Pleiotropic role of PAX cyclolipopeptides in the Xenorhabdus bacterium mutualistically associated with entomopathogenic nematodes
Noémie Claveyroles, Anne Lanois-Nouri, Imane El Fannassi, Jean-Claude Ogier, Sylvie Pagès, Adrien Chouchou, Guillaume Cazals, Gilles Valette, Alyssa Carré-Mlouka, Alain Givaudan

TL;DR
This study explores the ecological roles of PAX cyclolipopeptides in Xenorhabdus bacteria, revealing their impact on biofilm formation, motility, and nematode interactions.
Contribution
The study identifies multiple new ecological roles for PAX peptides in Xenorhabdus-nematode mutualism.
Findings
PAX peptides are present throughout the Xenorhabdus nematophila life cycle.
PAX peptides promote biofilm formation and reduce swimming motility.
PAX peptides enhance the production of infective Steinernema juveniles.
Abstract
Xenorhabdus is an entomopathogenic bacterium involved in a mutualistic relationship with Steinernema nematodes. Xenorhabdus produces a multitude of specialized metabolites by non-ribosomal peptide synthetase (NRPS) pathways to mediate bacterium-nematode-insect interactions. PAX cyclolipopeptides are a family of NRP-type molecules whose ecological role remains poorly understood. In this study, the pleiotropic role of PAX peptides in the life cycle of Xenorhabdus nematophila has been investigated. By mass spectrometry analysis, we first demonstrated that PAX peptides were detected from the pathogenic stage up to the necrotrophic stage. We discovered that the bromothymol blue adsorption phenotype historically used to discriminate Xenorhabdus variants was associated with the presence of PAX peptides. We found that PAX peptides were positively involved in biofilm formation and negatively…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
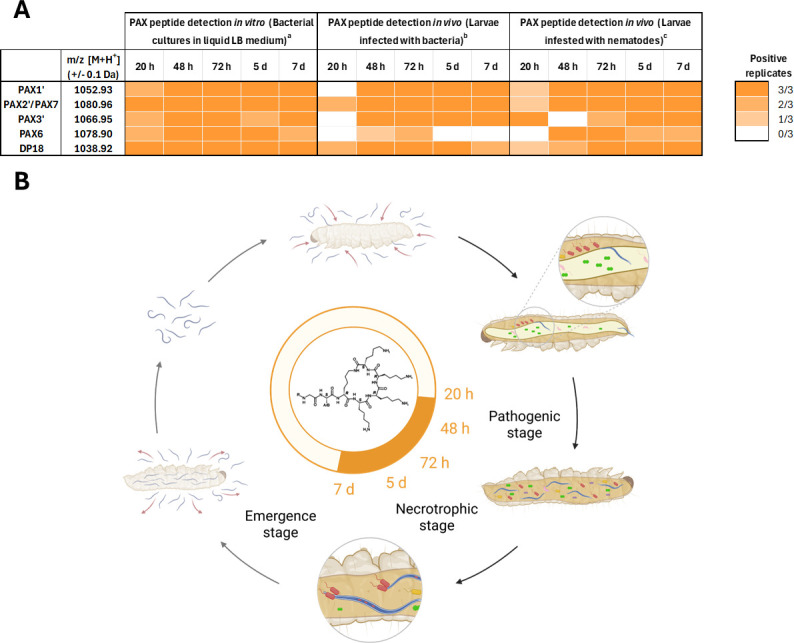 Fig 1
Fig 1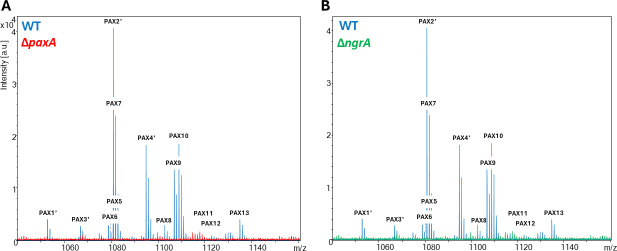 Fig 2
Fig 2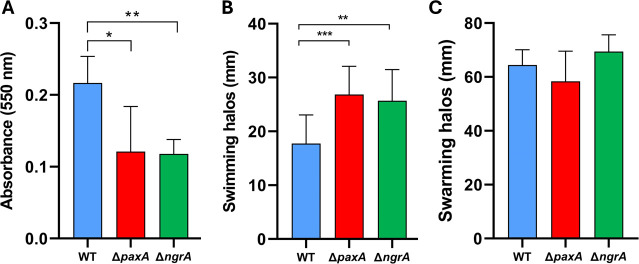 Fig 3
Fig 3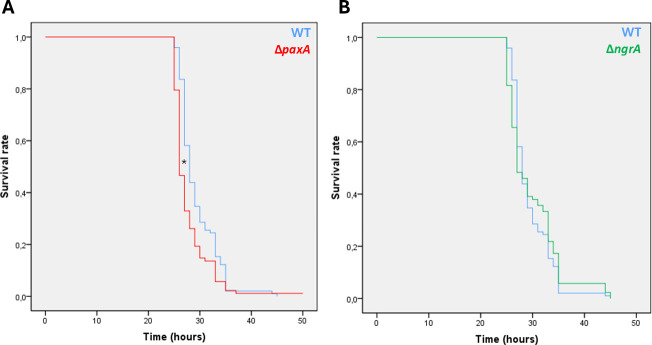 Fig 4
Fig 4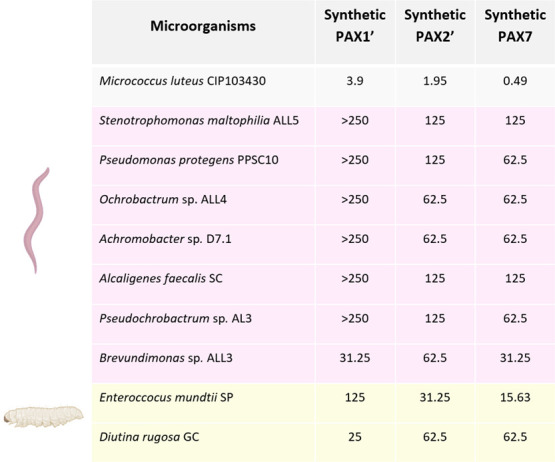 Fig 5
Fig 5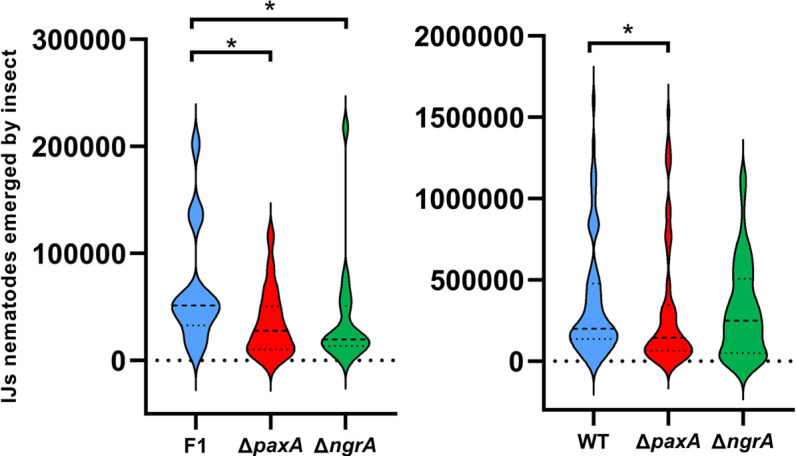 Fig 6
Fig 6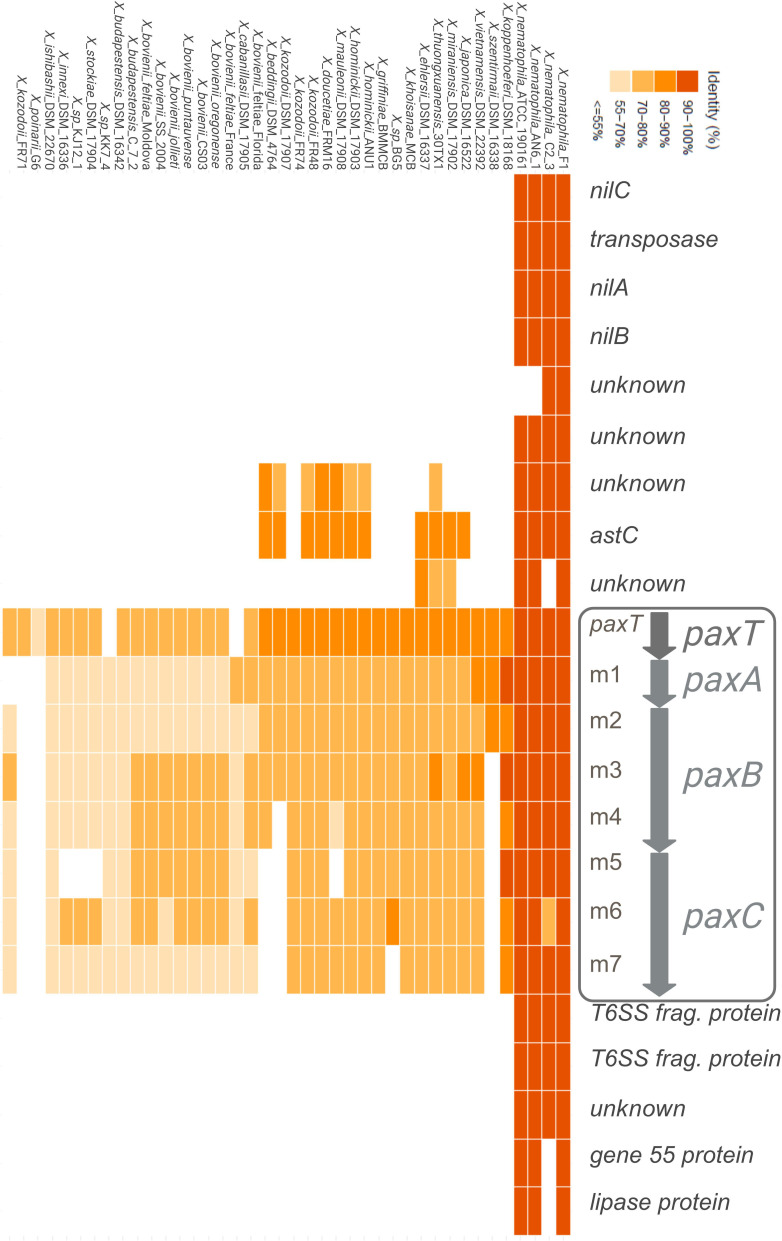 Fig 7
Fig 7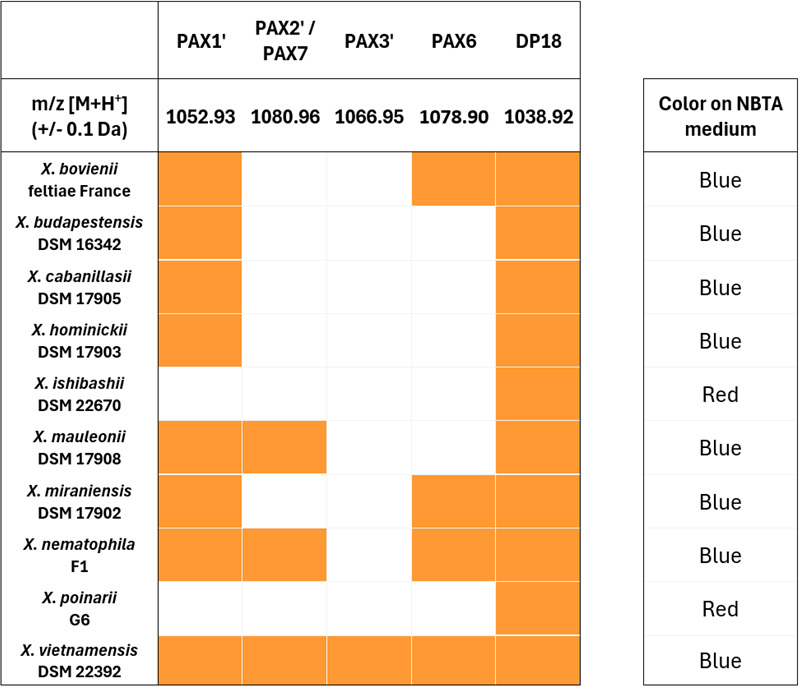 Fig 8
Fig 8| Phenotypic assay | WT | ∆ | ∆ |
|---|---|---|---|
| Lipolysis | |||
| Tween 20, Tween 40, Tween 60, Tween 80 | W+ | + | + |
| Tween 85 | W+ | + | + |
| Lecithinase-like activity | + | − | − |
| Antimicrobial activity ( | 37.5 | 37.6 | 15.7 |
| Hemolysis | T | T | T |
| Adsorption of bromothymol blue and TTC degradation into formazan | Blue | Red | Red |
| Antimicrobial peptide | WT | ∆ | ∆ |
|---|---|---|---|
| Cecropin A | 12.5 | 12.5 | 10.5 |
| Cecropin B1 | 6.25 | 6.25 | 6.25 |
| Cecropin C | 12.5 | 12.5 | 25 |
| Cecropin P1 | >12.5 | >12.5 | 12.5 |
| Drosocin | >50 | >50 | >50 |
| Colistin | 3.9 | 1.95 | 3.9 |
| Polymyxin B | 0.245 | 0.245 | 0.245 |
| NOSO 95C (odilorhabdin) | >1,700 | >1,700 | >1,700 |
| Synthetic PAX1′ | >1,500 | >1,500 | >1,500 |
| Synthetic PAX2′ | 187.5 | 375 | 375 |
| Synthetic PAX7 | 187.5 | 187.5 | 375 |
- —Institut National de la Recherche Agronomiquehttp://dx.doi.org/10.13039/501100006488
- —Université de Montpellierhttp://dx.doi.org/10.13039/501100008222
- —Université de Montpellierhttp://dx.doi.org/10.13039/501100008222
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEntomopathogenic Microorganisms in Pest Control · Insect symbiosis and bacterial influences · Nematode management and characterization studies
INTRODUCTION
Entomopathogenic bacteria of the genus Xenorhabdus (Morganellaceae) are hosted by nematodes of the Steinernema genus, with which they have co-evolved to establish long-lasting, mutually beneficial interactions (1). Xenorhabdus are carried into the insect larvae by the infective juvenile (IJ) stage of nematodes. IJs live in the soil, seeking insect larvae to infest. Once they have found a prey, IJs penetrate insect larvae through their natural orifices and release their Xenorhabdus bacteria into the insect hemocoel (2). Bacteria cause septicemia and death of the insect larvae within 24–48 h, enabling the nematodes to use the insect cadaver as a nutritive resource and a host for their development (3). Several nematode reproduction cycles occur successively inside the cadaver (4). During this necrotrophic stage, Xenorhabdus and Steinernema nematodes interact with a range of microorganisms from the nematode microbiota (frequently associated microbiota, FAM) and the insect microbiota (5–7). When nutrients are depleted, mutualistic bacteria and nematodes reassociate, and IJs emerge in the soil seeking a new host. Entomopathogenic nematodes are considered promising biological control agents of key insect pests (8).
To cope with changing conditions during its complex life cycle, Xenorhabdus can produce a wide variety of specialized metabolites, including non-ribosomal peptides (NRPs) (9, 10). Non-ribosomal peptide synthetases (NRPSs) are modular enzymes that catalyze the synthesis of a wide range of peptides from a variety of proteinogenic and non-proteinogenic amino acid substrates. In Xenorhabdus nematophila, the ngrA gene encodes the phosphopantetheinyl transferase (PPTase), an enzyme that activates NRPS by transferring a cofactor (4′-phosphopantetheine), thus enabling the production of NRP-type metabolites (11). It has been shown that ngrA-dependent compounds are responsible for most of the antimicrobial activity of X. nematophila and are required to eliminate bacterial competitors in vitro (12). It has also been demonstrated that ngrA-dependent compounds from X. nematophila are required for optimal growth and development of their nematode partner in vivo. Indeed, Steinernema carpocapsae reproduction was reduced in insects infected with an ngrA mutant (12). Similar results have been observed with Photorhabdus luminescens associated with the nematode Heterorhabditis bacteriophora (13).
Studies on NRP metabolites in Xenorhabdus have so far focused mainly on their antimicrobial potential (14). These include xenocoumacins, odilorhabdins, xenoamicins, xenematides, xenortides, rhabdopeptides, RXPs, GameXpeptides, nematophin, xenobactin, szentiamide, bicornitun, taxlllaids A-G, and PAX peptides (15–28). Some of these compounds also exhibit insecticidal (xenocoumacins [29], xenematides [18], RXPs, and [21]), antiprotozoal (xenoamicins [17], xenortides [19], RXPs [21], xenobactin [24], and taxlllaids A-G [27]), acaricide (xenocoumacins [30]), and anti-inflammatory (xenocoumacins [31]) properties.
Our study focuses on the cyclolipopeptides PAX (peptide antimicrobial from Xenorhabdus) family (28). NRPS enzymes involved in PAX peptide biosynthesis are encoded by the paxTABC gene cluster. The paxA, paxB, and paxC genes encode the three NRPS enzymes responsible for heptapeptide synthesis and cyclization. The paxT gene encodes a putative membrane transport protein suspected of exporting PAX peptides outside the cell. PAX cyclolipopeptides are cationic peptides containing seven amino acids, among which five lysines constitute a cycle, with an N-terminal fatty acid moiety (32). Different PAX peptides, with variations in the length of the fatty acid chain or the nature of the amino acid in position 2 (lysine or arginine), have been identified among different strains and species of Xenorhabdus (28, 32, 33). Bacterial specialized metabolites are widely investigated in the literature for their medical or agricultural applications, but their ecological roles are still poorly understood (34). PAX peptides were first studied for their antimicrobial activities against Micrococcus luteus and Fusarium oxysporum (28). PAX peptides exhibit no insecticidal activity against Spodoptera littoralis, Galleria mellonella, Locusta migratoria, Manduca sexta, and insect hemocytes, nor nematicidal activity against Caenorhabditis elegans (28, 35). As external addition of synthetic labeled PAX peptides to Xenorhabdus doucetiae resulted in their localization at the cell surface, it was suggested that PAX peptides may provide protection against cationic antimicrobial peptides (AMPs) produced by the insect via positive charge repulsion mechanisms (36).
In this study, we revealed by mass spectrometry that PAX peptides were detected from the pathogenic stage to the late necrotrophic stage of the X. nematophila life cycle. We therefore aimed to elucidate the role(s) of PAX peptides using two deletion mutants of X. nematophila F1, ∆paxA and ∆ngrA mutants (37). By comparing the two mutants with the wild-type (WT) strain, we demonstrated different phenotypic characteristics on agar media, in swimming motility, and in biofilm formation in vitro. By minimum inhibitory concentration (MIC) assays, we showed that PAXs were weakly antimicrobial toward natural competitors of Xenorhabdus. We also assessed the pathogenicity of bacteria against insect larvae. Using aposymbiotic nematodes reassociated with the WT strain or the ∆paxA or ∆ngrA mutants, we investigated the involvement of PAX peptides in nematode reproductive success in vivo. Finally, we have highlighted the distribution of paxTABC genes and detected PAX peptide production throughout the Xenorhabdus genus. These findings reveal that PAX peptides are highly conserved compounds playing multiple roles in the Xenorhabdus life cycle.
RESULTS
PAX peptides are mostly detected during the necrotrophic stage of the X. nematophila life cycle
To understand the importance of PAX peptides in the life cycle of X. nematophila F1, PAX peptide presence was investigated at each stage of the bacterial life cycle. We first characterized all PAX peptides produced in stationary-phase extracts of the WT strain in a 48-h liquid culture using MALDI-TOF-MS. Twelve PAX peptides previously identified in the literature by Fuchs et al. (32) were detected in stationary-phase extracts of the WT strain: PAX1′ (m/z = 1,052.772 Da), PAX3′ (1,066.787), PAX2′/PAX7 (1,080.783), PAX4′ (1,094.797), PAX6 (1,078.766), PAX8 (1,102.772), PAX9 (1,106.797), PAX10 (1,108.812), PAX11 (1,120.808), PAX12 (1,122.818), and PAX13 (1,134.828) (Table S1) (32).
Then, the presence of most frequently detected PAX peptides (PAX1′, PAX2′/PAX7, PAX3′, and PAX6) was analyzed at 20 h, 48 h, 72 h, 5 days, and 7 days under three different experimental setups: (i) in vitro bacterial cultures, (ii) insect larvae infected with X. nematophila WT bacteria, (iii) insect larvae infested with S. carpocapsae SK27 nematodes carrying X. nematophila WT bacteria (Fig. 1; Table S2). An extraction control was performed by adding a synthetic PAX derivative (DP18, Fig. S1) before the extraction steps. The m/z values (±0.1 Da) of selected PAX peptides and the extraction control DP18 were screened by MALDI-TOF-MS analysis. In bacterial culture in lysogeny broth (LB) culture medium, PAX peptides were consistently found from 20 h (late exponential stage, Fig. S2) up to 7 days. Under in vivo conditions of Galleria mellonella larvae infected with bacteria or infested with nematodes, higher variability was observed at 20 h between biological samples. However, PAX peptides were detected from 48 h to 7 days in both in vivo conditions, except for PAX6, which is less frequently found in bacterial-infected larvae. These results demonstrate that PAX peptides are detected from the end of the pathogenic phase (20 h) to the late necrotrophic phase (7 days) of the Xenorhabdus life cycle.
Presence of PAX peptides in extracts of X. nematophila WT grown in vitro and in vivo. (A) Detection by MALDI-TOF-MS analysis of selected PAX peptides (PAX1′, PAX2′/PAX7, PAX3′, and PAX6) and the extraction control DP18 in extracts of X. nematophila WT grown in vitro in liquid LB medium (a) and in vivo in 10 G. mellonella larvae injected with X. nematophila WT (b) or infested with Steinernema carpocapsae SK27 IJ nematodes (c). A synthetic PAX peptide derivative DP18 (m/z = 1,038.92 ± 0.1 Da) was added as a positive extraction control before the extraction steps. Experiments were performed in triplicate. Orange squares indicate the number of positive biological replicates in which the m/z value (±0.1 Da) of the corresponding PAX peptides was detected. The corresponding mass spectra are given in Table S2. (B) Visual representation of PAX peptide presence in different stages of the Xenorhabdus life cycle. The orange circle area represents the period of detection of PAX peptides. Figure was created with Biorender.
paxA mutation affects lecithinase-like activity, lipolytic activity, and bromothymol blue adsorption
To assess the impact of PAX peptide production on phenotypic characteristics, a deletion mutant (∆paxA) was constructed in X. nematophila F1 by inserting an Ω-cam interposon by allelic exchange at the beginning of the paxA gene, the first NRPS gene in the paxTABC cluster. As phosphopantetheinyl transferase enzymes enable NRPS mega-synthetase activation, a PPTase-deficient mutant (∆ngrA) of X. nematophila F1 was also used as a global non-producer of NRP-type metabolites. The absence of PAX peptide production by ∆paxA (Fig. 2A) and ∆ngrA (Fig. 2B) mutants was confirmed by MALDI-TOF-MS in extracts from stationary-phase cultures.
Presence of PAX peptides in extracts of X. nematophila WT (blue), ∆paxA (red), and ∆ngrA (green). (A) Merging of MALDI-TOF-MS mass spectra of extracts from 48 h stationary-phase cultures of WT and ∆paxA strains. (B) Merging of MALDI-TOF mass spectra of extracts from 48 h stationary-phase cultures of WT and ∆ngrA strains. The monoisotopic m/z values of PAX peptides are shown with their numbers previously identified by Fusch et al. (32). Peak heights represent intensity in arbitrary units (a.u). Exact m/z values (±0.05 Da) of each PAX peptide are listed in Table S1.
The phenotypic traits of the ∆paxA and ∆ngrA mutants were then tested through various phenotypic assays on agar media. The ∆paxA and ∆ngrA mutants displayed enhanced lipolytic activity on Tween 20, 40, 60, 80, and 85 and suppressed lecithinase-like activity compared to the WT strain (Table 1). However, no difference in hemolysis was observed. As expected, the ∆ngrA mutant showed weaker antimicrobial activity against M. luteus than the WT strain or ∆paxA mutant. Surprisingly, both mutant strains displayed red colonies on Nutrient Bromothymol Blue Agar (NBTA) medium, indicating no bromothymol blue (BBT) adsorption (Fig. S3). These observations demonstrate that PAX peptides are responsible for BBT adsorption. Together, these results indicate that the mutation of ΔpaxA and ΔngrA induces pleiotropic effects on phenotypes of X. nematophila that are similar for both mutants, except for differences in antimicrobial activity against M. luteus.
PAX peptides are involved in biofilm formation and swimming motility
Biofilm formation, swimming motility, and swarming motility were investigated in vitro for X. nematophila WT, ΔpaxA, and ΔngrA strains (Fig. 3). The ΔpaxA (P < 0.05) and ΔngrA (P < 0.01) mutants showed significantly lower biofilm formation than the WT strain (Fig. 3A). Conversely, ΔpaxA (P < 0.001) and ΔngrA (P < 0.01) mutants showed significantly enhanced swimming motility on 0.35% LB agar medium compared with the WT strain (Fig. 3B). No difference in swarming motility was observed between the three strains on a 0.7% Eiken agar plate (Fig. 3C). PAX peptides are therefore positively involved in biofilm formation and negatively in swimming motility.
*Biofilm formation, swimming motility, and swarming motility of X. nematophila WT (blue), ∆paxA (red), and ∆ngrA (green). (A) Biofilm formation was assessed by 1% crystal violet staining of 48 h static bacterial cultures and quantification of absorbance at 550 nm. Student’s t test, n = 4. Swimming motility (B) and swarming motility (C) were assessed after dropping 5 µL of overnight bacterial culture onto 0.35% LB agar (Student’s t test, n = 4) and 0.7% LB agar Eiken (Wilcoxon-Mann-Whitney test, n = 4) media, respectively. Halo diameter was measured after 18 h of growth. *P < 0.05, **P < 0.01, and **P < 0.001.
PAX peptides are slightly involved in bacterial virulence toward insects
To determine whether PAX peptides were involved in the virulence of X. nematophila in vivo, pathogenicity assays of X. nematophila WT, ΔpaxA, and ΔngrA strains were carried out by injection into S. littoralis larvae (Fig. 4). A weak difference (1h40, P < 0.05) of larval mortality between the ΔpaxA mutant and the WT strain was observed (Fig. 4A), but no difference between the ΔngrA mutant and the WT strain (Fig. 4B). In a control experiment, we showed that the WT and mutant strains displayed no growth difference when cultivated in LB (Fig. S2).
*Pathogenicity assay of X. nematophila WT (blue), ∆paxA (A, red), and ∆ngrA (B, green) toward S. littoralis larvae (L6 stage). Kaplan-Meier survival curves were generated with SPSS version 18.0. Each insect was injected with 1,000–3,000 CFUs of the respective strains. Wilcoxon-Mann-Whitney tests, n = 80, P < 0.05.
To test the hypothesis of a role for PAX peptides in the resistance of X. nematophila to insect AMPs (36), MIC assays were carried out using various insect AMPs (cecropin A, cecropin B1, cecropin C, cecropin P1, and drosocin) against the WT, ΔpaxA, and ΔngrA strains. MIC assays were also carried out using bacterial AMPs (colistin, polymyxin B, NOSO 95C, synthetic PAX1′, synthetic PAX2′, and synthetic PAX7) to determine whether PAX peptides would also enable resistance to competitive bacteria within the insect cadaver (Table 2; Fig. S1). No clear difference in MIC was detected between the WT and mutant strains for any of the bacterial or insect AMPs tested. In our experiments, PAX peptides were not involved in the resistance of X. nematophila to the tested insect and bacterial AMPs.
Microorganisms inhabiting the ecological niche of Xenorhabdus exhibit low susceptibility to PAX peptides
To determine whether the production of PAX peptides could enable Xenorhabdus to outcompete the microorganisms present in the insect cadaver, MIC assays were performed using synthetic PAX1′, PAX2′, and PAX7 (Fig. 5; Fig. S1). For this purpose, 10 microorganisms from the microbiota (FAM) of S. carpocapsae nematodes (Stenotrophomonas maltophilia ALL5, Pseudomonas protegens PPSC10, Ochrobactrum sp. ALL4, Achromobacter sp. D7.1, Alcaligenes faecalis SC, Pseudochrobactrum sp. AL3, and Brevundimonas sp. ALL3) and from the microbiota of the insect Spodoptera (Enterococcus mundtii SP and Diutina rugosa GC) were selected. M. luteus CIP103430 was also used as a positive control for the antimicrobial activity of synthetic PAX peptides, as it had already been identified as susceptible to PAX peptides (28). Microorganisms from the nematode microbiota appear to exhibit low susceptibility to synthetic PAX tested with a minimum MIC of 31.25 µg mL^−1^ for PAX1′ and PAX7 against Brevundimonas sp. E. mundtii and D. rugosa (insect microbiota) show higher susceptibilities, with MICs down to 15.63 µg mL^−1^ for PAX7 against E. mundtii and 25 µg mL^−1^ for PAX1′ against D. rugosa. The antimicrobial effect of synthetic PAX peptides against the microorganisms co-occurring with Xenorhabdus in the insect cadaver niche is moderate.
Minimum inhibitory concentration (µg mL−1) assays of synthetic PAX1′, PAX2′, and PAX7 on selected microorganisms from nematode microbiota (Steinernema carpocapsae, pink) and insect microbiota (Spodoptera larvae, yellow).
PAX peptides enhance infective juvenile nematode production
Since PAX peptides were detected during the necrotrophic stage (Fig. 1), we questioned whether PAX peptides are involved in the production of IJ nematodes. G. mellonella larvae were infested with S. carpocapsae SK27 reassociated with the WT strain, ΔpaxA, or ΔngrA mutants (Fig. 6). A difference in reproductive success was observed between the WT strain and both mutant strains (n = 14–19) at the end of the first generation (Fig. 6, left), then between the WT strain and the ΔpaxA mutant (n = 60) at the end of the second generation (Fig. 6, right, Wilcoxon-Mann-Whitney tests, P < 0.05). Additionally, no significant difference in the timing of IJ emergence was noticed in both mutants (Fig. S4). The second generation selects the best-performing IJs capable of completing a full life cycle, thereby reducing the differences observed between strains. Furthermore, no significant differences in CFU per IJs were detected at the second generation between the WT strain and both mutant strains (Fig. S6). PAX peptides are therefore involved in the production of S. carpocapsae IJs in vivo.
*Reproductive success of aposymbiotic Steinernema carpocapsae SK27 nematodes reassociated with X. nematophila WT, ∆paxA, or ∆ngrA in G. mellonella larvae. Emerging IJ nematodes were counted at 15 and 30 days post-mortem and cumulated. The total number of IJs emerging per insect is represented at the end of the first generation (n = 14–19) (left) and second generation (n = 60) on three independent experiments (right). Wilcoxon-Mann-Whitney test, P < 0.05.
PAX peptide-producing ability is widespread within the Xenorhabdus genus
To determine whether PAX peptide production ability is conserved within the Xenorhabdus genus, we first studied the distribution and the genetic environment of the paxTABC genes in 40 strains of Xenorhabdus belonging to 23 species compared to X. nematophila F1 (Fig. 7). The surrounding genes and their organization are conserved among X. nematophila species, especially the nilABC genes, but not in other Xenorhabdus species. For NRPS genes, modules (paxA [m1], paxB [m2, m3, and m4], and paxC [m5, m6, and m7]) were analyzed independently as functional entities. paxTABC genes are present in almost all studied Xenorhabdus strains, except Xenorhabdus poinarii G6 and Xenorhabdus ishibashii DSM 22670. Although the entire cluster is well distributed among the Xenorhabdus strains, the paxC gene (m5, m6, and m7) is absent in a few genomes (Xenorhabdus szentirmaii DSM 16338, Xenorhabdus kozodoii DSM 17907, and Xenorhabdus beddingii DSM 4764). Pairwise alignments showed that the paxTABC cluster exhibits high conservation across all strains within Xenorhabdus bovienii species and within X. nematophila species but shows lower identity between the two species (Fig. S5). Thus, the paxTABC cluster is present throughout the entire Xenorhabdus genus, but with variability among NRPS modules.
Distribution and genetic environment of paxTABC among the Xenorhabdus genus. The paxTABC and surrounding genes of X. nematophila F1 were aligned with those of 40 strains belonging to 23 Xenorhabdus species by NCBI BLAST+blastn. The NRPS genes within this cluster were specifically analyzed by comparing their module nucleotide sequences (paxA: m1; paxB: m2, m3, and m4; paxC: m5, m6, and m7). Orange gradient squares represent the nucleotide percentage identity of genes and modules for each orthologous sequence compared to their counterpart of X. nematophila F1. The paxTABC operon is shown at the top of the figure, along with its surrounding genes in X. nematophila F1 genome.
To correlate the presence of the paxABC genes with the actual production of PAX peptides, mass spectrometry analysis (MALDI-TOF-MS) was then performed on extracts from 48-h cultures of nine different strains representative of the main species of Xenorhabdus (Fig. 8; Table S3). The analysis focused on the m/z values (±0.1 Da) of the most detectable PAX peptides, PAX1′, PAX2′/PAX7, PAX3′, PAX6, and DP18 as the positive extraction control. The DP18 control was detected in all samples. At least one of the PAX peptides was detected in all tested strains, except X. poinarii G6 and X. ishibashii DSM 22670 (Table S3), for which no paxTABC cluster was evidenced (Fig. 6). The most conserved PAX peptide in all producing strains is PAX1′. These results revealed that the PAX peptide-producing ability is widespread in the Xenorhabdus genus.
MALDI-TOF-MS detection of selected PAX peptide production in extracts from different strains of Xenorhabdus and colony color on NBTA medium. Orange squares indicate that the m/z value of the corresponding PAX peptide was detected at ±0.1 Da. White squares mean that no m/z value corresponding to PAX peptides was detected. A synthetic PAX peptide derivative DP18 (m/z = 1,038.92 Da) was added as a positive extraction control before the extraction steps. The corresponding mass spectra are given in Table S3. Colony colors were assessed after streaking on NBTA medium and incubating for 48 h at 28°C.
To determine whether a correlation could be established between BBT adsorption and actual PAX peptide production, we examined the color of colonies grown on NBTA medium (Fig. 8). Similarly to the ∆paxA and ∆ngrA mutants (Fig. S3), colonies of X. poinarii G6 and X. ishibashii DSM 22670, lacking the paxTABC cluster and for which no PAX peptides were detected in liquid culture, were red on NBTA medium. Therefore, the Xenorhabdus strains for which PAX peptide production was demonstrated were systematically observed as blue colonies on NBTA medium, whereas red colonies corresponded to strains unable to produce PAX peptides.
DISCUSSION
BBT dye adsorption by bacterial colonies is a marker of PAX peptide production
PAX peptides constitute a family of positively charged cyclolipopeptides, presumably localized at the surface of Xenorhabdus bacteria (36). To investigate the role of PAX peptides, ΔpaxA and ΔngrA mutants were used. Mass spectrometry analysis confirmed the absence of PAX peptide production in both mutants, demonstrating that PAX peptide production in Xenorhabdus is ngrA-dependent. The NgrA PPTase is therefore likely involved in activation of the PaxABC synthetase, as well as other synthetases responsible for the production of NRP metabolites and optimal antibiotic activities in Xenorhabdus (11, 12, 38).
Phenotypic characterization of laboratory cultures revealed similar defects in both ΔpaxA and ΔngrA mutant strains. This suggests that the changes observed in the NRPS-deficient ΔngrA mutant, including the absence of lecithinase-like activity, increased Tween-lipase activity, abolition of BBT adsorption, reduced biofilm formation, and increased swimming motility, are primarily due to the absence of PAX peptides. We showed that both mutants exhibited an enhanced precipitation zone in the agar containing Tween 20, 40, 60, or 80 and abolished lecithinase-like activity. Initially, lecithinase-like activity in X. nematophila F1 was partially characterized through the purification of five compounds that cause lecithin precipitates on agar, without exhibiting phospholipase C activity (39). Given the absence of lecithin precipitates in the ΔpaxA mutant, it is likely that these five compounds correspond to the five PAX peptides described by (28), who employed the same methodology.
Additionally, our study uncovered another striking difference between both mutants and the WT strain, as colonies of both mutants displayed no adsorption of bromothymol blue dye on NBTA medium, unlike the WT strain. Historically, the phenotype of secondary variants has been used to distinguish variants in Xenorhabdus. Indeed, the WT strain shows blue colonies on NBTA medium, while secondary variants exhibit red colonies (40). Initially, this phenomenon was referred to as “phase variation.” Further characterization of X. nematophila secondary variants revealed additional phenotypic traits, such as reduced motility, antibiotic activity, hemolytic activity, and lipolytic activities (41–43). Volgyi et al. proposed the term “phenotypic variation” to more accurately describe this phenomenon (44). Recent studies have shown that secondary variants display a growth advantage in stationary phase both in vitro and in insects but are less efficiently transmitted by the nematode than the WT strain (43). The switch to secondary forms in Xenorhabdus has been linked to mutations in the lrp gene (43), and the expression of the paxAB gene is known to be Lrp-dependent (45). Indeed, an lrp mutant exhibits all the phenotypic traits of a secondary variant, whereas the ΔpaxA mutant shares only some of them, including BBT adsorption, lecithinase-like activity, and abolished PAX production (46). Moreover, Lrp is a global activator that affects the expression of NRPS genes in Xenorhabdus (47). Our mass spectrometry analysis confirmed that Xenorhabdus strains that adsorbed BBT on NBTA medium were able to produce PAX peptides in culture. Conversely, X. poinarii G6, X. ishibashii DSM 22670, and secondary variants form red colonies on NBTA medium, exhibit lecithinase-negative phenotypes (40), and do not produce PAX peptides in culture (Table S3).
Here, we demonstrated that PAX peptides were related to phenotypes involved in phenotypic variation in Xenorhabdus and that the blue phenotype on NBTA medium is a reliable marker for PAX peptide production.
PAX peptides display multiple roles throughout the life cycle of Xenorhabdus
While specialized metabolites produced by soil bacteria have been widely investigated in laboratory conditions for their potential use as antimicrobials in diverse applications, only a few studies have addressed their natural roles, and little is known about their effective occurrence in natural environments and/or in mutualistic relationships with hosts (48–50). Pioneer in vivo studies on Xenorhabdus specialized metabolites demonstrated antimicrobial activity partially attributed to the presence of xenocoumacin in G. mellonella larvae infected with X. nematophila, with detectable levels persisting for up to 10 days post-infection (51).
Similarly, we monitored the presence of Xenorhabdus PAX peptides in the insect cadavers through mass spectrometry analysis in a dynamic way over the course of its natural life cycle. We showed that under simple in vitro conditions, PAX peptides were detected from 20 h to 7 days. Under conditions aiming to reproduce the natural life cycle of X. nematophila in the insect cadaver (G. mellonella infected by X. nematophila or infested by S. carpocapsae), PAX peptides were detected quite uniformly from 48 h to 7 days post-infection/infestation, but higher variability was observed at 20 h between biological samples. Therefore, PAX peptides are present under natural conditions over a long period of the Xenorhabdus life cycle: from the pathogenic phase to the late necrotrophic phase, suggesting that PAX peptides might be involved in various biological functions in Xenorhabdus.
PAX peptides may be involved in crucial phenotypes that enable the bacteria to successfully complete the different steps of their infectious cycle, such as bacterial pathogenicity. In our pathogenicity assays using S. littoralis, we observed only minor differences between the ΔpaxA mutant and the WT strain. Contrary to our observations, delayed mortality of Manduca sexta larvae had previously been observed when infected with a ngrA mutant of X. szentirmaii (38). Moreover, the authors have shown that X. szentirmaii produces specific ngrA-dependent compounds that are absent in X. nematophila and could be involved in virulence (38). Previous studies suggested that PAX peptides may protect Xenorhabdus against the humoral immune response of insects, including AMPs. Because AMPs are cationic peptides, PAX peptides may induce repulsive force due to positive charge at the bacterial cell wall (36). However, we showed that the MICs of several insect AMPs were similar for the WT and ΔpaxA or ΔngrA mutants. The observed differences between our study and previous work (36) can be attributed to variations in bacterial species. Therefore, the potential role of PAX peptides during the pathogenic stage could not be inferred from these experiments.
PAX peptides were originally described as antimicrobial peptides (28); thus, they might play a role in inhibiting the growth of competing microorganisms introduced in the insect hemocoel upon infection by IJs (6, 7), or within the cadaver during the necrotrophic stage. However, we showed that nematode microbiota (FAM) (7) and insect microbiota displayed low susceptibilities to the three synthetic PAXs tested (PAX1′, PAX2′, and PAX7). PAX peptides, therefore, do not appear to be required to outcompete microbial competitors.
PAX peptides could be detected up to 7 days post-infection/infestation in our in vivo experiment (Fig. 1), which is the period when nematodes begin to emerge from the insect cadaver as IJs (Fig. S4). Nematode reproductive success assays in G. mellonella revealed significantly lower progeny using the ΔpaxA and ΔngrA mutants compared to the WT strain, demonstrating that PAX peptides are involved in the production of IJs. Previous studies had shown that ngrA-dependent compounds were required for the development of the nematode partner in two different nematode genera: H. bacteriophora associated with P. luminescens and S. carpocapsae associated with X. nematophila AN6 (12, 13). Our result strongly suggests that PAX peptides are the *ngrA-*dependent metabolites necessary for full IJ production in the X. nematophila-S. carpocapsae mutualism.
As cyclolipopeptides are often identified as involved in biofilm formation, swimming motility, and swarming motility (50), we assessed the involvement of PAX peptides in these phenotypes. Indeed, these features could play a role in successive bacterial colonization of the nematodes in the insect cadaver, prior to IJ emergence. We have highlighted that PAX peptides are positively involved in biofilm formation and negatively involved in swimming motility, but no difference was found in swarming motility. It has been shown that X. nematophila can produce biofilms to adhere to the heads of C. elegans nematodes, while secondary variants that are unable to produce PAX peptides or a biofilm-deficient mutant lack this ability (52). Another study that conducted experimental evolution involving Pseudomonas lurida and C. elegans showed that biofilm formation was a bacterial trait associated with a mutualistic lifestyle and motility associated with a free-living lifestyle (53). In addition, P. lurida produces cyclolipopeptides involved in a mutualistic relationship with the nematode C. elegans (54). We can presume that PAX peptides, which promote biofilm formation, could help the bacterium to associate with its nematode. Other studies revealed that C. elegans nematodes could directly sense cyclolipopeptides (serrawettin W2) produced by harmful bacteria (Serratia marcescens) with their chemosensory neurons, which had a repulsive effect on the nematodes (55). It could be hypothesized that PAX cyclolipopeptides may be sensed by the nematode partner to promote mutualism.
All together, these results revealed that PAX peptides are involved in biofilm formation, swimming motility, and the production of nematode IJs.
The PAX peptide-producing ability is widespread in the genus Xenorhabdus
We highlighted that the paxTABC cluster is present throughout all the Xenorhabdus genus but is restricted to that genus only. A distinct genomic environment around the paxTABC cluster was observed among different Xenorhabdus species, and a high degree of variability in the percentage of identity within the NRPS gene sequences (paxABC) was noticed when blasted to the X. nematophila F1 genome. This variability, due to the high rate of recombination, is characteristic of NRPS genes, unlike genes encoding proteins via the ribosomal pathway (10, 56, 57). We therefore performed the NRPS gene analysis on the modules rather than on the whole genes. Despite the NRPS module’s sequence variability, the seven different strains representative of the main Xenorhabdus species all produced PAX1′ (lysine in position 2), while PAX peptides with an arginine in position 2 (PAX2′ or PAX6) were also detectable in a few strains. In addition to our analysis, the production of PAX peptides has already been confirmed in eight other strains, including X. nematophila ATCC 19061, Xenorhabdus stockiae DSM 17904, Xenorhabdus sp. KJ12.1, Xenorhabdus sp. PB62.4, X. doucetiae FRM16, Xenorhabdus khoisanae SB10, X. khoisanae J194 and X. doucetiae DSM 17909 (9, 32, 33, 36, 58) . Nevertheless, it is important to note that not all PAX peptides are detected in all the analyzed strains, and the only PAX peptide consistently detected is PAX1′. The ability to produce PAX peptides is therefore both widespread and unique to the Xenorhabdus genus.
The only non-PAX-producing species among those tested are X. ishibashii and X. poinarii. X. poinarii has the particularity of being defective in the production of IJs of its nematode partner Steinernema glaseri, being avirulent without its nematode and showing strong genomic reduction (40, 59–61). Moreover, a large proportion of S. glaseri nematodes are naturally aposymbiotic (40). It is conceivable that the absence of PAX peptide production by X. poinarii may contribute to its inefficient association with its nematode partner. Furthermore, in X. nematophila, the paxTABC genes are clustered close to the nilABC genes, whose proteins have been shown to be involved in species-specific colonization of S. carpocapsae (62–65). The nilABC cluster is restricted to the X. nematophila species, whereas the paxTABC genes are present throughout the whole Xenorhabdus genus. We can therefore assume that the PAX peptides unique to the Xenorhabdus genus are metabolites involved in the mutualistic association between the bacterium and its nematode host.
Finally, PAX peptides unique to the Xenorhabdus genus are metabolites involved in many functions such as swimming motility, biofilm formation, and the production of infective juvenile nematodes, suggesting their involvement in the interaction between the bacterium and its nematode host.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions
The strains and plasmids used in this study are listed in Fig. S4. Bacteria were routinely grown in Lysogeny Broth (Sigma-Aldrich) medium at 28°C (Xenorhabdus, S. maltophilia, P. protegens, Ochrobactrum sp*., Achromobacter* sp*., A. faecalis, Pseudochrobactrum* sp*., Brevundimonas* sp*., E. mundtii,* and D. rugosa) or 37°C (Escherichia coli and M. luteus). When required, antibiotics were used at the following final concentrations: gentamicin (Sigma-Aldrich), 20 µg mL^−1^ and chloramphenicol (Sigma-Aldrich), 20 µg mL^−1^ for E. coli strains and 15 µg mL^−1^ for X. nematophila (or 8 µg mL^−1^ for allelic exchange).
Molecular genetic techniques
DNA manipulations were performed as previously described (66). Plasmids were introduced into E. coli by transformation and transferred to X. nematophila by conjugative mating with WM3064 as donor strain (67). All constructs were sequenced by Eurofins Genomics Germany GmbH. The primers used in this study (IDT) are described in Table S5.
Construction of X. nematophila ΔpaxA mutant
The ΔpaxA mutant was constructed using the same experimental strategy as for the construction of the ΔngrA mutant from the WT strain X. nematophila F1 (37). The upstream and downstream regions of the paxA gene were amplified by PCR with the L-PCR1-paxA-SalI and R-PCR1-paxA-BamHI primers for the upstream region (621 bp) and the L-PCR2-paxA-BamHI and R-PCR2-paxA-SpeI primers for the downstream region (601 bp). The two fragments obtained were inserted, together with the 3.8 kb BamHI fragment ΩCam cassette from pHP45-ΩCm, conferring resistance to chloramphenicol, into pJQ200SK digested with SalI and SpeI for insertion of the ΩCam cassette between the two PCR fragments. The resulting plasmid, pJQ-paxA::ΩCm, was used to transform E. coli WM3064 and was introduced into X. nematophila F1 by mating. Allelic exchange was performed as previously described (68). ΩCam insertion was confirmed by PCR analysis using primers L-paxTA and R-verif-pax. The absence of PAX peptide products was assessed by mass spectrometry analysis, as described below. The clone obtained was called ΔpaxA.
Phenotypic assays
Phenotypic characteristics of ΔpaxA and ΔngrA mutants were compared to those of the WT strain by streaking or dropping 5 µL of overnight preculture on the following agar media, as previously described by Boemare et al.: NBTA (Difco Nutrient Agar 1.5% [BD]) supplemented with 25 mg L^−1^ Bromothymol blue (Merck Millipore) and 40 mg L^−1^ triphenyl 2,3,5 tetrazolium (Sigma-Aldrich) (41); Difco Nutrient Agar 1.5% supplemented with 0.1 g L^−1^ CaCl_2_ and 1% Tween 20 (polyoxyethylene sorbitan monolaurate), Tween 40 (polyoxyethylene sorbitan monopalmitate), Tween 60 (polyoxyethylene sorbitan monostearate), Tween 80 (polyoxyethylene sorbitan monooleate), or Tween 85 (polyoxyethylene sorbitan monotrioleate) (Sigma-Aldrich); Difco Nutrient Agar 1.5% supplemented with 1% egg lecithin (VWR Avantor); Trypticase Soy Agar (BioMerieux) supplemented with 7% sheep blood (Eurobio Scientific) (69); Difco Nutrient Agar supplemented with a 6 g L^−1^ soft agar overlay containing 2% M. luteus preculture; 0.35% LB Agar medium for swimming motility assays (70); and LB supplemented with 0.7% of Eiken Agar (Gerbu) for swarming motility assays (71).
Biofilm assays
Crystal violet biofilm assays were carried out following the methods described by Pothula et al., using LB broth instead of LPB medium (72). Briefly, cultures grown for 48 h without agitation were stained with 1% crystal violet (Sigma-Aldrich), then dissolved in 30% acetic acid after washing steps. The absorbance of samples in a 96-well plate was quantified at 550 nm in a TECAN Infinite 200 plate reader. Each experiment was conducted with four independent clones and repeated four times for each mutant strain. Data were analyzed using GraphPad Prism 9 software, and significance was tested using Student’s test.
MIC determination
MICs were determined in accordance with CLSI guidelines (73) with the following modifications: 5 mL of MHB broth (Bio-Rad) was inoculated with an overnight culture in LB broth and grown to a 0.6 < OD_600_ < 0.9. The inoculum for the microplate was prepared as described in the CLSI guide, with dilution in MHB to approximately 10^4^ CFU mL^−1^. Ninety-six-well low-binding plates were incubated for 48 h at 28°C with slow shaking. The peptides used in the analyses were supplied by the following providers: cecropin A (Sigma-Aldrich), cecropin B1 (NeoMPS S.A.) (74), cecropin C (Proteogenix), cecropin P1 (Sigma-Aldrich), drosocin (Eurogentec), colistin (Sigma-Aldrich), polymyxin B (Sigma-Aldrich), and NOSO 95C (odilorhabdin, Nosopharm S.A. [16]). Synthetic PAX1′, PAX2′, and PAX7 were provided by the SynBio3 platform (IBMM, Montpellier, France), and their structure is shown in Fig. S1.
Pathogenicity assays
Bacterial pathogenicity was assessed by injecting Xenorhabdus into S. littoralis, as previously described (75). Briefly, bacterial cultures in LB broth (OD_600_ ≈ 0.8) were diluted in the culture medium, and 20 µL of the resulting bacterial suspension, containing 1,000–3,000 CFUs, was injected into the hemolymph of 20 sixth-instar larvae of S. littoralis. The number of bacterial cells injected into the larvae was determined by plating on nutrient agar and counting the CFUs. After the bacterial injection, the insect larvae were incubated at 23°C, and mortalities were monitored for up to 50 h. The pathogenicity of bacterial isolates was determined by measuring the time required for 50% of the insect larvae to be killed (LT_50_). The results of four independent experiments were combined and analyzed with the software Statistical Package for the Social Sciences version 18.0. Significant differences between two data sets were assessed with non-parametric Wilcoxon-Mann-Whitney tests, with a 95% confidence interval.
Nematode colonization assays
Bacterium–nematode complexes were produced using an in vivo technique (2, 76) in which G. mellonella (wax moth) larvae were infected with X. nematophila F1 strain and aposymbiotic IJs of S. carpocapsae (SK27, Plougastel) nematodes. Aposymbiotic nematodes were obtained previously (77). Approximately 100 aposymbiotic IJs per larvae were first placed on a piece of filter paper in individual Eppendorf tubes and incubated with 20 larvae of G. mellonella at 23°C. After 24 h of infestation, insect larvae were injected with 20 µL (~3,000 bacteria) of WT, ΔpaxA, or ΔngrA strains that had been grown until OD_600_ ≈ 0.8. After insect death (24–48 h post-infestation), cadavers were transferred to White traps (78). Fourteen and thirty days after infestation, the IJs emerging from the cadavers were collected in Ringer’s solution (Aguettant). Once again, these IJs were then used to infest new G. mellonella larvae. Nematodes were collected and washed with water through a 20 µm filter and stored in Ringer’s solution at 9°C in tissue culture flasks. Reproductive success (number of IJ nematodes emerging from an insect larva) was determined by cumulating the number of nematodes collected and counted at 15 and 30 days post-infestation. Direct counts of average CFU/nematode were determined via a grinding assay, wherein collected nematodes were equalized by density, serially diluted, and plated on NBTA plates (Fig. S6). Each experiment was repeated three times. Data were analyzed using GraphPad Prism 9 software, and significance was tested using non-parametric Wilcoxon-Mann-Whitney tests.
Mass spectrometry analysis
PAX peptide extraction from Xenorhabdus cultures
PAX extraction methods were adapted from those previously described by (28, 32, 36). Tubes of 5 mL LB were inoculated with 200 µL preculture of overnight Xenorhabdus strains and grown for 48 h at 28°C under agitation. Cultures were centrifuged at 6,000 × g for 10 min, and the supernatant fraction was discarded. The cell pellet fraction was resuspended in 1 mL LB, and 45 µg of DP18 (R-3-Hydroxytetradecanoic acid-Gly-Orn-[DLys-Lys-DLys-DLys-Lys], m/z = 1,038.92 Da), a synthetic PAX peptide derivative, was added as an extraction control. Samples were sonicated for 2 min, then centrifuged at 16,000 × g for 5 min. The supernatant corresponding to the cytosolic fraction (S1) was conserved. The new pellet containing membrane debris was resuspended in 500 µL methanol/H_2_O (vol/vol) and acidified with 1% formic acid. Samples were incubated for 5 min in the ultrasonic bath. The insoluble fractions were then separated by centrifugation (16,000 × g, 5 min). The resulting supernatant S2 was pooled with supernatant S1 and then filtered with 0.45 µm filters (Filtropur, Sarstedt). Samples were suspended at a 1:1 ratio in 0.1 M NaCl, 0.02 M Tris, pH 9 (vol/vol), then loaded onto Sep-Pak CarboxyMethyl Short Cartridge (Accell Plus CM, Waters) prepared according to the manufacturer’s instructions. Cartridges were eluted with 0.5 M NaCl, 0.02 M Tris, pH 9, and 0.1% trifluoroacetic acid. The resulting peptide extract was then loaded onto Sep-Pak C18 Short cartridge (Sep-Pak Plus C18, Waters) prepared according to the manufacturer’s instructions. Peptides were eluted with 100% acetonitrile (ACN) and freeze-dried.
Preparation of in vivo samples
G. mellonella larvae were injected with 5 × 10^3^ to 7 × 10^3^ CFUs of X. nematophila F1 (as described in Pathogenicity assays) or infested with 100 S. carpocapsae SK27 IJ nematodes (as described in Nematode colonization assays) and incubated at 23°C. Larvae were weighed and frozen at −20°C at each time point post-injection/infestation: 20 h, 48 h, 72 h, 5 days, and 7 days. Larvae surfaces were first disinfected with 70% ethanol, after which the heads were removed. Insect forceps were then used to extrude internal contents. The insides of 10 larvae per condition were pooled and added to 2 mL 0.05 M Tris, pH 7, before grinding using FastPrep-24 5G (MP Biomedicals) with sterile glass beads (6.5 m/s, 30 s, three times). Samples were centrifuged at 6,000 × g for 30 min, and cell pellets were resuspended in 1.5 mL 0.05 M Tris, pH 7. PAX peptide extraction was then performed as described above, starting from the step of DP18 addition to the samples. Experiments were performed in triplicate.
MALDI-TOF-MS analysis
Extracted peptides were analyzed with a RapifleX MALDI TOF/TOF (Bruker) equipped with a Pulse Smart Beam 3D laser at 335 nm. The following instrument parameters were used: frequency, 10,000 Hz; delayed extraction time, 160 ns; source, positive mode; reflectron mode. Samples were resuspended in ACN and mixed at a 1:1 ratio with 1 µL of saturated α-cyano-4-hydroxycinnamic acid. A volume of 1 µL of the mix was spotted onto a stainless-steel target and air-dried. A targeted analysis of the spectra was carried out on compounds with m/z ratios between 1,000 and 1,200, and corresponding monoisotopic mass lists were generated using Flex Analysis 3.4 (Bruker).
NRPS genomic analysis
The paxTABC and surrounding gene sequences of 40 strains among 23 Xenorhabdus species were extracted from the MicroScope MaGe database (https://mage.genoscope.cns.fr/microscope/home/index.php). Module sequences of NRPS genes (paxA: m1; paxB: m2, m3, and m4; paxC: m5, m6, and m7) were extracted using the antiSMASH tool implemented in the MicroScope MaGe database. Strains with genomes of poor assembly quality (>350 contigs) were removed from the analysis. A blastn analysis using the NCBI BLAST+ tool implemented on the Galaxy version 2.10.0+ galaxy1 platform (Galaxy Pasteur, https://galaxy.pasteur.fr/) was performed using the sequence of X. nematophila F1 as query with a cutoff value of 0.1 and a minimum coverage percentage >30%. Percentage identities have been used to construct Fig. 6 using the ggplot2 package on RStudio version 2024.09.1+394. Details of the data used for these analyses can be found in Tables S6 and S7.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Goodrich-Blair H, Clarke DJ. 2007. Mutualism and pathogenesis in Xenorhabdus and Photorhabdus: two roads to the same destination. Mol Microbiol 64:260–268. doi:10.1111/j.1365-2958.2007.05671.x 17493120 · doi ↗ · pubmed ↗
- 2Sicard M, Brugirard-Ricaud K, Pagès S, Lanois A, Boemare NE, Brehélin M, Givaudan A. 2004. Stages of infection during the tripartite interaction between Xenorhabdus nematophila, its nematode vector, and insect hosts. Appl Environ Microbiol 70:6473–6480. doi:10.1128/AEM.70.11.6473-6480.200415528508 PMC 525208 · doi ↗ · pubmed ↗
- 3Stock SP. 2019. Partners in crime: symbiont-assisted resource acquisition in Steinernema entomopathogenic nematodes. Curr Opin Insect Sci 32:22–27. doi:10.1016/j.cois.2018.10.00631113627 · doi ↗ · pubmed ↗
- 4Nguyen KB, Hunt DJ, Mráček Z. 2007. Steinernematidae: species descriptions, p 121–609. In Entomopathogenic Nematodes: Systematics, Phylogeny, and Bacterial Symbionts. Vol. 5. Nematology Monographs and Perspectives. Koninklijke Brill BV, Leiden, Netherlands.
- 5Walsh KT, Webster JM. 2003. Interaction of microbial populations in Steinernema (Steinernematidae, Nematoda) infected Galleria mellonella larvae. J Invertebr Pathol 83:91–96. doi:10.1016/s 0022-2011(03)00079-x 12788281 · doi ↗ · pubmed ↗
- 6Singh S, Reese JM, Casanova-Torres AM, Goodrich-Blair H, Forst S. 2014. Microbial population dynamics in the hemolymph of Manduca sexta infected with Xenorhabdus nematophila and the entomopathogenic nematode Steinernema carpocapsae. Appl Environ Microbiol 80:4277–4285. doi:10.1128/AEM.00768-1424814780 PMC 4068695 · doi ↗ · pubmed ↗
- 7Ogier JC, Pagès S, Frayssinet M, Gaudriault S. 2020. Entomopathogenic nematode-associated microbiota: from monoxenic paradigm to pathobiome. Microbiome 8:25. doi:10.1186/s 40168-020-00800-532093774 PMC 7041241 · doi ↗ · pubmed ↗
- 8Lacey LA, Grzywacz D, Shapiro-Ilan DI, Frutos R, Brownbridge M, Goettel MS. 2015. Insect pathogens as biological control agents: Back to the future. J Invertebr Pathol 132:1–41. doi:10.1016/j.jip.2015.07.00926225455 · doi ↗ · pubmed ↗