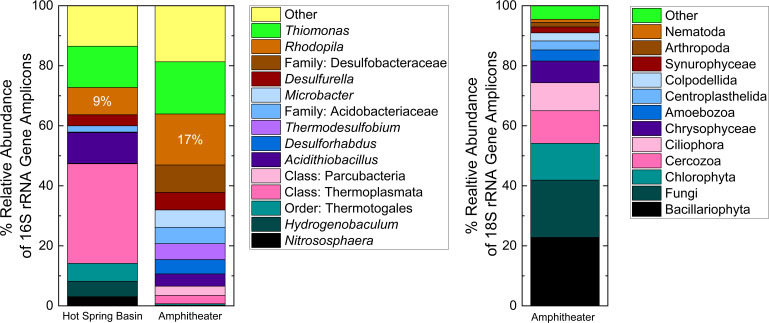 Fig 2
Fig 2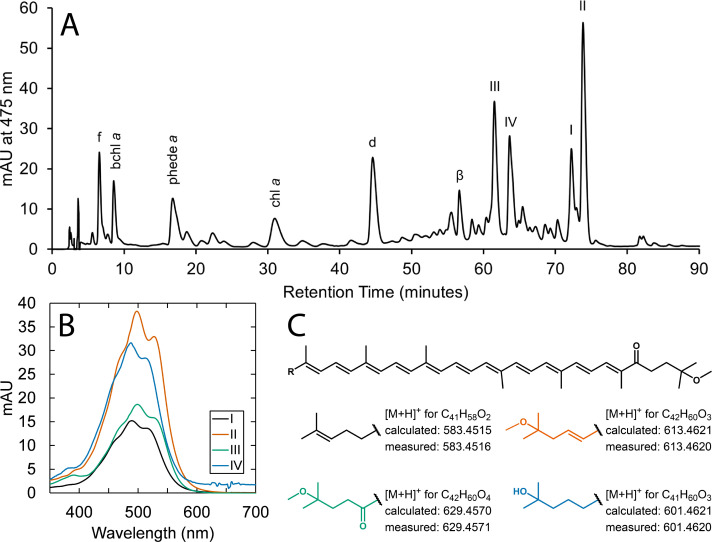 Fig 3
Fig 3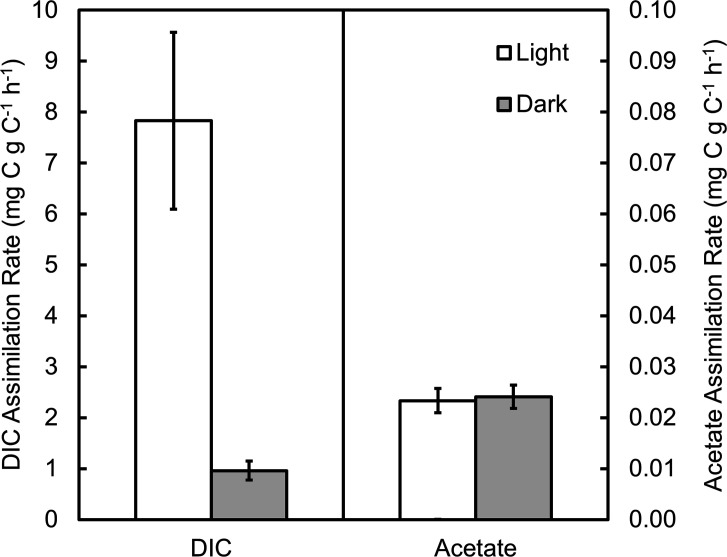 Fig 4
Fig 4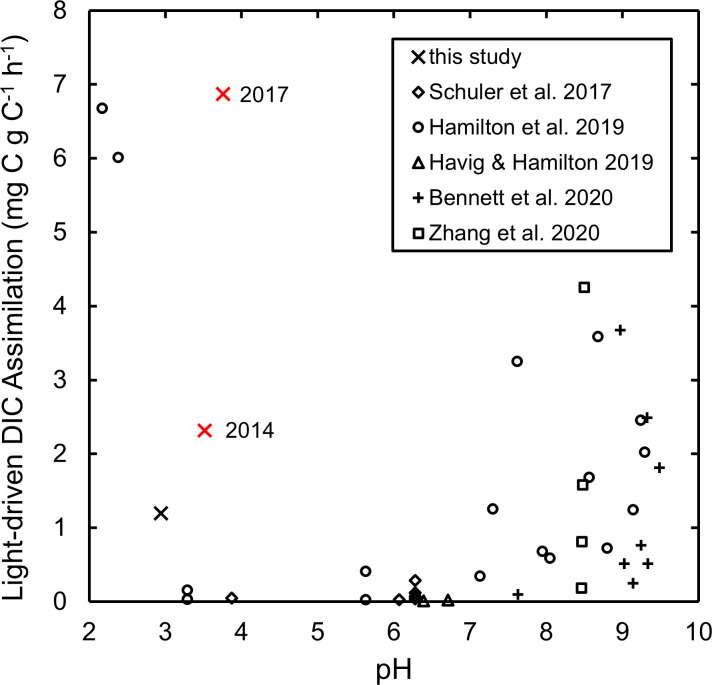 Fig 5
Fig 5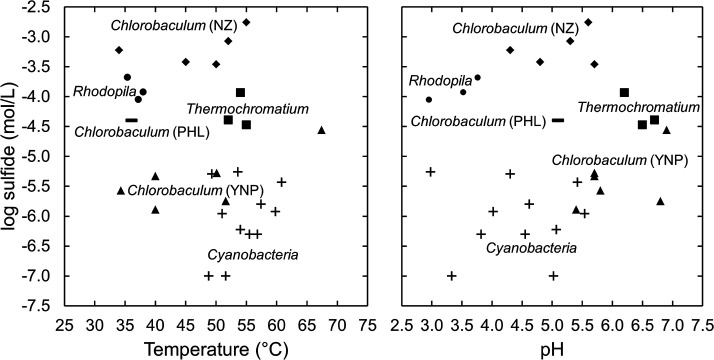 Fig 6
Fig 6| Amphitheater 2017 | Amphitheater 2014 | Lemonade Creek | Hot Spring Basin | |
|---|---|---|---|---|
| Sample ID (YYMMDD) | 170722TO | 140803SK | 170722TP | 170730TQ |
| Easting | 0521755 | 0521755 | 0521760 | 0557919 |
| Northing | 4960245 | 4960245 | 4960228 | 4954198 |
| Elevation, m | 2,289 | 2,289 | 2,295 | 2,563 |
| Temperature, °C | 35.4 | 38.0 | 12.6 | 37.2 |
| Specific conductance, µS/cm | 308.0 | 422.6 | 50.1 | 1,021 |
| pH (field/calculated) | 3.76/3.77 | 3.52/3.55 | 5.28/5.47 | 2.95/2.39 |
| Sulfide, µM | 210 | 120 | 0.65 | 90 |
| Dissolved oxygen, µM | 2.0 | 8.9 | 180 | 16 |
| Silica, mM | 2.6 | 2.5 | 1.0 | 2.9 |
| Fluoride, mM | 0.0031 | 0.0065 | 0.0026 | 0.022 |
| Chloride, mM | 0.079 | 0.10 | 0.016 | 0.021 |
| Bromide, µM | 0.13 | 0.12 | 0.032 | 0.060 |
| Sulfate, mM | 1.0 | 1.3 | 0.058 | 4.2 |
| Nitrate, µM | 0.13 | 0.93 | 0.15 | 0.62 |
| Lithium, µM | 3.1 | 4.0 | 0.17 | 3.0 |
| Sodium, mM | 0.97 | 1.2 | 0.082 | 1.1 |
| Potassium, mM | 0.43 | 0.53 | 0.045 | 0.58 |
| Magnesium, mM | 0.056 | 0.075 | 0.034 | 0.11 |
| Calcium, mM | 0.13 | 0.18 | 0.073 | 0.23 |
| Ammonium, µM | 32 | 34 | 0.41 | 280 |
| Ferrous iron, µM | 57 | 53 | 0.36 | 50 |
| Formate, µM | 2.5 | Not measured | Not measured | 2.3 |
| Acetate, µM | 0.83 | Not measured | Not measured | 0.99 |
| δ2H-H2O, ‰ vs VSMOW | −136.3 ± 0.6 | −140.1 ± 0.3 | −133.0 ± 0.6 | −136.9 ± 0.7 |
| δ18O-H2O, ‰ vs VSMOW | −18.0 ± 0.2 | −18.0 ± 0.1 | −17.7 ± 0.2 | −17.3 ± 0.1 |
| DIC, mM | 11 | 7.9 | 2.1 | 6.0 |
| δ13C-DIC, ‰ vs VPDB | −3.5 ± 0.2 | −4.3 ± 0.2 | −6.4 ± 0.4 | −3.5 ± 0.2 |
| DOC, mM | 0.12 | 0.13 | 0.16 | 0.088 |
| δ13C-DOC, ‰ vs VPDB | −25.4 ± 0.2 | −24.5 ± 0.3 | −26.7 ± 0.2 | −25.4 ± 0.6 |
| Mat C, wt % (dry mass basis) | 9.11 ± 0.07 | 10.63 ± 0.07 | Not measured | 12.0 ± 0.3 |
| δ13C-Mat, ‰ vs VPDB | −26.4 ± 0.1 | −26.1 ± 0.1 | Not measured | −25.6 ± 0.1 |
| Mat N, wt % (dry mass basis) | 1.315 ± 0.007 | 1.355 ± 0.005 | Not measured | 0.58 ± 0.01 |
| δ15N-Mat, ‰ vs air | −2.8 ± 0.1 | −4.9 ± 0.1 | Not measured | −5.9 ± 0.1 |
| Sediment C:N, mol:mol | 8.08 ± 0.07 | 9.15 ± 0.07 | Not measured | 24.1 ± 0.7 |
| Δ13C, ‰ | −22.9 ± 0.2 | −21.8 ± 0.2 | Not measured | −22.1 ± 0.2 |
| Charge imbalance, % | 0.30 | 1.1 | 23 | −69 |
- —National Aeronautics and Space Administrationhttp://dx.doi.org/10.13039/100000104
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Community Ecology and Physiology · Marine and coastal ecosystems · Genomics and Phylogenetic Studies
INTRODUCTION
Among anoxygenic phototrophic bacteria, purple nonsulfur bacteria in particular exhibit great metabolic flexibility, thought to grow best photoheterotrophically under anaerobic conditions but with most species also capable of photoautotrophy or chemoheterotrophic growth in the dark via either respiration or fermentation (1). Some species of purple nonsulfur bacteria may be considered extremophiles, including thermophilic, psychrophilic, alkaliphilic, and acidophilic members (1, 2). However, acidophilic anoxygenic phototrophs appear not to be taxonomically diverse (2, 3). A possible contributing factor to the apparent failure of anoxygenic phototrophs to more extensively radiate into acidic habitats is that extremely acidic habitats did not exist prior to oxygenation of the atmosphere following the evolution of oxygenic photosynthesis (4, 5), hundreds of millions of years after the diversification of anoxygenic phototrophs in anoxic, higher-pH environments. The apparent paucity of acidophily among anoxygenic phototrophic bacteria mirrors that of oxygenic phototrophic bacteria (i.e., Cyanobacteria), which are thought to be excluded from all environments with pH values below 4.0 (6), though the exact pH limit may be lower and difficult to discern (7). Thus, Cyanobacteria have apparently not been able to colonize many acidic environments made possible by their own metabolic production of oxygen. Instead, microbial phototrophy in acidic environments is dominated by algae (7–12). Nevertheless, a variety of acid-tolerant anoxygenic phototrophic bacteria have been characterized that grow aerobically rather than anaerobically, collectively referred to as the aerobic anoxygenic phototrophic bacteria (AAPB). Constituents of the AAPB are not capable of autotrophic growth and may be thought of as chemoheterotrophs that supplement energetic requirements via photophosphorylation (13, 14).
The diversity of anaerobic acidophilic anoxygenic phototrophs is presently limited to four species of purple nonsulfur bacteria across three genera within the Alphaproteobacteria. Strains of Rhodoblastus acidophilus, previously Rhodopseudomonas acidophila (15), were isolated from a variety of environments, including lakes, peat bogs, and swamps, and exhibited a pH minimum for growth of 4.8 and temperature optima of 25°C–30°C (16), though the lower pH limit is likely closer to 4.5 based on further study (17). Another species with similar characteristics, Rhodoblastus sphagnicola, was later isolated from a Sphagnum peat bog (18). Rhodovastum atsumiense was isolated from paddy soil; its temperature optima were found to be 30°C–35°C, and no growth was observed below a pH of 5.0 (19, 20). Rhodopila globiformis, originally Rhodopseudomonas globiformis (21), was isolated from a warm sulfur spring near the Gibbon River in Yellowstone National Park (YNP) in northwest Wyoming and was found to have a lower pH limit for growth of 4.2 and temperature optima of 30°C–35°C (22). Phylogenetically, its sister genus is Acidiphilium, a member of the AAPB found in a variety of acidic habitats, including acid mine drainage (23, 24) and geothermal environments such as YNP (7, 9, 12, 25), and it exists within the Acetobacteraceae, being closely related to the acetic acid bacteria (26).
R. globiformis has received significant attention for some unique biochemical characteristics, which contributed to it being reassigned to its own genus (21, 27). The species synthesizes both a high redox potential iron-sulfur protein (HiPIP) and a soluble cytochrome c_2_, each with particularly high redox potentials (28, 29). The cytochrome c_2_ was found to be similar to cytochromes associated with mitochondria and eukaryotic cytochromes based on its amino acid sequence (30) and crystal structure (31), respectively. The HiPIP has been extensively investigated electrochemically and spectroscopically (32–36), with observations suggesting it is involved in phototrophic electron transfer by reducing cytochromes associated with the reaction center (35). The main pigments of R. globiformis are bacteriochlorophyll a (22, 37) and methoxylated ketocarotenoids that have not been reported to occur in other species (38, 39). The light-harvesting 1-reaction center complex is membrane-bound via an N-terminal α-helix of the cytochrome subunit, rather than with acyl lipids as found in other purple bacteria, and includes polypeptides similar to the γ-polypeptides known only in species synthesizing bacteriochlorophyll b instead of bacteriochlorophyll a (40). R. globiformis produces menaquinone-9 and ubiquinone-9 (41, 42), as well as nearly equal abundances of rhodoquinone-9 and -10, a unique quinone composition when compared to members of the Rhodospirillaceae (43). The lipopolysaccharides were also found to be somewhat different from some other purple nonsulfur bacteria (Rhodospirillium spp.) in both the primary amino sugar (2,3-diaminoglucose) and hydroxyl fatty acid content (44). A novel strain of Rhodopila, likely a new species based on genetic differences, was isolated from a sulfidic spring in Lassen Volcanic National Park in northern California and found to produce 3-methylhopanoids, previously identified only in aerobic species (45). Elsewhere in YNP (Fig. 1), another strain of R. globiformis was isolated from a spring near Nymph Lake (2, 27), and R. globiformis was also detected via 16S rRNA gene sequencing with low relative abundances at two sample locations in the Sylvan Springs thermal area in the Gibbon Geyser Basin, ~3 km northwest of its original source spring (12). Nevertheless, in contrast to its biochemical attributes, there remains little known of its ecology.
Top, map of Yellowstone National Park depicting sample locations from this study and other locations where Rhodopila globiformis has been observed (black points), major roads (red), the continental divide (orange), and the 0.64 Ma caldera boundary (purple), as well as topography and major park locations and water features. The map was derived from the nps.gov website, with selected layers removed in Adobe Illustrator. Bottom, photographs of the Amphitheater site (left) and the Hot Spring Basin site (right). Scale bars are 10 cm (top) and 1 cm (bottom). Photographs were taken during research conducted under research permit YELL-2017-SCI-5434.
Fundamental aspects of the physiology of R. globiformis examined in culture are inconsistent with what is thus far known of its natural habitat. The pH range for growth has been reported to be 4.2–6.5 (22, 27); however, the pH of the spring from which R. globiformis was first isolated was reported to be 3.0 (22) and other springs in the area also putatively harboring R. globiformis mostly have pH values 3–4, with some as low as 2.7 (27). The temperature of the source spring was reported to be approximately 40°C, which is its upper limit for growth in culture; in fact, no growth was observed at 40°C (22, 27). Additionally, sulfide was found to be growth inhibitory at low concentrations, yet the springs where R. globiformis has been found have been described as having large amounts of sulfur, suggesting the presence of sulfide (3, 27). A reduced sulfur source other than sulfide, such as thiosulfate or cysteine, was found to be required for growth (22), but concentrations of amenable sulfur sources are likely to be quite low in acidic springs. Thiosulfate concentrations were found to generally be less than 2 µM in acidic springs in YNP (46), and free amino acid concentrations have been observed at sub-micromolar levels in YNP hot springs (47). It was later determined that sulfate at concentrations below 1 mM could serve as an assimilatory sulfur source, with best growth at 0.1 mM, whereas growth-inhibitory dysregulation of the sulfate assimilation pathway was observed when sulfate exceeded 1 mM (48). Yet, since acidic spring waters in YNP are the result of the oxidation of sulfide to sulfuric acid (49–51), sulfate concentrations would be expected to approach or exceed 1 mM in springs with pH values of ~3. Finally, the organism was found to grow photoheterotrophically with a limited suite of organic compounds (including gluconate, mannitol, and fructose), a characteristic which could prove growth-limiting in nature, as there are likely to be only trace concentrations of specific organic compounds that are able to be assimilated by R. globiformis. However, its genome suggests the capability for metabolizing a more diverse array of organic substrates, suggesting there is a selective advantage for flexibility with respect to substrate usage (52).
Here, we report observations of two warm acidic springs in geographically distinct thermal areas in YNP not previously known to feature populations of R. globiformis. Physicochemical measurements were performed to characterize the habitat of R. globiformis and provide insights into the underlying hydrogeochemical processes resulting in the warm acidic waters of the springs. The microbial communities were investigated with 16S and 18S rRNA gene sequencing and complemented by analyses of phototrophic pigments, including identification of ketocarotenoids specific to R. globiformis. Microcosm-based carbon assimilation assays were conducted in situ to evaluate carbon metabolism in a natural population and related to inferences gleaned from stable isotope ratios of biomass and dissolved inorganic carbon (DIC). Collectively, these observations comprise the first evaluation of the physiological ecology of the most acidophilic anaerobic anoxygenic phototroph presently known.
MATERIALS AND METHODS
Field measurements and sampling
Temperature, pH, and specific conductance (conductivity normalized to 25°C) were measured using portable meters (YSI-30 and WTW 3110) as previously described (53). In 2017, dissolved oxygen was measured optically using a PreSens Fibox 4 meter and a DP-PSt3-L2.5-St10-YOP-HT sensor calibrated to 100°C (PreSens, Regensburg, Germany) as previously described (54); dissolved oxygen was determined colorimetrically in 2014 using the indigo carmine method with AccuVac Ampuls (Method 8316, Hach, Loveland, CO, USA) and a DR2400 spectrophotometer (Hach) according to the manufacturer’s protocol. Total dissolved sulfide was determined via the methylene blue method (Hach method 8131) using Hach reagents and DR2400 and DR1900 spectrophotometers in 2014 and 2017, respectively, on unfiltered water samples diluted with deionized water and analyzed immediately after collection. Total dissolved silica and ferrous iron were determined via the silicomolybdate (Hach method 8185) and the 1,10-phenanthroline (Hach method 8146) methods, respectively, using the same spectrophotometers on 0.2 micron-filtered (Supor membrane, Pall Corporation, Port Washington, NY, USA) water samples. Samples of microbial mat were collected aseptically in 2 mL cryovials and frozen immediately on dry ice and subsequently stored at −80°C until further processing. Reported elevations and Universal Transverse Mercator coordinates (zone 12T) are estimated from USGS 7.5 minute topographical maps and satellite imagery (Google Earth), respectively, as these were deemed more accurate than those obtained on site with a handheld GPS unit.
Filtered (0.2 micron, Supor membrane) water samples for laboratory analyses were collected and stored following previously described procedures (7). Samples for anions and cations were collected in 30 mL high-density polyethylene bottles that had been soaked and rinsed with multiple aliquots of deionized water; bottles for cations were spiked with 6 M methanesulfonic acid, resulting in a final concentration of ~20 mM, in order to prevent volatilization of ammonia or precipitation. These samples were frozen at −20°C as soon as possible after collection and maintained at that temperature until analysis. Samples for organic acid analysis were collected in 20 mL Qorpak amber glass bottles with PTFE cap inserts (55). The bottles were rinsed three times with deionized water, combusted (500°C, 24 hours), then sealed until sampling with the PTFE cap inserts that had been soaked in deionized water for 24 hours and dried. Samples were refrigerated at 4°C upon return to the laboratory until analysis. DIC samples were collected in acid-washed 40 mL amber glass vials and sealed with black butyl rubber septa without any headspace. Unfiltered DIC samples were also collected directly into vials at the sample site using a test tube holder. Dissolved organic carbon (DOC) samples were collected in combusted (450°C, 24 hours) 40 mL amber glass vials spiked with 0.1 mL of 85% phosphoric acid (Thermo Scientific, Waltham, MA, USA) and sealed with Teflon-lined septa without any headspace. Samples for stable isotope ratios of water were collected in 30 mL Qorpak square glass bottles that had been rinsed with deionized water and dried at 100°C (>12 hours); bottles were sealed without any headspace with gas-tight polymer-lined Polycone caps and stored at room temperature (~23°C).
Geochemical analyses
Major anions (F^−^, Cl^−^, Br^−^, SO_4_^−2^, NO_3_^−^) and cations (Li^+^, Na^+^, K^+^, Mg^+2^, Ca^+2^, NH_4_^+^) were determined on separate Dionex DX-600 4 mm ion chromatography systems using suppressed-conductivity detection as previously described (9). Total (carboxylic acid + carboxylate) acetate and formate concentrations were analyzed with the same Dionex DX-600 system used for major anions but equipped with AG-/AS-24A anion exchange columns. Injections of undiluted samples were from an AS40 autosampler using 0.5 mL vials (one injection/vial) onto a 100 µL sample loop. The eluent was held isocratically at 2.5 mM for 20 minutes, after which the eluent concentration was raised to 45 mM over 5 minutes then held at 45 mM for 9 minutes to elute other ions, and finally, the eluent was returned to 2.5 mM over 1 minute and re-equilibrated at 2.5 mM hydroxide for 10 minutes prior to the next injection. The flow rate was constant at 1 mL/minute, and the suppressor current was 112 mA. A linear calibration curve was constructed from a series of solutions prepared using 1,000 µg/mL acetate and formate standards (High Purity Chemicals, Charleston, SC, USA) over the concentration range of 2–750 µg/L (acetate) or 8–750 µg/L (formate). The solutions also contained 2 mg/L fluoride obtained from a 1,000 µg/mL mixed anion standard (Environmental Express) in an effort to match the standard and sample matrices, as acetate elutes on the tail of the fluoride peak.
Analyses of DIC and DOC were performed with an OI Wet Oxidation TOC analyzer coupled to a Thermo Delta Plus Advantage IRMS as described previously (9). Isotope ratios were standardized via analysis of solutions of three glycine working standards: low, δ^13^C = −39.64‰, δ^15^N = 1.35‰; mid, δ^13^C = −8.36‰, δ^15^N = 27.9‰; high, δ^13^C = 15.67‰, δ^15^N = 51.8‰, versus Vienna Pee Dee Belemnite (VPDB) and air, respectively. Stable isotope ratios of water were measured via cavity ring-down spectroscopy using a Los Gatos Research DLT-100 isotope analyzer as previously described (56).
Charge imbalances were calculated using the geochemical speciation code EQ3/6 (57) using activity coefficients calculated via an extended Debye-Hückel equation and equilibrium constants derived from the Helgeson-Kirkham-Flowers equation of state (58, 59). Calculations were performed via the AqEquil Python package (60) accessed from the Water-Organic-Rock-Microbe portal (worm-portal.asu.edu). Charge imbalance is reported as the percent of the mean charge (49).
DNA extraction, PCR, and 16S/18S rRNA gene sequencing
DNA was extracted from thawed mat samples (~0.25 g; excluding Lemonade Creek) using the ZymoBIOMICS DNA miniprep kit with a stated binding capacity of 25 µg according to the manufacturer’s instructions. Excess sample fluid was removed via centrifugation prior to the addition of lysis buffer and bead beating. Extracted DNA was screened for bacterial 16S rRNA genes by polymerase chain reaction (PCR) using 1100F/1492R primers (annealing temperature of 55°C) as previously described (61). Additionally, the same PCR methods were used to screen for the presence of bchY to generate ~500 base pair amplicons using previously described primers and an annealing temperature of 50°C (62).
DNA extracts were sent to the Arizona State University (ASU) Genomics Facility for next-generation sequencing of 16S and 18S rRNA genes using the Earth Microbiome Project primer sets (515F/806R and 1391F/EukB, respectively) and protocol (63–65). Only the DNA extracts from samples collected in 2017 are described herein, with 18S rRNA gene sequencing being performed only for the Amphitheater site. Sequencing was performed on an Illumina-MiSeq (v2, 2 × 250). The FastQ files received from the ASU Genomics Facility underwent quality analysis using FastQC (66). The “.fasta” files were uploaded to QIIME2 (v. 2023.7) for sequence cleaning, trimming, and amplicon sequence variant (ASV) generation using DADA2 (67, 68). The SILVA genes database (v. 138.1) was used for taxonomic classification via the RESCRIPt plug-in available on QIIME2 (69). ASVs that remained inadequately assigned after taxonomic classification via QIIME2 were manually classified using BLASTn via the National Center for Biotechnology Information (NCBI) website (https://blast.ncbi.nlm.nih.gov). The conversion of the sequence counts to percent relative abundances was performed in R (70) using the vegan package (v 2.6-4; 71). Plotting of the percent relative abundances was performed using Origin software (v. 2023 b.).
Pigment analysis
Acetone-soluble pigments were extracted from thawed mat samples collected at the Amphitheater spring in 2017 using bead beating in multiple aliquots of 7:2 (vol:vol) acetone:methanol as previously described (7). The pooled extract was analyzed using liquid chromatography-diode array detection-tandem mass spectrometry (LC-DAD-MS/MS) on an Agilent 1290 high-performance liquid chromatography (HPLC) system coupled to an Agilent 6530 accurate mass quadrupole-time of flight mass spectrometer (Agilent Technologies, Santa Clara, CA, USA). Fifty microliters of sample were injected onto a YMC Carotenoid C30 reverse-phase column (3 × 250 mm × 5 µm particle size; YMC, Kyoto, Japan) and eluted at a constant flow rate of 0.425 mL/minute using a solvent system modified after that of Sander et al. (72). The eluent was initially isocratic with 100% solvent A (81:15:4 methanol:methyl tert-butyl ether:water) for 30 minutes followed by a linear gradient to 100% solvent B (6:90:4 methanol:methyl tert-butyl ether:water) at 90 minutes. At that time, the eluent was returned to 100% A over 1 minute and then re-equilibrated for 19 minutes. The eluent system was adapted for use with an HPLC system equipped with a binary pump by preparing the eluent endmember compositions to construct the gradient; the solvent B mixture was biphasic, with the top, organic-rich phase being used for chromatography. Pigments were monitored via the diode array detector (Agilent 1260) at 360, 475, and 665 nm and spectra were collected from 325–800 nm at 10 Hz. The eluate was then directed into the mass spectrometer and ionized using atmospheric pressure chemical ionization in positive ionization mode with parameters based on those of van Breemen et al. (73). Specific ionization conditions were a corona current of 8 µA, vaporizer temperature of 350°C, ionization gas flow rate of 10 L/minute with a temperature of 325°C, and nebulizer gas pressure of 45 psi. Mass spectra were collected from 200 to 1,500 m/z at a scan rate of 1 Hz. Data-dependent MS/MS spectra were obtained for the most abundant ion in each cycle (if >0.01% relative abundance and >200 counts) at fixed collision energies of 15, 30, or 45 V with argon as the collision gas. Mass accuracy was verified prior to analysis, and accurate mass (<2 ppm) was maintained by infusion of an internal reference mass solution for mass correction over the course of the chromatographic analysis.
Carbon assimilation assays and solid-phase carbon/nitrogen analyses
Rates of biological DIC assimilation and acetate assimilation and oxidation to DIC (i.e., dissimilation or mineralization) were evaluated using microcosms spiked with ^14^C-enriched substrates with methods modified from those previously reported (10, 74). In the laboratory prior to field work, 20 mL serum bottles were crimp-sealed with butyl rubber septa and flushed extensively with ultra-high purity nitrogen. In the field, the bottles were equilibrated to the lower local atmospheric pressure by removal of excess nitrogen via a plastic syringe and 16-gauge needle. Spring water was collected using a 160 mL syringe and aliquoted to a 10 mL syringe via a 3-way stopcock to minimize gas exchange prior to injection of 10 mL of spring water into each bottle. Benthic biomass/sulfur and spring water were collected aseptically in a 50 mL Falcon tube, and the tube was shaken vigorously to yield a slurry. To prepare microcosms, 1 mL of slurry was injected via a 1 mL plastic syringe and 16-gauge needle into each bottle. A set of microcosms designated as killed controls was spiked with 0.1 mL of 50 mM HgCl_2_. A separate set of microcosms was wrapped in aluminum foil to evaluate light-independent rates. In 2017, all microcosms for the DIC assay were spiked with 0.1 mL of 1 mM 3-(3,4-dichlorophenyl)-1,1-dimethylurea (DCMU) in 50% ethanol (vol/vol) to exclude photosystem II-dependent (i.e., oxygenic) photosynthesis; in 2014, DCMU was not employed, nor was it employed in acetate assays. All assays (light, dark, and killed) were conducted in triplicate. To initiate the assays, microcosms were injected with 0.2 mL solution of either NaH^14^CO_3_ (54 mCi/mmol) or Na^14^CH_3_^14^COO (110 mCi/mmol), resulting in final activities of 5.3 µCi and 0.71 µCi per vial, respectively. Microcosms were incubated in the source of the spring for 60 minutes, then immediately placed in the dark on dry ice to terminate the assay and subsequently stored at −20°C until further processing.
Aliquots of remaining slurry (1 mL; n = 6) were collected in 2 mL cryovials, frozen on dry ice in the field, and stored at −20°C until processing. Thawed aliquots were individually collected on pre-weighed plastic weighing boats, dried at ~80°C for 3 days, and then weighed to determine the dry mass added to each microcosm. The dried biomass was then pooled, homogenized with an agate mortar and pestle, and milligram quantities were aliquoted into tin capsules using a microbalance. Sealed capsules were analyzed for C and N contents and stable isotope ratios using an ECS 4010 Elemental Analyzer (Costech Analytical Technologies, Valencia, CA, USA) coupled to a Thermo Delta Plus Advantage isotope ratio mass spectrometer (IRMS). The resulting CO_2_ and N_2_ were separated via gas chromatography, and their molecular ions were monitored as ion chromatograms at 44 m/z and 28 m/z, respectively, with peak areas related to elemental abundances via linear calibration curves using NIST 2710 (Montana soil). Carbon isotope ratios were standardized using the same glycine working standards employed for DIC and DOC analyses. Carbon and nitrogen analysis for the Hot Spring Basin site was also completed using the same methods with a thawed mat sample aliquot.
For analysis, microcosms were allowed to thaw and then injected with 1 mL of 1 M HCl to volatilize DIC. Vials for DIC assimilation assays were unsealed and allowed to equilibrate in a fume hood dedicated to radioisotope work for >2 hours, whereas acetate assay vials remained sealed during equilibration (also >2 hours). To evaluate rates of acetate dissimilation (oxidation of acetate to DIC), 5 mL of headspace in each acetate assay vial was removed via a 10 mL syringe and stopcock and then injected into an evacuated 12 mL serum bottle containing 1 mL of Carbo-Sorb E (Perkin-Elmer, Santa Clara, CA, USA) and allowed to equilibrate at 4°C for 16–24 hours. The Carbo-Sorb E solution was then transferred to scintillation vials for liquid scintillation counting (described below). To estimate the fraction of the headspace removed for this assay, the total volume of sealed 20 mL bottles was estimated by comparing the mass of bottles filled with deionized water and sealed without headspace versus the initial combined mass of each individual empty bottle, cap, and septum and using the density of pure water at the laboratory temperature to calculate the associated volume. The total volume of the headspace after expansion into the syringe was then calculated by subtraction of the known volumes of liquids/solids added to each assay vial. The calculated fraction of the 5 mL sample (0.25) was subsequently employed in the calculation of the dissimilation rates (see below).
After equilibration, the contents of the microcosm vials were filtered onto 0.2 µm pore size, 25 mm diameter Nucleopore polycarbonate membranes (Whatman, Florham Park, NJ, USA). The filters were washed with ~10 mL of deionized water and dried in scintillation vials at ~80°C for ~16 hours. Ten milliliters of Cytoscint ES scintillation cocktail (MP Biomedicals, Irvine, CA, USA) were added to each scintillation vial (both those containing dried filters and those containing Carbo-Sorb E), ensuring that filters were completely submerged in the fluid. Radioactivity associated with each vial was measured using a LS 6500 liquid scintillation counter (Beckman Coulter, Indianapolis, IN, USA), with disintegrations per minute (DPM) being calculated from counts per minute via a quench curve. Substrate conversion rates were then calculated as
where DPM_T_ is the mean measured DPM in each treatment, DPM_K_ is the mean DPM in the associated killed control, mmol_C_ is the total (labeled plus native) amount of substrate in each microcosm, MC is the molar mass of carbon, νc is the number of moles of carbon per mole of substrate, α is the isotopic discrimination factor (1.05; 75), DPM_added_ is the amount of radioactivity added to each microcosm, gdw is the dry mass of mat material in each microcosm in grams, t is the duration of the assay in hours, and Fs is the fraction sampled for measurement (1 for assimilation rates, 0.25 for dissimilation rates, see above). Uncertainties in each variable above were propagated through the calculation and reflected in the reported uncertainties in the rate, though the largest source of uncertainty is the experimental variation of the microcosm replicates. To summarize, the rate is the fraction of radioactive substrate converted, scaled to the total amount of substrate in each assay, normalized to experimental duration and solid-phase carbon (as a proxy for biomass amount). While substrate transformation in the microcosms presumably follows first-order kinetics (i.e., the “bottle effect”), the rates are approximated using zero-order kinetics given the extremely small extents of substrate conversion over the short duration of the experiment.
RESULTS AND DISCUSSION
Field sites
Two acidic, warm springs with flocculent pink and yellow mats, thought to represent benthic suspensions of Rhodopila globiformis cells associated with precipitated sulfur, were located in geographically distinct thermal areas in YNP (Fig. 1). The primary study site is located in Amphitheater Springs, a thermal area along the Norris-Mammoth corridor outside of the 0.64 Ma caldera ~5 km north of Nymph Lake, where other springs hosting R. globiformis have been reported, and ~13 km north from where it was originally isolated (2, 27). The spring is located in the extreme southern portion of the thermal tract along Lemonade Creek, the primary drainage of the basin, and has a distinct source physically isolated from the creek, into which its effluent flows. There are also locations nearby this spring in the creek bottom where bubbling gas and similar pink and yellow mats are evident. Upstream of the spring, the physicochemical data provided herein (Table 1) indicate Lemonade Creek has received minimal hydrothermal influence, whereas downstream in the extreme northern portion of the basin, its waters consist primarily of the hot (>60°C) effluents from several acidic, sulfur-depositing, high-discharge springs located along and near the creek (76). Downstream of this input, once the creek waters have cooled to below the upper temperature limit for photosynthesis under acidic conditions (~56°C; 77), extensive mats of the acidophilic alga Cyanidioschyzon merolae cover the creek bed (78–82). The second site is located in Hot Spring Basin, an extensive, remote, gas-rich thermal area in the Mirror Plateau region, also outside of the 0.64 Ma caldera (83–86).
Community composition
Amplicons for bchY, the gene encoding the Y subunit of chlorophyllide oxidoreductase, the first enzyme in the chlorophyll biosynthetic pathway specific for bacteriochlorophylls, and therefore, a genetic marker for anoxygenic phototrophs (62), were obtained from DNA extracted from both sample sites, confirming the presence of anoxygenic phototrophic bacteria. As hypothesized, 16S rRNA gene ASV affiliations indicated that Rhodopila was one of the most abundant genera, and the most abundant bacterial phototroph, in both the Amphitheater and Hot Spring Basin spring communities (17% and 9%, respectively) (Fig. 2). The most abundant ASV at both sites exhibited 100% sequence similarity to the type strain, R. globiformis DSM 161^T^. Only the genus Thiomonas had a higher relative abundance at the Amphitheater site (Fig. 2), characterized as being facultatively chemolithoautotrophic with the ability to use sulfur compounds such as elemental sulfur, sulfide, or thiosulfate as electron donors (87, 88). Strains of Thiomonas have been implicated in the oxidation of arsenite in arsenic-rich acidic hot springs in YNP (89, 90), yet the total dissolved arsenic concentration at the Amphitheater site is several orders of magnitude lower (0.16 µM [E. L. Shock, unpublished data]). In the Hot Spring Basin spring, Acidithiobacillus, as well as Thiomonas, both possibly active in sulfur and/or sulfide oxidation, were more abundant than Rhodopila, whereas the most abundant taxon was the archaeal class Thermoplasmata, though the most abundant ASV exhibited poor sequence similarity (86%) with cultured representatives. Other notable taxa assigned to significant ASVs at the Hot Spring Basin site are the hydrogen oxidizer Hydrogenobaculum (99% sequence similarity [91]) and the archaeal ammonia oxidizer Nitrososphaera (83%–85% sequence similarity [92]). Most other abundant taxa in the springs are known to be involved in sulfur cycling, including Desulfobacteraceae, Desulfurella, Thermodesulfobium, and Desulforhabdus (Fig. 2).
Taxonomic affiliations of 16S (left) and 18S (right) rRNA gene sequences, with those for Rhodopila explicitly indicated. Sequence results correspond to DNA extracted from samples collected in 2017; 18S rRNA gene sequencing was completed for the Amphitheater site only.
16S rRNA gene sequences most closely affiliated with other bacterial phototrophs exhibited significantly lower relative abundances (<1%) than those affiliated with Rhodopila in the Amphitheater and Hot Spring Basin spring communities, including those affiliated with Rhodoblastus, Rhodovastum, and Rhodobacter, as well as the AAPB genera Acidiphilium and Acidisphaera. No sequences were most closely affiliated with Cyanobacteria. 18S rRNA gene sequences obtained from the Amphitheater spring include several taxa of algae as major constituents, specifically Bacillariophyta (diatoms), Chlorophyta (green algae), Chrysophyceae, and Synurophyceae (Fig. 2). In contrast, Rhodophyta comprised <1% of the 18S rRNA gene ASVs, and these were not most closely affiliated with Cyanidiophyceae, a class of unicellular rhodophytes that includes C. merolae, known to dominate phototrophic mats in acidic aquatic environments in YNP above 40°C, e.g., downstream portions of Lemonade Creek (8, 78, 79, 81). Fungi collectively comprised the largest non-phototrophic eukaryotic taxon (19% relative abundance), exceeded only by Bacillariophyta (23% relative abundance).
Pigment composition of the Amphitheater mat
R. globiformis synthesizes several unique methoxylated ketocarotenoids that have not been identified in other species (38–40). Analysis of pigment extracts from mat material collected at the Amphitheater site by LC-DAD-MS/MS confirmed the presence of the four ketocarotenoids (R.g. keto I–IV) previously identified under normal culture conditions (Fig. 3), supporting the sequencing results in demonstrating R. globiformis as a predominant phototroph in the mat. The online diode array spectra yielded absorption maxima in agreement with those previously reported (38), and the high-resolution molecular ions were consistent with the molecular formulas of the carotenoids. Collision-induced fragmentation of the molecular ions yielded fragments resulting from loss of methanol due to cleavage of one (M+H−32) or both (M+H−64) methoxy groups (or, in the case of IV, M+H−50, due to loss of a methoxy group and a hydroxyl group), consistent with the carotenoid structures.
(A) Diode array chromatogram at 475 nm of the pigment extract of Amphitheater sediments. The four ketocarotenoids diagnostic for Rhodopila globiformis are indicated with Roman numerals; other major peaks are labeled with their assignments as follows: f, fucoxanthin; bchl a, bacteriochlorophyll a; phede a, pheophorbide a; chl a, chlorophyll a; d, diatoxanthin; and β, β-carotene. (B) Diode array spectra of the four ketocarotenoids. (C) General structure of the ketocarotenoids (top) with the variable terminal R-groups colored as in (B) and high resolution mass to charge ratios for the molecular ions in the corresponding mass spectra averaged over the retention time of the diode array response (bottom). Further details are compiled in the supplemental material (Table S1).
A predominant chlorophyll in the pigment extract was bacteriochlorophyll a, which is the primary chlorophyll employed by R. globiformis (22, 37). Chlorophyll a was also detected, indicating the presence of oxygenic phototrophs and consistent with the presence of algae indicated by 18S rRNA gene sequencing. Degradation products of these chlorophylls, including bacteriopheophytin a, pheophytin a, pheophorbide a, and pyropheophorbide a, were also identified, all of which are common in moderately acidic springs (7). Aside from the ketocarotenoids associated with R. globiformis, other abundant carotenoids in the pigment extract were identified as diatoxanthin and fucoxanthin, carotenoids found in diatoms and other algae, in particular other Heterokont taxa, including Chrysophyceae and Synurophyceae (39, 93, 94), consistent with the 18S rRNA gene sequencing results. There was, however, no evidence for the presence of chlorophylls b or c, inconsistent with expectations given the algal taxa detected via 18S rRNA gene sequencing, except that pheophytin c1 and pheophytin c2 were identified, which possibly are the result of pheophytinization (i.e., loss of magnesium) of chlorophyll c. There were also no mass spectra consistent with Zn-bacteriochlorophyll a, which is synthesized by members of the genus Acidiphilium (95, 96) and has been detected in some other moderately acidic YNP springs (7). In spite of the enhanced resistance to pheophytinization of Zn-bacteriochlorophyll a (97), thought to be advantageous for acid tolerance by Acidiphilium, its sister genus Rhodopila apparently does not chelate bacteriochlorophyll a with zinc to a detectable extent, certainly not to the extent observed in Acidiphilium. A compilation of the results of the pigment analysis is located in the Supplemental material (Table S1).
Rates of inorganic and organic carbon assimilation
Microcosm carbon assimilation experiments were conducted at the Amphitheater site to assess the carbon metabolism of the microbial community. In 2017, a significantly higher rate of DIC assimilation was observed in microcosms exposed to light than in those excluded from light, indicating light-driven DIC assimilation was occurring in the mat at the time of the assay (Fig. 4). These microcosms were amended with DCMU to a final concentration of 10 µM, which is effective at decreasing light-driven assimilation of DIC to levels close to those observed in the dark in other acidic microcosms (Fig. S1), such as microcosms from the outflow of Dragon Spring where the predominant phototrophs are algae (10, 98). Thus, assimilation by algae in microcosms amended with DCMU is negligible, and the light-driven DIC assimilation rate observed at the Amphitheater spring may be attributed to anoxygenic phototrophs such as R. globiformis.
Rates of DIC assimilation (left) and acetate assimilation (right) in microcosms containing mat and spring water from the Amphitheater site in 2017. Microcosms were incubated in situ and were either exposed to light (white bars) or were excluded from light using aluminum foil (gray bars). Microcosms for assessing DIC assimilation were amended with 3-(3,4-dichlorophenyl)-1,1-dimethylurea to exclude oxygenic photosynthesis.
The light-driven DIC assimilation rate in 2017 was 7 ± 2 mg C g C^−1^ h^−1^, which is similar to, or in some cases greatly exceeds, rates of light-driven DIC assimilation measured in the absence of DCMU in some other acidic hot springs in YNP where algae (C. merolae) are the major phototrophs (Fig. 5) (12). As is the case for most purple nonsulfur bacteria (1), the genome of R. globiformis contains all genes necessary for carbon fixation via the reductive pentose phosphate cycle (i.e., Calvin cycle [52]). Such a high rate of light-driven DIC assimilation, rivaling or exceeding that of algae, suggests R. globiformis may be growing photoautotrophically under the conditions at the time of the assay. However, purple nonsulfur bacteria are known to employ the Calvin cycle as a means of dissipating excess reducing power to maintain redox balance during photoheterotrophic growth (99–103). For example, in wild-type Rhodopseudomonas palustris, it was estimated through ^13^C-metabolic flux analysis that 15.9% of biomass carbon was obtained via the Calvin cycle during photoheterotrophic growth on acetate (101). Anaplerotic reactions during growth on other organic substrates could also contribute to the observed assimilation of DIC. Thus, while the high rate of DIC assimilation may be sufficient to support the growth of R. globiformis in its natural habitat, it is nevertheless unclear if and to what extent DIC assimilation is paired with light-driven assimilation of organic compounds. Regardless, the observed light-driven assimilation of DIC represents a significant amount of primary production by anoxygenic phototrophs in the Amphitheater spring, which are almost exclusively comprised of R. globiformis.
Rates of light-driven DIC assimilation at the Amphitheater site in 2014 and 2017 (red crosses; Table S3) and rates at other hot spring locations in Yellowstone and Tengchong, China (black symbols) reported in the literature (12, 104–107) or included in this study (Table S4) versus the pH of the hot spring fluids at the sample location. Rates are calculated as the difference between rates measured using microcosms exposed to light and those determined using microcosms excluded from light (wrapped in aluminum foil). Rates for spring locations in China were calculated as the average rate over the afternoon (1–6 PM) photoperiod (107). The year that the rate was determined is indicated for the Amphitheater site. The rate at the outflow of Dragon Spring (Table S4) is not included, as only a minimum rate could be determined. For clarity, uncertainties in the rates are not depicted; rates that were not statistically distinguishable from zero are not included in the plot.
To examine the possibility of active photoheterotrophy, acetate was chosen as the organic substrate in light of its central role in carbon metabolism and the existence of measurable concentrations in many YNP hydrothermal fluids (74, 108). Rates of acetate assimilation in the light were over two orders of magnitude lower than rates of DIC assimilation in the light, and in contrast to DIC assimilation, rates of acetate assimilation were not different between microcosms incubated in the light and those excluded from light (Fig. 4), indicating that light-driven assimilation of acetate was not occurring in the spring when sampled. While it certainly remains possible that photoheterotrophy was occurring with other organic compounds, these results suggest that R. globiformis may execute photoautotrophy as its primary mode of carbon assimilation under typical conditions found at the Amphitheater spring. Nevertheless, it is plausible that rain events or other intermittent external processes may lead to spikes in DOC that would lead to an opportunistic temporary shift to photoheterotrophy, analogous to shifts between chemoautotrophy and chemoheterotrophy for which there is evidence in some hot springs in YNP (74, 109, 110). The DOC of acidic springs tends to be terrestrially derived (111, 112), consistent with intermittent surficial input of organic matter that could periodically drive photoheterotrophy. Both springs lie below marshy ground containing mosses and/or ferns and were also found to contain wood (Fig. 1), with the Amphitheater spring being bordered by a decaying log (presumably lodgepole pine, Pinus contorta). For example, this exogenous material could leach pentose sugars such as xylose into the spring waters via acid hydrolysis of pentosans such as xylan (8), consistent with R. globiformis possessing the genetic capacity for xylose metabolism (52).
A lack of light-driven assimilation of acetate may not be surprising given that growth of R. globiformis in culture was completely inhibited with acetate as the sole carbon source in early characterization studies by Pfennig (22). However, at the pH of the growth medium (5.6), a non-trivial portion (12.6%) of the acetate is speciated as acetic acid (Table S2). Growth medium was prepared by adding 1 g/L (12 mM) sodium acetate, resulting in a concentration of the neutral (protonated) acid of 1.5 mM at equilibrium. At such a concentration of the neutral species, the flux of acetic acid across the plasma membrane via passive diffusion may be detrimental to growth either by causing acidification of the cytoplasm upon deprotonation at the higher pH of the cytosol, forcing an unsustainable input of energy to maintain a suitable ΔpH across the plasma membrane (113), or due to toxicity of acetate itself (114, 115), thought to be caused by imbalances in essential metabolite concentrations created by efforts to counteract the increase in osmotic pressure (113). Regardless of mechanism, it may be that failure to observe growth on acetate was due to acetate toxicity rather than metabolic inability, the latter of which would be a notable shortcoming for a purple bacterium, as most purple bacteria can assimilate acetate (1, 116). All of the simple aliphatic carboxylates tested by Pfennig similarly inhibited growth, and only carboxylates where the calculated concentration of the neutral form was over an order of magnitude lower were found to support growth (Fig. S3). Thus, we hypothesized that in nature where total acetic acid + acetate concentrations are ~4 orders of magnitude less than those employed by Pfennig (0.83 µM in spring water; 1.4 µM in microcosms), despite being speciated more toward the protonated form (0.75 µM and 1.3 µM, respectively), R. globiformis may be capable of assimilating acetate into biomass by taking advantage of a manageable flux of acetic acid into the cytoplasm via passive diffusion. Indeed, there is evidence in the annotated genome (52) for the presence of acetate—CoA ligase, a key enzyme that would allow acetate to enter the tricarboxylic acid (TCA) cycle as acetyl-CoA, with carbon derived from acetate siphoned from the cycle as needed for anabolic processes. Replenishment of TCA cycle intermediates diverted to biosynthesis could be facilitated by the glyoxylate cycle (117), for which R. globiformis possesses the genetic capability (52). While it is possible that R. globiformis may lack one or more transport enzymes necessary for acetate metabolism, it is perhaps more plausible that for natural populations it is generally more efficient to take advantage of the significant DIC concentration of its environment (Table 1) and grow photoautotrophically, possibly via free membrane diffusion of aqueous carbon dioxide, the predominant form of DIC at low pH, rather than utilize much lower and perhaps dynamic concentrations of simple organic compounds for photoheterotrophic growth. It also remains possible R. globiformis may assimilate acetate independently of light, as chemotrophic growth in the dark at low O_2_ concentrations was reported with other organic substrates (22).
Acetate was dissimilated (oxidized to DIC) at a rate of ~15% of the rate of acetate assimilation, but only in microcosms excluded from light (Fig. S4), indicating that heterotrophs present in the Amphitheater mat only respired acetate in the dark. In comparison to rates of acetate metabolism at higher temperatures where phototrophs are excluded (74), the microbial community at the Amphitheater site exhibited a greater rate of assimilation relative to the rate of dissimilation than most hot springs examined, in particular when considering acidic springs (Fig. S5). On a dry mass basis, the acetate transformation rates are several orders of magnitude greater than those observed in high-temperature locations, which is undoubtedly due to the significantly larger amount of biomass per gram at the Amphitheater site than exists in the silica-rich sediments found in typical hot spring locations hosting chemosynthetic microbial communities.
Geochemical habitat
Physicochemical data were obtained to characterize the habitat of R. globiformis and the biomass present in the mat, which are compiled in Table 1. The Amphitheater spring fluids are acidic with moderate concentrations of sulfate and little dissolved chloride. These observations indicate the spring waters are derived from meteoric water with input of volcanic gases containing hydrogen sulfide, like other acidic springs in YNP, with little to no input of deeply sourced hydrothermal waters (49). The stable isotope ratios of H_2_O from the spring lie close to the local meteoric water line, only slightly shifted due to modest amounts of evaporation (Fig. S6). The pH values reported here are consistent with the limited previous observations of other YNP springs where R. globiformis has been observed (27). The pH of the Hot Spring Basin spring measured in the field may be erroneous, as suggested by charge balance calculations, presumably due to the probe being inadequately submerged in the extremely shallow (~1 cm) water and/or contacting spring sediments. The pH calculated on the basis of achieving charge balance suggests that the lower pH limit for R. globiformis is well below 3. In spite of a reported pH range for growth of 4.2–6.5 when grown photoheterotrophically (22, 27), clearly, R. globiformis is capable of at least photoautotrophic growth under more acidic conditions. The temperatures observed at the springs were below 40°C, the reported upper temperature limit, but in excess of the optimum temperature range observed for photoheterotrophic growth in culture (22, 27). 16S rRNA gene sequences affiliated with R. globiformis were previously obtained from locations in the Sylvan Springs area with much higher temperatures at the time of sampling (47.2°C and 49.1°C [12]), indicating that either the thermotolerance of at least some strains of R. globiformis is much greater than previously observed, or that the sequences were derived from inactive and/or exogenous cells, perhaps originating from acidic sinter soils surrounding the thermal waters.
The total dissolved sulfide concentration at the Amphitheater site in 2017 was 210 µM, the highest observed in this study (Table 1). Initial growth studies indicated that sulfide completely inhibited growth at low concentrations, yet the lowest concentration tested was~0.4 mM (0.01%, assumed to refer to Na_2_S∙9H_2_O [22, 48]), roughly twice the highest concentration observed in the springs. While it is possible sulfide concentrations are maintained at lower concentrations in the mat than in the overlying spring waters due to biological sulfide oxidation, it appears that R. globiformis can tolerate sulfide at least in the ~0.1 mM range. Indeed, a strain of Rhodopila isolated from Lassen Volcanic National Park was grown in a liquid culture medium containing 0.14 mM sulfide (45). It remains an open question, however, whether R. globiformis is capable of utilizing sulfide either as an assimilatory sulfur source or as an electron donor for photosynthesis. In culture, thiosulfate has typically been employed as a sulfur source but was found to largely be oxidized to tetrathionate, indicating R. globiformis is capable of dissimilatory thiosulfate oxidation, though it was questioned whether this was sufficient to support photoautotrophy (118). Under acidic conditions (pH < 6), thiosulfate disproportionates to elemental sulfur and sulfite (46), which would be expected to maintain low thiosulfate concentrations in nature that may be insufficient to serve as an assimilatory source of sulfur or as an electron donor. Alternatively, sulfate can serve as a sulfur source at concentrations <1 mM (48), but the concentrations of sulfate observed in springs hosting R. globiformis in this study are equal to or exceed 1 mM (Table 1), suggesting sulfate may also prove inadequate as an assimilatory sulfur source in nature. Though its genome appears to lack a complete sox pathway for sulfide oxidation (52), a preponderance of the above nevertheless indicates that the metabolic role of sulfide in R. globiformis merits attention. Alternatively, hydrogen could potentially serve as a reducing agent for photosynthesis, with all genes encoding for a Ni-Fe hydrogenase being present in the genome of the type species (52). Though not measured in this study, a moderate concentration of hydrogen was measured in gases emanating from a frying pan at Amphitheater springs (0.37 mol% [85, 86]), thus it is plausible that a metabolically relevant aqueous concentration of hydrogen exists at the Amphitheater spring hosting Rhodopila.
As noted previously, the springs feature plentiful DIC that R. globiformis appears to exploit for photoautotrophy. The carbon isotopic ratio (δ^13^C) of the Amphitheater biomass was −26‰ in both years of sampling (Table 1). Relative to the isotopic composition of the DIC (−4‰), these ratios represent a fractionation (Δ^13^C) of −22‰, which is consistent with biomass synthesis via the reductive pentose phosphate (Calvin) cycle (119, 120), being very close to the fractionation during photoautotrophic growth via the Calvin cycle in the purple sulfur bacterium Thermochromatium tepidum (−20.5‰ [121]). As the DIC is speciated in essence completely as aqueous carbon dioxide due to the acidic pH (99.7% of the DIC), it is possible that R. globiformis takes advantage of passive membrane diffusion of aqueous carbon dioxide, which is the DIC uptake mechanism of the acidophilic alga C. merolae (122). Isotopic ratios of mat nitrogen were slightly more variable and are more difficult to interpret without isotopic data of potential nitrogen substrates, though they are consistent with those of other acidic hot springs in Yellowstone (112, 123, 124), where fluids are enriched in isotopically light ammonium putatively derived from input of vapor-phase gases from the hydrothermal system (123, 124). Both springs have ammonium concentrations that are unlikely to be limiting for ammonium assimilation by R. globiformis (Table 1), its preferred nitrogen source in culture (125). Though it has the genetic capacity for both ammonium assimilation and N_2_ fixation (52), diazotrophy is repressed in the presence of non-limiting ammonium concentrations (125).
The springs in this study are geochemically similar to moderately acidic hot springs that host Cyanobacteria and algae (7), but are distinct in their lower temperatures and moderate concentrations of sulfide (Table 1; Fig. 6). In both cases, the moderately acidic pH values and moderate temperatures suggest a larger ratio of meteoric water to volcanic gas (vapor phase of the hydrothermal system) when compared to other acidic hot springs in YNP (7). Here, the persistence of sulfide in the spring water suggests that the injection of volcanic gases into oxygenated groundwater occurs very close to the surface, leading to the incomplete oxidation of sulfide. The modest temperatures of the springs additionally might indicate that the hydrothermal gases have cooled somewhat prior to mixing with groundwater. Both sites occur on the edges of thermal areas and beneath abundant terrestrial vegetation, indicative of cold nearby ground temperatures. The acidic spring waters do show evidence of some water-rock/soil reaction, given the major cation concentrations (Table 1). At the Amphitheater site, the major ion data indicate the spring water was more dilute in 2017 than in 2014, perhaps indicating a lower fluid flux or deeper input of gases in 2014 that would allow more time for sulfide oxidation and water-rock reactions, resulting in a lower sulfide concentration, lower pH, higher specific conductance, and slightly higher major ion concentrations.
The logarithm of the total dissolved sulfide concentration versus temperature (left) and pH (right) of selected moderately acidic to circumneutral springs in Yellowstone where the predominant phototrophs are from the genus Rhodopila (●; this study), the genus Thermochromatium (■; 121, 126), the phylum Cyanobacteria (+; 7), or the genus Chlorobaculum (▲, 127). Locations with Chlorobaculum mats in New Zealand (♦, 128) and the Philippines (▬, 127) are also included. In cases where ranges are reported for sites with Chlorobaculum, the highest sulfide concentration, the highest temperature, and the lowest pH are plotted.
The aforementioned purple sulfur bacterium T. tepidum (originally Chromatium tepidum [129]) has been studied in (and originally isolated from) springs in the Mammoth area in northern YNP with sulfide concentrations similar to those observed here (Fig. 6) (121, 126). Relative to R. globiformis, this organism is characterized by greater thermotolerance, being found in springs with temperatures of 44°C–55°C (121, 126, 130) and exhibiting a maximum temperature of 57°C in culture (130). These springs are also of higher pH, with values of 6.2–6.7 (121, 126), and the optimum pH of T. tepidum in culture is 7, though the pH range for growth has not been reported (130). The pH of spring waters at Mammoth is controlled by subsurface water-rock reactions with limestone, leading to fluids supersaturated with carbon dioxide that precipitate carbonates (travertine) upon degassing of carbon dioxide at the surface (131). Thus, these spring waters are buffered against significant decreases in pH resulting from the oxidation of sulfide, but depending on the extent of acid tolerance of T. tepidum, it is possible that springs with pH values <6 similar to those hosting R. globiformis but of somewhat higher temperature that could be habitable to T. tepidum may exist elsewhere in YNP. Thus, temperature and perhaps pH would dictate whether a moderately acidic spring with moderate levels of sulfide is more favorable to R. globiformis or T. tepidum. The green sulfur bacterium Chlorobaculum tepidum (previously Chlorobium tepidum; 132) inhabits waters with concentrations of dissolved sulfide of 0.3–1.8 mM in hot springs where it has been studied in New Zealand (Fig. 6) (128), though Chlorobaculum spp. in YNP were found in springs with low sulfide concentrations, similar to sulfide concentrations in springs hosting Cyanobacteria (Fig. 6) (127). However, YNP strains were successfully enriched in media containing 1 mM sulfide, indicating that if springs with high (~mM) concentrations of sulfide are identified in YNP, such conditions may exceed the tolerances of either the aforementioned purple bacteria (R. globiformis and T. tepidum) and could select for Chlorobaculum.
Though we report on the observation of springs featuring Rhodopila as the predominant anoxygenic phototroph in two additional thermal areas in YNP, springs with physicochemical conditions like those characterized here sustaining populations of R. globiformis nevertheless still appear to be relatively rare, such that it is essentially an endangered species (27). Important physicochemical components of the niche space for R. globiformis seem to include modest temperatures (30°C–40°C), acidic pH (<4), and moderate dissolved sulfide (~0.1 mM). Acidic springs may be found in YNP from ambient temperatures up to boiling (133), but the hydrological conditions allowing for a relatively small flux of hydrothermal gases into groundwater very close to the surface, resulting in moderate sulfide concentrations in springs only slightly elevated above ambient temperature, are apparently comparatively uncommon. Claims of low concentrations of sulfide being inhibitory for the growth of R. globiformis notwithstanding, moderate sulfide concentrations instead appear to be essential for the establishment of abundant natural populations. A continuous flux of hydrogen sulfide consumes dissolved oxygen in oxidation reactions and thereby maintains the low oxygen concentrations necessary for anaerobic or microaerobic phototrophic growth that otherwise may be exceeded due to the influx of atmospheric oxygen. Similarly, sulfide oxidation may quench oxygen produced in the spring by algae, the presence of which is indicated by 18S rRNA gene sequences (Fig. 2) and detection of chlorophyll a and the carotenoids diatoxanthin and fucoxanthin (Fig. 3), though it is unclear if and to what extent these algae are active in the spring and whether they are autochthonous. Additionally, the observed sulfide concentrations likely exceed the tolerances of at least some acidophilic algae, such as C. merolae, due to sulfide inhibition of electron transfer associated with photosystem II (10, 134), thereby freeing R. globiformis of some degree of interspecific competition for photo(auto)trophic niche space.
Conclusions
Since its isolation roughly 50 years ago, R. globiformis has undergone numerous culture-dependent investigations revealing interesting features of its biochemistry, but knowledge of its ecology has not similarly expanded over that time, previously limited to brief reports of the isolation of similar organisms from new locations. In this work, we more thoroughly characterized the physicochemical conditions of its natural habitat and conducted carbon uptake experiments in the field to observe aspects of its physiology in situ. As purple nonsulfur bacteria are thought to favor and excel at photoheterotrophic growth, they were originally isolated and have been exclusively grown in the lab with organic substrates. The rate measurements here demonstrate that in its natural environment, it is responsible for a significant amount of primary production, utilizing the abundant supply of aqueous carbon dioxide offered by the flux of volcanic gases (i.e., magmatic CO_2_) into the spring waters, though to what extent this is combined with assimilation of organic compounds remains unclear. To our knowledge, the pH of the Amphitheater spring is the lowest at which light-driven DIC assimilation has been observed in a member of the domain Bacteria.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Madigan MT, Jung DO. 2009. An overview of purple bacteria: systematics, physiology, and habitats. Adv Photosynth Respir 28:1–15.
- 2Madigan MT. 2003. Anoxygenic phototrophic bacteria from extreme environments. Photosynth Res 76:157–171. doi:10.1023/A:102499821268416228575 · doi ↗ · pubmed ↗
- 3Imhoff JF. 2017. Anoxygenic phototrophic bacteria from extreme environments, p 427–480. In Hallenbeck PC (ed), Modern topics in the phototrophic prokaryotes: environmental and applied aspects. Springer International Publishing.
- 4Colman DR, Poudel S, Hamilton TL, Havig JR, Selensky MJ, Shock EL, Boyd ES. 2018. Geobiological feedbacks and the evolution of thermoacidophiles. ISME J 12:225–236. doi:10.1038/ismej.2017.16229028004 PMC 5739016 · doi ↗ · pubmed ↗
- 5Colman DR, Keller LM, Arteaga-Pozo E, Andrade-Barahona E, St Clair B, Shoemaker A, Cox A, Boyd ES. 2024. Covariation of hot spring geochemistry with microbial genomic diversity, function, and evolution. Nat Commun 15:7506. doi:10.1038/s 41467-024-51841-539209850 PMC 11362583 · doi ↗ · pubmed ↗
- 6Brock TD. 1973. Lower p H limit for the existence of blue-green algae: evolutionary and ecological implications. Science 179:480–483. doi:10.1126/science.179.4072.4804196167 · doi ↗ · pubmed ↗
- 7Fecteau KM, Boyd ES, Lindsay MR, Amenabar MJ, Robinson KJ, Debes RV II, Shock EL. 2022. Cyanobacteria and algae meet at the limits of their habitat ranges in moderately acidic hot springs. JGR Biogeosciences 127:e 2021 JG 006446. doi:10.1029/2021 JG 006446 · doi ↗
- 8Brock TD. 1978. Thermophilic microorganisms and life at high temperatures. Springer-Verlag, New York.