Evidence of ESBL plasmid transfer and selective persistence of multiple host-associated Escherichia coli isolates in a chicken cecal fermentation model
J. Leng, M. Ferrandis-Vila, R. Oldenkamp, J. W. Mehat, A. S. Fivian-Hughes, S. Kumar Tiwari, B. Van der Putten, V. Trung Nguyen, A. Bethe, J. Clark, P. Singh, T. Semmler, S. Schwarz, J. Alvarez, N. T. Hoa, M. Bootsma, C. Menge, C. Berens, C. Schultsz, J. M. Ritchie

TL;DR
This study explores how different Escherichia coli isolates from various hosts survive and transfer antibiotic resistance in a chicken gut model.
Contribution
The study reveals isolate-specific genomic differences influence persistence and plasmid transfer of antibiotic resistance in complex gut environments.
Findings
Pig and cattle-associated E. coli isolates persisted longer in the chicken gut model.
Most recovered ESBL E. coli acquired a blaCTX-M-1 plasmid from a generalist isolate.
Higher inoculum doses or ceftiofur addition helped stabilize ESBL E. coli populations.
Abstract
The guts of animals and humans harbor diverse microbial communities that are regularly exposed to bacteria originating from food, water, and their surroundings. Species such as Escherichia coli are adept at colonizing multiple hosts, along with surviving in the environment. By encoding pathogenic traits and transmissible forms of antimicrobial resistance (AMR), E. coli can also pose a zoonotic risk. Our understanding of the factors that govern host residency is limited. Here, we used a chicken cecal fermentation model to study survival and the AMR transfer potential of 17 host-associated extended-spectrum β-lactamase (ESBL)-producing E. coli isolates. Vessels containing chicken cecal contents were stabilized for 4 days before the addition of a cocktail comprising ESBL-producing E. coli obtained from human, cattle, pig, and chicken hosts. Consecutive sampling showed that pig and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
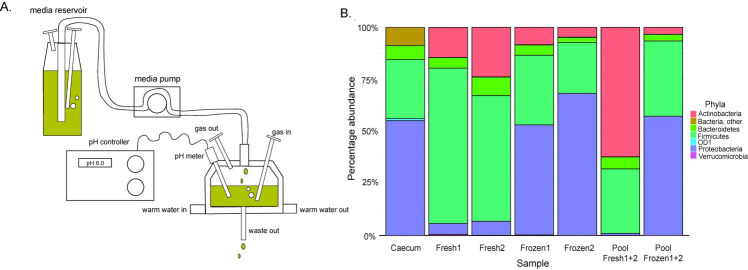 Fig 1
Fig 1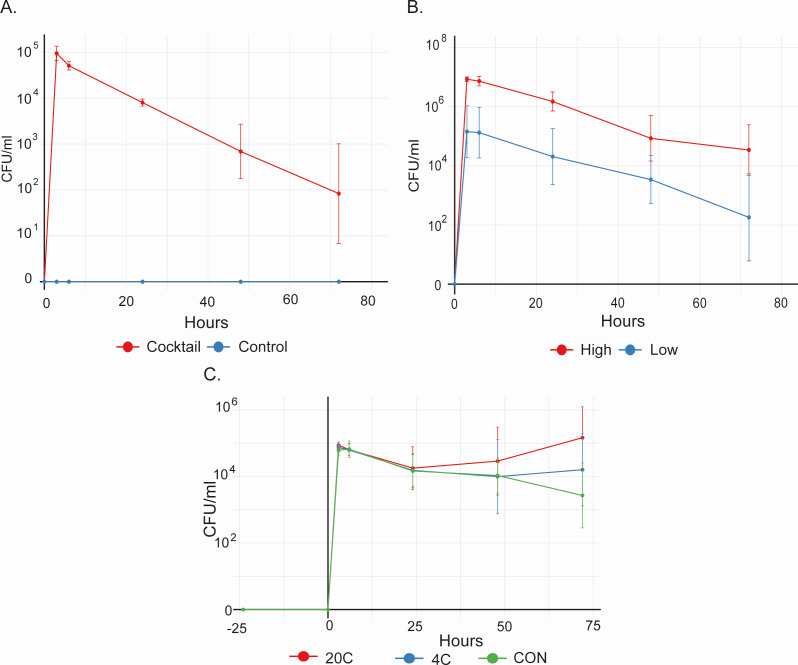 Fig 2
Fig 2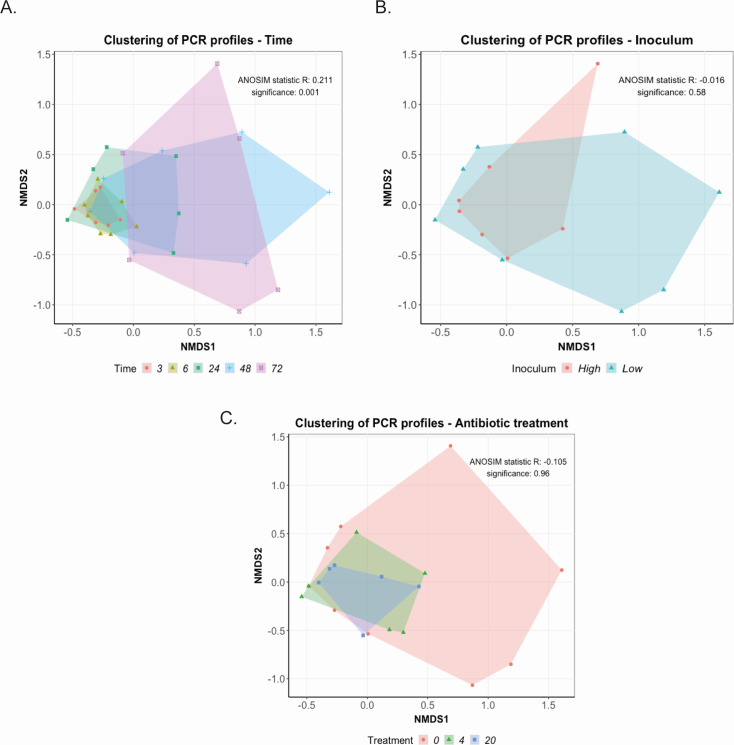 Fig 3
Fig 3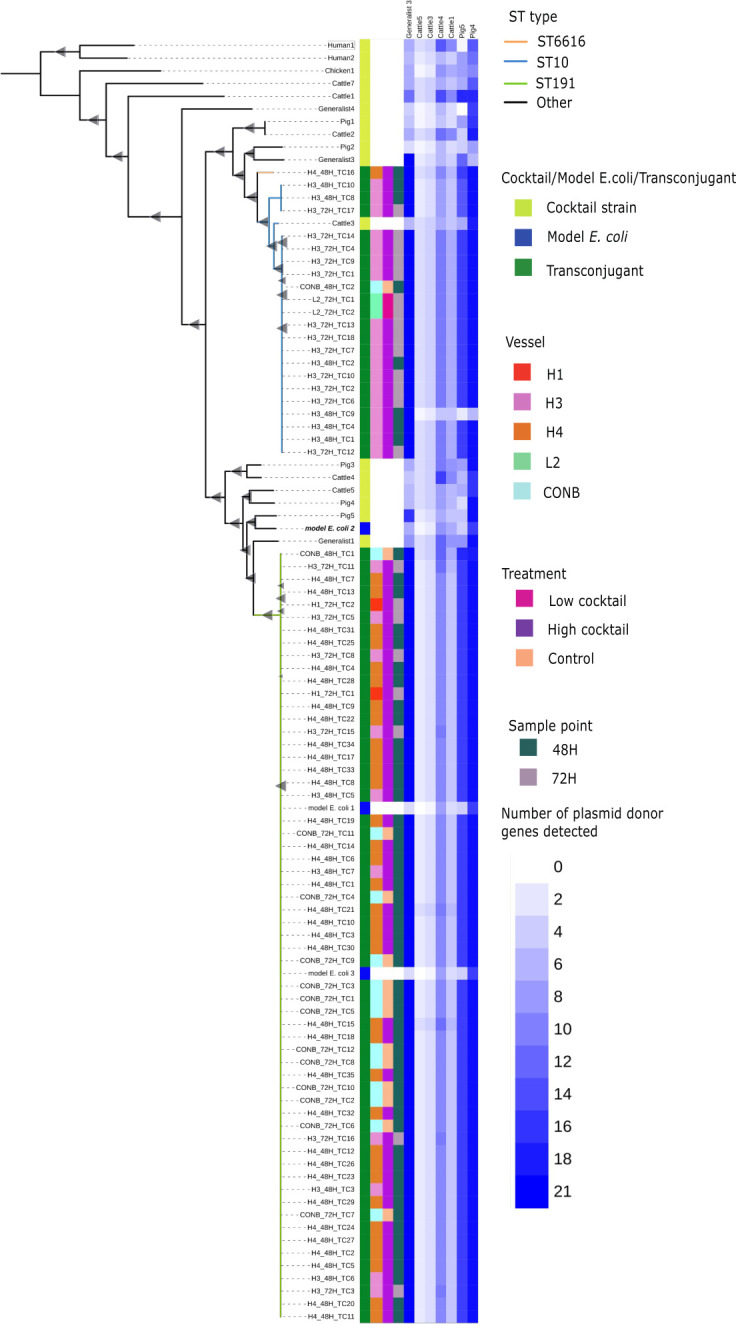 Fig 4
Fig 4| Isolate ID | Source | Country of origin | Health status | Year | Phylogroup | Sequence type (ST) | Resistance genes |
|---|---|---|---|---|---|---|---|
| Chicken1 | Chicken | Vietnam | Healthy | 2013 | F | ST1163 | |
| Human1 | Human | UK | Disease | 2017 | B2 | ST131 | |
| Human2 | Human | UK | Disease | 2017 | B2 | ST636 | |
| Cattle1 | Cattle | Germany | Disease | 2016 | D | ST362 | |
| Cattle2 | Cattle | Germany | Healthy | 2015 | A | ST361 | |
| Cattle3 | Cattle | Germany | Disease | 2007 | A | ST10 | |
| Cattle4 | Cattle | Germany | Disease | 2014 | C | ST88 | |
| Cattle5 | Cattle | Germany | Disease | 2005 | B1 | ST448 | |
| Cattle7 | Cattle | Germany | Healthy | 2011 | F | ST648 | |
| Pig1 | Pig | Germany | Disease | 2016 | A | ST361 | |
| Pig2 | Pig | Germany | Disease | 2011 | A | ST206 | |
| Pig3 | Pig | Germany | Healthy | 2011 | C | ST410 | |
| Pig4 | Pig | Germany | Healthy | 2016 | B1 | ST641 | |
| Pig5 | Pig | Germany | Healthy | 2016 | B1 | ST2067 | |
| Generalist4 | Chicken | Spain | Healthy | 2016 | D | ST1011 | |
| Generalist3 | Human | Germany | Disease | 2009 | A | ST12717 | |
| Generalist1 | Chicken | Vietnam | Healthy | 2013 | B1 | ST162 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Vibrio bacteria research studies · Salmonella and Campylobacter epidemiology
INTRODUCTION
The gut microbiota of animals and humans is regularly exposed to bacteria originating from food, water, the environment, or other hosts (1, 2). Some microorganisms behave as transient visitors that are rapidly lost, whereas others establish and become part of the resident microbiota (3, 4). Phenotypic traits such as motility (5), metabolic adaptability (6), and adaptation to substances such as bile (7) likely contribute to residency. Moreover, bacterial defense strategies, including the ability to produce bacteriocins, could help isolates compete within a host (8). Individual replication rates and an ability to acquire or lose genetic material may also confer advantages over less successful bacteria (9). Finally, inter- and intra-species interactions could impact population dynamics, with individual populations negatively affected by competition for resources (10) or positively affected by cross-feeding (11).
Escherichia coli is a well-known inhabitant of the human and animal intestine, where it may exist as a commensal or pathogenic member of the microbiota (12). There is growing interest in E. coli population dynamics due to its propensity for horizontal transfer of antimicrobial resistance (AMR) genes, its prevalence and pathogenesis in multiple hosts, and its survival in the natural environment. Yet the mechanisms determining whether E. coli from one host establishes in a new host species are not clear. Several studies suggest that certain clones of E. coli exhibit host preference so that they are more likely to become resident within the human intestine (4, 13). Moreover, high-resolution, isolate-level analyses of E. coli originating from humans and animals indicate that the two populations are genetically distinct (14–17). Consistent with this, genome-wide association studies (18) and logistic regression analysis of single nucleotide polymorphisms (19) have found evidence for host-specific determinants, although data from experimental mixed-isolate infections is generally lacking.
Resistance to third-generation cephalosporins due to plasmid-encoded extended-spectrum β-lactamases (ESBLs) is commonly detected in bacteria present in the mammalian gastrointestinal tract (20). For example, 16.5% of healthy individuals present in the community (21) and 10%–45% of pigs and cattle (22–27) were found to harbor ESBL E. coli. The zoonotic risks associated with ESBL E. coli from different hosts are not clear, and quantitative data on the potential for plasmid transfer in the gastrointestinal system are limited. Transfer of ESBL-encoding plasmids from avian-derived E. coli isolates to human commensal E. coli has previously been observed using in vitro models of the human gastrointestinal system (28, 29), but data from food-producing animal models are rare.
In vitro models using feces/gut content from humans or animals can offer a reproducible and controlled way to study the bacterial populations in the gut without the need for in vivo animal models. Here, we utilized a chicken cecal fermentation model to simultaneously monitor the ability of 17 previously described host-associated ESBL E. coli (18) to persist in the presence and absence of antimicrobial selection pressure and used whole-genome sequencing to determine which isolate(s) transferred their ESBL-encoding plasmid to the native E. coli present in the community. Collectively, our data indicated that the chicken-associated isolate was no better at persisting than isolates associated with cattle, pigs, or humans, and that an isolate lacking host-specific traits (18) appeared largely responsible for the spread of an ESBL plasmid to native E. coli within the cecal community.
RESULTS
Validating the use of pooled chicken cecal content as a model inoculum
We used a previously described single-stage, continuous flow chicken cecal fermentation model (30) in our study (Fig. 1A). To facilitate model operation, we evaluated whether frozen, pooled chicken cecal content could replace the use of freshly collected content as an inoculum. As can be seen in Fig. 1B, the bacterial composition in vessels seeded with frozen cecal samples, individual or pooled, after 4 days of incubation, more closely resembled the profile in the cecum than when fresh content was used. For example, the individual vessels inoculated with frozen material contained 52%–68% Proteobacteria and 24%–36% Firmicutes, which were comparable to the proportions identified in the ceca (55% and 28%, respectively). In contrast, the two vessels inoculated with fresh cecal samples had 60%–75% Firmicutes, while Actinobacteria, which were identified as <1% in the ceca, increased to 62% in the pooled fresh sample. The bacterial class composition of samples from frozen pooled chicken ceca was also observed to be most similar to those identified in the cecal samples (Item S1). The relative abundance of the three most common bacterial classes identified in the models inoculated with pooled frozen cecal content was 40% Clostridia, 23% Gammaproteobacteria, and 24% Alphaproteobacteria. This was comparable to 28% Clostridia, 24% Gammaproteobacteria, and 28% Alphaproteobacteria identified in the chicken cecal samples analyzed. Thus, frozen pooled chicken cecal content was used as an inoculum in all subsequent experiments.
Overview and microbial content of a continuous flow chicken cecal fermentation model. (A) Schematic representation of the single vessel model maintained under anaerobic conditions with automatic adjustment for pH and temperature. (B) Bacterial composition based on 16S rRNA gene sequencing of samples from the chicken ceca and from the model after 4 days of fermentation. Bars represent the mean percentage abundance for bacterial phyla found in the chicken cecum (N = 3), two vessels inoculated with fresh cecal content from two independent chickens (Fresh1, Fresh2), two vessels inoculated with content Fresh1 and Fresh2 combined (Pool Fresh 1+2), vessels inoculated with cecal content from independent chickens, frozen at the time of collection and subsequently re-used (Frozen1 and Frozen2), and two vessels inoculated with a frozen pooled sample (Pool Frozen 1+2). Duplicate vessels were run for each condition.
Immigrant E. coli isolates were outcompeted by the native cecal community
We previously described a panel of 17 individual ESBL-producing E. coli isolates, hereafter referred to as the E. coli cocktail, isolated from a range of hosts and each phylogenetically classified as associated with a particular host (30, Table 1). Herein, rifampicin-resistant (RIF^R^) mutants of the E. coli cocktail (10^8^ total CFU) were simultaneously introduced into the chicken cecal fermentation model to assess the ability of individual isolates to persist. Using selective media, we found that the concentration of total bacteria and Enterobacteriaceae recovered on LB and Gassner agar, respectively, remained relatively constant in three different experiments (Items S2A and S2B), while the mean concentration of the ceftiofur-resistant (CTF^R^) bacteria decreased by 2–3 logs (Item S2C). The concentration of E. coli cocktail bacteria (identified based on CTF^R^ and RIF^R^) similarly declined (Fig. 2A) with isolates only appearing to stabilize in vessel 4 (V4) between 48 and 72 h post-inoculation (Item S3A). No CTF^R^ organisms were recovered from control vessels that did not receive the E. coli cocktail. This indicated that most immigrant E. coli isolates were unable to be stably maintained in the cecal fermentation model. In our second experiment, increasing the inoculum dose to 10^10^ total CFU E. coli cocktail bacteria led to the recovery of approximately 2 log higher concentrations of CTF^R^ bacteria over time (Fig. 2B; Item S2C), with similar increases in the recovery of total bacteria and Enterobacteriaceae (Item S2A and S2B). Notably, concentrations of E. coli cocktail bacteria now appeared to stabilize in half of the vessels (H3, H4, and H6), with concentrations plateauing by 48 and 72 h (Item S3B). The higher concentration of CTF^R^ bacteria compared to E. coli cocktail bacteria in some vessels (H3 and H4, Item S3B) was suggestive of ESBL plasmid transmission to native bacteria. Finally, the addition of both low and high concentrations of CTF increased the recovery of immigrant E. coli over time (Fig. 3C), while the number of total bacteria and Enterobacteriaceae remained highest in control vessels that did not receive any antibiotics (Items S2A and S2B).
Effect of dose and ceftiofur on the recovery of E. coli cocktail bacteria (CFU/mL). A 17-isolate E. coli cocktail was added to a continuous flow vessel inoculated with chicken cecal contents. Bacteria were recovered following serial dilution and growth on Gassner media supplemented with rifampicin (50 µg/mL) and ceftiofur (4 µg/mL). (A) First experiment—vessels were dosed with 108 CFU E. coli cocktail or sterile phosphate-buffered saline (PBS) (control), N = 6 vessels each. (B) Second experiment—vessels were dosed with 1010 CFU (high dose) or 108 CFU (low dose) E. coli cocktail, N = 6 vessels each. (C) Third experiment—vessels were treated with ceftiofur at 20 µg/mL (20C), 4 µg/mL (4C), or PBS (no antibiotic control, CON) 24 h before, immediately prior to, and 24 h after dosing with E. coli cocktail (108 CFU), N = 4 vessels each.
Non-metric multidimensional scaling (NMDS) plots showing changes in E. coli cocktail composition in the chicken cecal fermentation model. Continuous flow vessels containing chicken cecal contents were inoculated with a 17-member E. coli cocktail, and changes in the cocktail composition were determined using a PCR-based analysis following DNA extraction of bacterial lawns recovered on Gassner media supplemented with rifampicin (50 µg/mL) and ceftiofur (4 µg/mL). PCR presence or absence data were analyzed by NMDS to show changes in composition: (A) over time at 3, 6, 24, 48, and 72 h post-inoculation, (B) at high (1010 CFU) or low (108 CFU) doses of the cocktail, and (C) following the addition of ceftiofur (at 0, 4, or 20 µg/mL). ANOSIM was used to test for statistical differences between groups. N = 4 to 6 biological replicates per group.
Maintenance of immigrant E. coli isolates in the chicken cecal fermentation model was independent of designated host association
Having established parameters facilitating transient maintenance of immigrant isolates in the chicken cecal fermentation model, we sought to quantify the survival of individual members within the E. coli cocktail. Using a PCR-based approach (31), we found that E. coli associated with cattle (isolates cattle2 and cattle7) and pigs (isolate pig1) were maintained in at least half of the vessels over the 72 h incubation period (Item S4). Conversely, we found that some isolates were eliminated by 72 h (human2, cattle1, and pig3) or failed to establish in the vessels (generalist4). The single chicken-associated isolate (chicken1) did not appear to survive better than other host-associated isolates and was undetectable in most vessels by 72 h. Similar patterns were seen when the total number of bacteria in the E. coli cocktail was increased to 10^10^ total CFU and, for some isolates (chicken1, cattle3, cattle7, pig3, pig4, and generalist1), this change in concentration led to an increase in the number of vessels in which they could be recovered (Item S5). Interestingly, the addition of CTF did not result in consistent overgrowth of the E. coli cocktail, as few isolates were retained in all vessels. The isolates chicken1 and human1 were exceptions to this. However, they were also retained in untreated control vessels, suggesting that this was a community- or isolate-mediated effect rather than due to the presence of the antibiotic. Similarly, isolates cattle1, cattle5, and pig1 were retained in a higher percentage of control vessels, while cattle4 was retained in a higher percentage of vessels treated with a low level of the antibiotic (Item S6). Overall, adding CTF had no clear effect on which isolates were able to persist and for how long.
The PCR data facilitated the analysis of the changes in the composition of the E. coli cocktail (absence/presence) over time using a non-metric multi-dimensional scaling (NMDS) model. Time had a significant effect on the profile of isolates present in the cecal fermentation model (P = 0.001, Fig. 3A), with E. coli communities becoming more dissimilar as the incubation progressed. Isolate profiles at the start of the incubation differed from those at 72 h, where there was also greater variation in the vessels. However, analysis of NMDS plots showed that there was no significant effect of inoculum dose or antibiotic treatment on cocktail composition (P > 0.05, Fig. 3B and C), suggesting that the divergence of the E. coli isolate composition in each vessel is most likely due to uncharacterized inter- or intra-species interactions.
Immigrant isolates were able to transfer ESBL plasmids to native E. coli
As previously mentioned, the difference in CFU counts of CTF^R^ and CTF^R^ RIF^R^ microorganisms was indicative of ESBL plasmid transfer to native bacteria. Thus, we used replica plating and short-read whole-genome sequencing to determine the genotype of recipient E. coli and the likely origin of the plasmid. A total of 81 putative transconjugants were recovered from the following vessels over the three experiments: 10 from H3 at 48 h, 35 from H4 at 48 h, 2 from H1 at 72 h, 18 from H3 at 72 h, 2 from L2 at 72 h (from the second experiment), 2 from CON B at 48 h and 12 from CON B at 72 h (from the third experiment). In addition, three native E. coli isolated from chicken cecal content before cocktail addition were included in the analyses. Sequencing confirmed that transconjugants were E. coli belonging to sequence type (ST) 191 (61 isolates), ST10 (21 isolates), and ST6616 (1 isolate) (Item S7). Phylogenetic reconstruction of the 17 isolates from the E. coli cocktail, 3 native E. coli recovered from the fermentation mix, and 81 transconjugants showed that transconjugants were phylogenetically distinct from donor isolates (Fig. 4).
Phylogenetic relationship of resistant E. coli, three native E. coli present in the chicken ceca fermentation model, and the E. coli cocktail isolates. Continuous flow vessels containing chicken cecal contents were inoculated with a 17-member ESBL-producing E. coli cocktail in different experiments. Bacteria that were ceftiofur-resistant (CTFR), but rifampicin-sensitive (RIFS), were subject to whole-genome sequencing. Abricate was used to generate resistance gene profiles that led to seven members of the E. coli cocktail being identified as putative donors. The heatmap shows the number of genes in seven potential donor isolates that were identical to the 10-gene region on either side of blaCTX-M-1 in the putative transconjugant (score out of 21). Key: ST sequence type; Vessel H (20 µg/mL ceftiofur), L (4 µg/mL ceftiofur), C (no antibiotic control); treatment: high dose cocktail, low dose cocktail, control; sample point time post-inoculation.
A total of 11 AMR genes, with blaCTX-M-1 being the most common, were identified in the potential transconjugants. Three combinations of resistance genes within the transconjugants persisted at higher frequencies and were recovered from multiple different vessels: (i) aadA1, aadA2, blaCTX-M-1, blaECblaTEM-1, cmlA1, Inu(G), and sul3 (17 of 81 isolates), (ii) aadA22, blaCTX-M-1, blaEC-18, Inu(G), and tet(B) (47 of 81 isolates), and (iii) aadA22, blaCTX-M-1, blaEC-18, blaTEM-1, Inu(G), and tet(B) (13 of 81 isolates). There were four other isolates that did not fall into these three distinct groups; three of these formed another small clonal group, and the last (H4 48H TC16) was not genetically similar to any other isolate.
We noted that the blaCTX-M-1 gene was found in all transconjugants but absent from the native E. coli present in the cecal content used to seed the models, indicating that this was a marker of plasmid transfer. To identify which member(s) of the E. coli cocktail had shared their plasmid, the up- and down-stream regions adjacent to this locus were compared in the cocktail isolates that harbored blaCTX-M-1. Specifically, a custom BLAST database of the 10 genes immediately up- and down-stream of blaCTX-M-1 was compiled using generalist3, cattle1, cattle3, cattle4, cattle5, pig4, and pig5 sequences. All 21 genes associated with generalist3 were identified in all transconjugants (Fig. 4), implicating this isolate as the most likely donor for most transconjugants. While a high proportion of genes present in isolate pig4 were also detected in the transconjugants (20 out of 21 genes), alignment of the blaCTX-M-1 regions in pig4, generalist3, and a representative transconjugant revealed that only three genes immediately adjacent to blaCTX-M-1 were identical with the remaining 16 genes encoded elsewhere in the genome (Item S8). Additionally, these genes were also found in most other E. coli we sequenced, including model E. coli ST191 recipients initially isolated from cecal contents.
DISCUSSION
Although the digestive tract of vertebrates often harbors multiple isolates of the same species at the same time (32), few studies have attempted to compare their relative persistence or ability to contribute to AMR transmission. Here, employing a chicken cecal fermentation model, we were able to track the dynamics of multiple ESBL-producing E. coli isolates residing simultaneously in a complex community and provide data identifying the ESBL-harboring isolate(s) most likely to have contributed to AMR transmission to native E. coli residing in the community.
Our data indicated that not all immigrant E. coli isolates persisted to the same extent. In our experiments, some pig- and cattle-associated isolates were recovered for longer periods than those associated with chicken or those lacking specific host-associated traits (generalists). Given that equivalent numbers of cells were added to each isolate, these data demonstrate that for some of these naturally occurring isolates, host exposure is less likely to result in establishment in the intestine (4, 13). In addition, it appears that host association, determined for these isolates through a combined analysis of their phylogeny and source metadata (18), did not necessarily restrict isolates associated with other hosts from establishing in the chicken cecum (19) or guarantee an ability for chicken-associated isolates to re-establish in the same host species. Interestingly, isolates belonging to phylogroups A (cattle2, pig1) and F (cattle7) were among those most often recovered, consistent with findings from healthy adult volunteers (4, 13).
The factors that govern residency remain poorly understood. Colonization resistance (33), a phenomenon conferred by the native microbiota on immigrant bacteria (34), is widely acknowledged. In addition, studies describing the properties of resident isolates can offer a glimpse into the attributes that may be important for successful immigrant isolates (4). Yet despite this knowledge, our ability to predict which immigrant isolates take up residence is limited. Determining this would go a long way to support a fuller understanding of zoonotic potential, transmission risk, and exploitation of probiotic therapies.
Dosing the vessels with ceftiofur helped to maintain immigrant populations, yet this did not result in consistent outgrowth of all resistant bacteria, challenging the assumption that resistant isolates typically outcompete susceptible isolates in the presence of a selective agent (35). It is possible that the acquisition of the ESBL-encoding plasmid reduced the fitness of the putative transconjugants (36, 37), a phenotype that can be exacerbated in the presence of a community (38). For example, a 20% reduction in bacterial growth rates was attributed to loss of the porins OmpC/OmpF following ESBL plasmid acquisition in E. coli (39). Whether this explains our inability to recover putative transconjugants from vessel H4 at 72 h, despite finding a large number (35 of 81 recovered in total) at earlier time points, is not known.
In addition to community effects on isolate fitness, it is possible that the presence of other microorganisms affected how members of the E. coli cocktail responded to antibiotics. Recent studies using both simple (two species [40]) and more complex (gut microbiotas [41]) microbial communities found that the growth and evolution of AMR in immigrant lineages were suppressed when other organisms were present. There is now a growing body of evidence that interspecies interactions can alter responses to antibiotics, and as such, the ecological context in which AMR is found must be considered when designing effective antibiotic stewardship criteria for both clinical and natural settings (42).
Finally, our data suggest that ESBL plasmid acquisition by native E. coli residing in the community occurred largely from a single representative, generalist3, a non-host-associated E. coli belonging to phylogroup A. Before we assessed the ability of isolates to conjugate in the chicken ceca model, we tested all isolates in individual mating experiments. This showed that all cocktail isolates had the ability to transfer their ESBL genes, conferring beta-lactam resistance to another E. coli in vitro (data not shown here). Surprisingly, generalist3 was not the most persistent cocktail isolate recovered in vessels where newly resistant isolates were detected. However, it should be noted that our data do not allow the absolute levels (CFU/g) of individual isolates to be determined, only their presence or absence. Thus, it is possible that this strain persisted at high densities during the initial stages of incubation before declining below detection limits. There is also a possibility that other gene transfer mechanisms occurred. For example, two of the three resistance gene profile groups mentioned previously included both blaCTX-M-1 and blaTEM-1, raising the possibility of transposase-mediated transfer of blaTEM-1 (43) by one of the six isolates that encoded both genes. It is also possible that significant fitness costs were associated with the uptake of other plasmids, leading to our inability to detect them within the cecal community. The gene blaCTX-M-1 has been identified as the most common ESBL gene found in livestock in Germany (44), the country of origin of generalist3. Found on highly transferable IncI1 or IncN plasmids (45, 46), transmission between individual E. coli isolates that can colonize animals or humans has been implied following detection of IncI1 blaCTX-M-1 plasmids in poultry, chicken meat, and human isolates in the Netherlands (47). Collectively, while our study demonstrates the complexities of understanding individual isolate dynamics in mixed-species communities, further studies are necessary to better understand AMR transmission (48).
The chicken cecal fermentation model we have used to observe ESBL transfer in vitro suffers from several limitations. Firstly, it is unlikely that the fermentation vessels contain all the taxonomic diversity present in the ceca due to the loss of strict anaerobes during inoculum preparation and the absence of “re-seeding” due to the transfer of feed particles and microorganisms from the small intestine. Secondly, we have chosen to use a pool of cecal content from 20 chickens to initiate our models instead of content from single birds. This ensured that we had enough cecal content to run all experiments, and this pool could be used to bring uniformity to the bacterial populations so that any effects seen were due to differing experimental conditions (e.g., different cocktail concentrations). We appreciate that this removed the intra-individual variation seen in the chicken cecal microbiota in vivo. Thirdly, there is no host involvement, as it is known that microorganisms within the gut respond to host-derived compounds, metabolites, and cells. Finally, while the fermentation vessels support a continuous flow of material, the cyclical emptying seen in live animals is not reproduced. Regardless, this study highlighted that plasmid transfer events between transient and resident E. coli can be observed in a complex microbiota in vitro. The molecular basis and multiple drivers of bacterial AMR transmission and evolution are difficult to study, but moving beyond a focus on single-species systems using in vitro models will help unveil these mechanisms.
MATERIALS AND METHODS
Continuous flow chicken cecal fermentation model
A modified continuous flow model originally described by Card et al. was used in this study (30). The model system consisted of six individual, unconnected fermentation vessels. Temperature (held at 42°C by a circulating water bath; Optima) and pH (kept within the range of 5.8–6.0 by Mettler-Toledo pH probes and Fermac 260 pH measurement and control modules) were automatically controlled (49). Media, provided via a multi-channel, variable-speed peristaltic pump (Electrolab) at a rate of 1.25 mL/h, supplied the vessels. All vessels and media reservoirs were under constant stirring using magnetic fleas and kept under anaerobic conditions by a continuous flow of anaerobic gas mixture (80% N_2_, 10% CO_2_, 10% H_2_) directly into the vessels and fresh media reservoir. Prior to each run, vessels (18 mL) and reservoirs (1 L) were filled with sterile Viande-Levure culture medium (10 g/L tryptone, 5 g/L NaCl, 5 g/L yeast extract, 2.5 g/L glucose, 2.4 g/L beef extract, 0.6 g/L cysteine.HCl) and left to gas anaerobically overnight.
Collection of cecal contents from chickens was approved by the University of Surrey’s animal ethics committee (NERA-2017-010-SVM). Twenty commercial broiler chickens were euthanized by cervical dislocation, and cecal contents were aseptically collected. Cecal samples were either used immediately as fermentation inoculum or frozen at −80°C for the generation of pooled inoculum. For this, samples from 15 individual animals were subsequently defrosted, combined, manually homogenized, and dispersed into 1 g aliquots for storage at −80°C for later use. Microbial communities were established by adding 2 mL of chicken cecal content slurry (1:1, cecal content:phosphate-buffered saline [PBS]) to each vessel, followed by batch culture for 24 h (no media flow) and 4 days of fermentation (with media flow) to allow stabilization of bacterial communities.
Chicken cecal fermentation model experiments
Inoculum validation experiment
Fresh or frozen, individual or pooled samples of chicken cecal content were assessed as an inoculum source. Duplicate vessels were inoculated with either fresh or frozen cecal material from two chickens or with content pooled from both chickens (Item S9). After 24 h of batch culture and 4 days of fermentation (with media flow), samples were collected, and 16S rRNA gene profiles of microbial communities from the vessels were compared to those of chicken cecal content (see DNA extraction and sequencing for genomic analyses section below).
Selection of isolates in the E. coli “cocktail”
A defined cocktail of 17 host-associated ESBL-producing isolates was generated from a previously described collection of E. coli isolates (18). Host association was determined by using a library of 1,198 E. coli isolates collected in Germany, Spain, the UK, and Vietnam. One hundred fifty phylogenetic trees (bootstrap replicates) were generated, based on the alignment of the core genome of all isolates. For each tree and taking source into account, a host specificity score ranging from 0 to 1 was assigned to represent the extent to which a particular isolate was clustered with other isolates of the same host. For each isolate and host species, the geometric mean over the values per tree was used. The geometric mean approach was chosen to ensure only isolates that consistently clustered with the same-host isolates across most phylogenetic reconstructions received a relatively high host specificity score. The generalist score (which may serve as an indicator of an isolate's ability to colonize multiple host species) was determined by calculating the geometric mean of the host specificity scores for cattle, human, swine, and chicken hosts. In order to select suitable isolates for inclusion in the cocktail, the following criteria were applied: (i) isolates belonged to E. coli sensu stricto (no cryptic clade isolates), (ii) isolates either had a relatively high host specificity score (the top 20% for that host), or if the isolate had no pronounced specificity scores for any host, it was predicted to be a generalist, (iii) isolates carried an ESBL gene, (iv) isolates contained as few known virulence genes as possible and had been isolated preferably from a clinically healthy host, and (v) all countries and time periods were equally represented in the final list. To facilitate recovery from complex communities, rifampicin-resistant mutants were generated and used (31).
Preparation of E. coli cocktail
Single colonies from each isolate were aseptically transferred to separate tubes containing 10 mL LB broth supplemented with 4 µg/mL CTF and 50 µg/mL RIF. After aerobic incubation with shaking for 16 h, cultures were diluted 1:50 into fresh LB broth and grown to an OD_600_ of 0.5 before a further 1:10 dilution into fresh LB broth. Thereafter, pre-determined volumes were used to achieve equivalent isolated cell numbers in the cocktail (Item S10). In the first experiment, where 10^8^ CFU E. coli cocktail bacteria were added to each vessel, the pre-determined culture volumes were centrifuged at 3,000 × g for 3 min, supernatants were removed, and each cell pellet was resuspended in 0.5 mL sterile PBS. All the resuspended culture volumes were combined, and the final inoculum volume was made up to 10 mL with the addition of 1.5 mL PBS. Each vessel received 100 µL of the final mixture, amounting to 10^8^ CFU per vessel (5.88 × 10^6^ CFU of each isolate). In the second experiment, where 10^10^ CFU E. coli cocktail bacteria were added to each vessel, triplicate volumes of each isolate were generated, and each pellet was resuspended in 50 µL PBS. Three sets of inocula were prepared, each containing 50 µL of each isolate and 150 µL PBS to bring the final volume to 1 mL. Each vessel received 1 mL of the final mix, amounting to 10^10^ CFU per vessel (5.88 × 10^8^ CFU of each isolate). One milliliter of fermentation media was removed from each vessel prior to cocktail addition.
E. coli cocktail experiments
To assess the relative persistence and conjugative ability of isolates in the E. coli cocktail, several fermentation experiments were performed. In the first experiment, vessels were inoculated with 10^8^ CFU E. coli cocktail or sterile PBS as a control. In the second experiment to explore the impact of inoculum dose, three vessels were inoculated with 10^8^ (low, L) or 10^10^ (high, H) CFU E. coli cocktail. Both experiments were performed in duplicate, providing a total of six replicate vessels in the first (C1-6 control and V1-6 E. coli cocktail) and in the second (L1-6 low dose and H1-6 high dose) experiment. In the third experiment, CTF at 4 or 20 µg/mL was added to two vessels, with two control vessels (CON) receiving sterile PBS. CTF was added to the vessels 24 h before, immediately before, and 24 h after the addition of the E. coli cocktail (10^8^ CFU) to mimic a 3-day dosing regimen. Duplicate experiments were performed, giving four replicate vessels of each condition: 4C A-D contained 4 µg/mL CTF, 20C A-D contained 20 µg/mL CTF, and CON A-D contained PBS only.
Culture-based analyses
In all experiments, samples were taken from the vessels at the following time points: immediately before the E. coli cocktail addition (0 h) and 3, 6, 24, 48, and 72 h post-inoculation. In addition, for the third experiment, a sample was taken immediately before the first dose of antibiotics (−24 h).
Bacterial enumeration
Samples were serially diluted and plated using the Miles and Misra method (50) onto LB agar (for determination of total aerobic bacteria), Gassner agar (for total Enterobacteriaceae), Gassner supplemented with 4 µg/mL CTF (for CTF^R^ Enterobacteriaceae), and Gassner supplemented with 50 µg/mL RIF and 4 µg/mL CTF (for E. coli cocktail isolates). Plates were incubated at 35°C for 18 h, counted, and the CFU/mL of each of the four groups of bacteria was calculated. Replica plating was performed to assess if any potential transconjugants were present in the vessels. Colonies from the Gassner agar plates supplemented with CTF were picked and transferred onto fresh Gassner agar plates with CTF and Gassner agar plates with RIF and CTF. Any colonies that did not grow on the RIF and CTF plates but did grow on the CTF plates were deemed to be putative transconjugants and stored at −80°C for further analyses.
Fitting CFU counts under sampling uncertainty
To make optimal use of all count data generated in the experiments, we estimated the most likely CFU concentration per sample while accounting for sampling uncertainty associated with individual CFU counts. As each sample was serially diluted and plated, we obtained, per sample, n replicate measurements X_i_ (1 ≤ i ≤ n) of the CFU count in a certain volume V_i_ (mL). The volume V_i_ represents the effective volume sampled, including a correction for the dilution factor used. We assumed that the CFU count X_i_ follows a Poisson distribution with mean , where C is the actual concentration of cocktail bacteria present in the sample (CFU/mL). The probability mass function, i.e., the chance to find X_i_ CFU given the mean μ_i_, is
The likelihood L(C) of the n observations is
And the corresponding log-likelihood l(C) is given by
The last summation does not depend on the concentration of C and can, therefore, be ignored:
Per sample, we estimated the most likely concentration by maximizing this log-likelihood function. Confidence intervals were computed based on the likelihood ratio, i.e., encompassing all possible values of C where , in which is the quantile function of the chi-square distribution with one degree of freedom, with p being the coverage of the desired confidence interval (e.g., 0.95 for a 95% confidence interval).
Through these calculations, the most likely concentration was calculated per sample, based on all dilutions of that sample. Lower CFU counts, which are associated with higher sampling uncertainty, contribute less than relatively high CFU counts (i.e., at lower dilutions). The total sampling uncertainty around is reflected in its 95% confidence interval.
Detection and analysis of individual members of the E. coli cocktail
PCR-based identification of E. coli isolates
Bacterial growth on Gassner agar supplemented with 50 µg/mL RIF and 4 µg/mL CTF was resuspended in 2 mL sterile PBS and heated at 95°C for 10 min to lyse the cells. DNA was subjected to a quadruplex ORFan gene multiplex PCR assay to assess the presence of the 17 cocktail isolates at each time point as previously described (31). Data were expressed as isolate presence or absence for each of the 17 isolates in each vessel, at each time point.
Non-metric multidimensional scaling of PCR profiles
PCR presence/absence data were analyzed using NMDS to evaluate whether the composition of the E. coli cocktail changed over time, in response to increasing dose, or in the presence of CTF. Entries where no E. coli cocktail isolates were detected by PCR were removed, as these did not provide any information on the community composition. Using the Jaccard index as the distance matrix, the NMDS analysis was run multiple times, increasing the number of dimensions from 2 to 10. Each run with 15 tries (iterative) converges on the best solution based on the lowest stress. Then, based on a threshold stress of 0.1 (a good representation of actual dissimilarities), k = 3 dimensions was selected. An analysis of similarities (ANOSIM) test was performed to test whether the heterogeneity found between groups was significantly larger than the heterogeneity found within the groups. Finally, convex hull plots were created for the first two dimensions, visualizing the clustering of the PCR profiles. The R package vegan (51) was used for all analyses, and ggplot2 (52) was used to generate the visualizations.
DNA extraction and sequencing for genomic analyses
Samples from the cecal fermentation model for DNA extraction were processed by centrifugation (11,337 × g for 10 min), the supernatant was discarded, and DNA was extracted from the cell pellet using the DNeasy Power Soil kit (QIAGEN) in accordance with the manufacturer’s protocol. Eluted DNA was resuspended in 100 µL of DNA-free water.
16S rRNA gene sequencing
A total of 12 DNA extracts from the validation experiment, along with DNA extracted from three chicken cecal content samples, were sent for 16S rRNA gene sequencing on the MiSeq Illumina platform (Animal and Plant Health Agency, Weybridge, UK). The V4 and V5 regions of the 16S rRNA genes were amplified using the primers U515F (5′-GTGYCAGCMGCCGCGGTA-3′) and U927R (5′-CCCGYCAATTCMTTTRAGT-3′) (53). Amplification was performed using the following PCR conditions: 95°C for 3 min, 25 cycles of 95°C for 30 s, 55°C for 35 s, and 72°C for 1 min, followed by 72°C for 8 min. Amplicons were purified using Ampure XP magnetic beads (Beckman Coulter). Each sample was tagged with a unique pair of indices and sequencing primers using Nextera XT v2 Index kits and 2× Kalpa HiFi HotStart ReadyMix. The PCR conditions used for this were: 95°C for 30 s, 55°C for 30 s, 72°C for 30 s, and 72°C for 5 min. The concentration of each sample was quantified using the Quantiflour assay (Promega), and concentrations were normalized before pooling all samples. Sequencing was performed on an Illumina MiSeq 2 × 300 bp (Illumina, Cambridge, UK).
16S rRNA gene sequencing files were processed and analyzed using QIIME2 (qiime2-2018.4) (53). All sequence files were imported and converted into a single QIIME2 data file (qiime tools import). DADA2 (54) was used to denoise and remove low-quality reads at positions 20 and 280. Alignment was performed on the sequences (qiime alignment mafft), and this alignment was masked to remove positions that were highly variable (qiime alignment mask). FastTree was used to generate a phylogenetic tree from the masked alignment (qiime phylogeny fastree), and midpoint rooting was applied (qiime phylogeny midpoint-root). The reference database Greengenes (version 2018) (55) was utilized and trained on the sequences generated from the study (qiime feature-classifier classify-sklearn). Taxonomic composition of all samples and samples by groups was generated (qiime taxa barplot).
Whole-genome sequencing
DNA was extracted from the 17 E. coli cocktail isolates using the Promega Wizard DNA extraction kit (Wisconsin, USA) according to the manufacturer’s instructions. Genomic DNA libraries were prepared using Nextera XT Library Prep Kit (Illumina, San Diego, USA) with the following modifications: input DNA was increased twofold, and PCR elongation time was increased to 45 s. DNA quantification and library preparation were carried out on a Hamilton Microlab Star automated liquid handling system (Hamilton Bonaduz AG, Switzerland). Libraries were sequenced at MicrobesNG (Birmingham, UK) on Illumina NovaSeq 6000 using a 250 bp paired-end protocol. Sequence adapters were trimmed using Trimmomatic (version 0.30) with a sliding cut-off of Q15 and a sliding window of 5 base pairs. De novo assembly was performed using SPAdes V3.7 (56). Contig files were uploaded to the University of Surrey’s remote Linux server and the following analyses carried out: MLST (v2.17.0) (57) to identify the sequence types (ST) of the transconjugants, ParSNP (v1.6) (58) to generate a phylogenetic tree (using auto-picked reference genome of CONB72HTC7), Abricate (v1.0.0) (58) to identify resistance genes (using the NCBI database, release 245) and plasmids (using the PlasmidFinder database) (v2.0.1). Sequencing files from the 16S rRNA gene amplicon sequencing of cecal content and fermentation liquid, along with the whole-genome sequencing of trans-conjugant isolates, can be accessed on the NCBI Sequence Read Archive under BioProject ID PRJNA926526. Some of the genomes of the cocktail isolates were uploaded previously to the NCBI Sequencing Read Archive, and a table including all cocktail isolates, their Bioproject and SRA accession numbers can be found in Item S12.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ercumen A, Pickering AJ, Kwong LH, Arnold BF, Parvez SM, Alam M, Sen D, Islam S, Kullmann C, Chase C, Ahmed R, Unicomb L, Luby SP, Colford JM Jr. 2017. Animal feces contribute to domestic fecal contamination: evidence from E. coli measured in water, hands food, flies, and soil in Bangladesh. Environ Sci Technol 51:8725–8734. doi:10.1596/3136928686435 PMC 5541329 · doi ↗ · pubmed ↗
- 2Zhang C, Derrien M, Levenez F, Brazeilles R, Ballal SA, Kim J, Degivry M-C, Quéré G, Garault P, van Hylckama Vlieg JET, Garrett WS, Doré J, Veiga P. 2016. Ecological robustness of the gut microbiota in response to ingestion of transient food-borne microbes. ISME J 10:2235–2245. doi:10.1038/ismej.2016.1326953599 PMC 4989305 · doi ↗ · pubmed ↗
- 3Martínez I, Muller CE, Walter J. 2013. Long-term temporal analysis of the human fecal microbiota revealed a stable core of dominant bacterial species. P Lo S One 8:e 69621. doi:10.1371/journal.pone.006962123874976 PMC 3712949 · doi ↗ · pubmed ↗
- 4Martinson JNV, Pinkham NV, Peters GW, Cho H, Heng J, Rauch M, Broadaway SC, Walk ST. 2019. Rethinking gut microbiome residency and the Enterobacteriaceae in healthy human adults. ISME J 13:2306–2318. doi:10.1038/s 41396-019-0435-731089259 PMC 6776003 · doi ↗ · pubmed ↗
- 5Kim W, Racimo F, Schluter J, Levy SB, Foster KR. 2014. Importance of positioning for microbial evolution. Proc Natl Acad Sci USA 111:E 1639–E 1647. doi:10.1073/pnas.132363211124715732 PMC 4000849 · doi ↗ · pubmed ↗
- 6Conway T, Cohen PS. 2015. Commensal and pathogenic Escherichia coli metabolism in the gut. Microbiol Spectr 3. doi:10.1128/microbiolspec.MBP-0006-2014 PMC 451046026185077 · doi ↗ · pubmed ↗
- 7Delmas J, Gibold L, Faïs T, Batista S, Leremboure M, Sinel C, Vazeille E, Cattoir V, Buisson A, Barnich N, Dalmasso G, Bonnet R. 2019. Metabolic adaptation of adherent-invasive Escherichia coli to exposure to bile salts. Sci Rep 9:2175. doi:10.1038/s 41598-019-38628-130778122 PMC 6379400 · doi ↗ · pubmed ↗
- 8Niehus R, Oliveira NM, Li A, Fletcher AG, Foster KR. 2021. The evolution of strategy in bacterial warfare via the regulation of bacteriocins and antibiotics. Elife 10:e 69756. doi:10.7554/e Life.6975634488940 PMC 8423443 · doi ↗ · pubmed ↗