Impact of Asaia bacteria on Leishmania major development in sand flies: implications for vector control strategies
Marketa Stejskalova, Magdalena Jancarova, Katerina Pruzinova, Kristina Capova, Ilaria Varotto-Boccazzi, Sara Epis, Petr Volf

TL;DR
This study explores how Asaia bacteria affect Leishmania parasites in sand flies, suggesting potential for controlling disease spread.
Contribution
First demonstration of Asaia bacteria colonizing and vertically transmitting in Phlebotomus duboscqi sand flies.
Findings
Asaia siamensis and A. krungthepensis colonized sand fly guts and transmitted vertically.
These bacteria reduced Leishmania major infection intensity by day 8 post-blood meal.
A. krungthepensis increased metacyclic Leishmania forms, while AsaiaWSP had no significant effect.
Abstract
Asaia spp., bacteria originally isolated from tropical plants, have also been identified in various insect species, including blood-feeding ones. Their ability to colonize different host tissues and transmit vertically between generations makes these bacteria good candidates for paratransgenesis. However, most existing data derived from studies on mosquitoes and other important vectors, such as phlebotomine sand flies (Diptera: Psychodidae), remain understudied. In this study, we investigated the ability of wild-type Asaia siamensis, Asaia krungthepensis, and a genetically modified strain of Asaia expressing the Wolbachia surface protein (AsaiaWSP) to colonize Phlebotomus duboscqi. In addition, we studied their vertical transmission and their interactions with Leishmania major during superinfection. Phlebotomus duboscqi females were provided with Asaia via a sugar meal. Bacterial…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
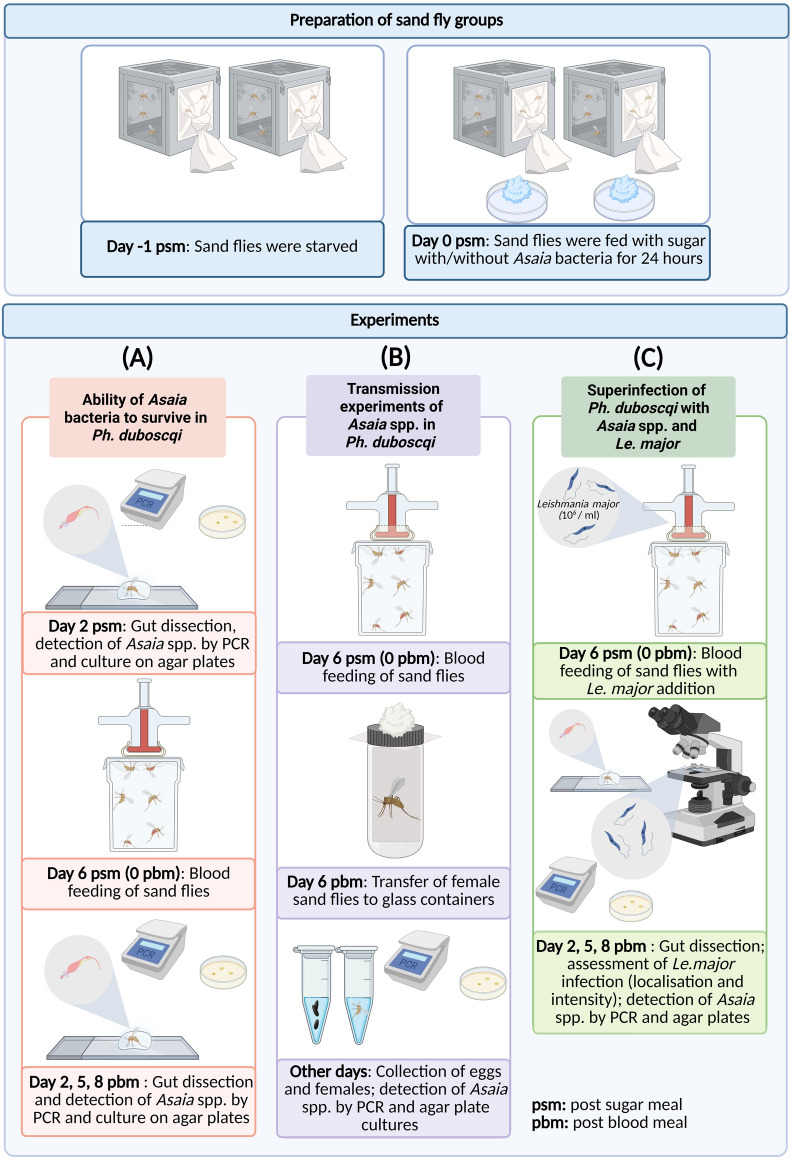 Figure 1
Figure 1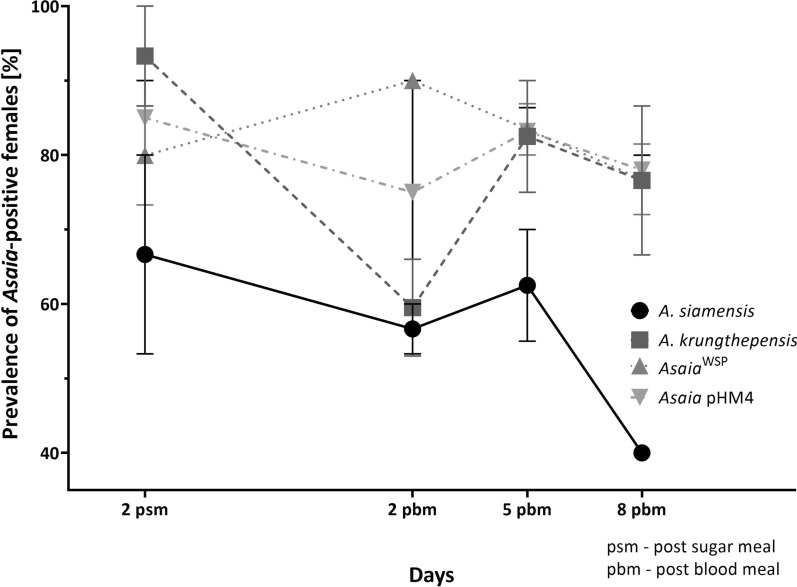 Figure 2
Figure 2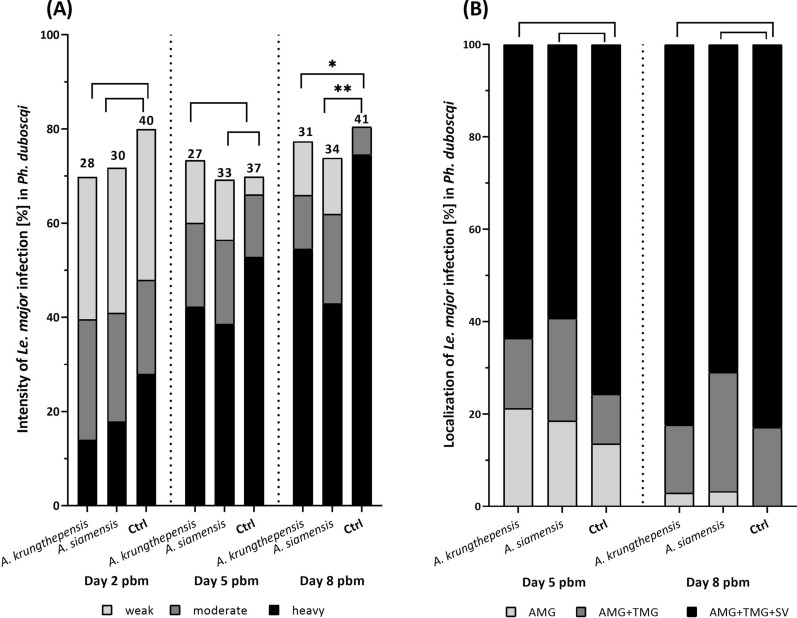 Figure 3
Figure 3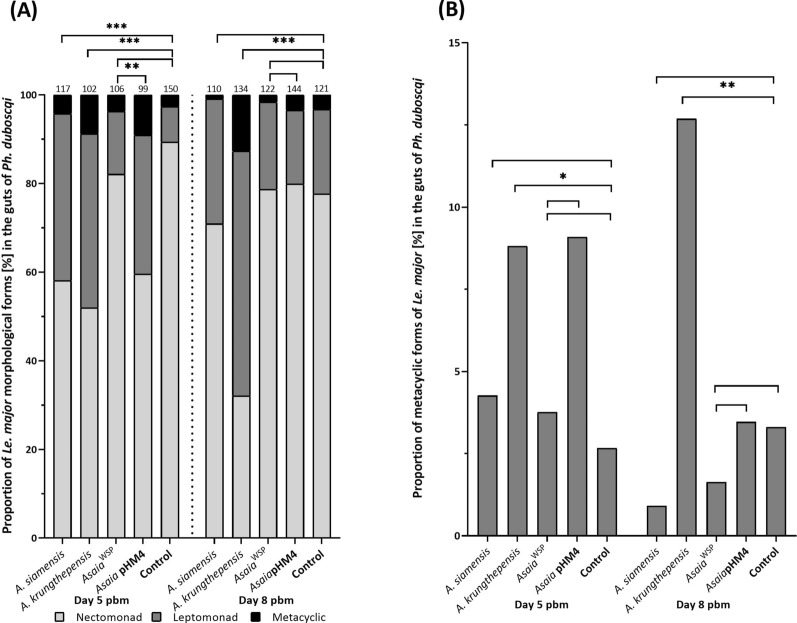 Figure 4
Figure 4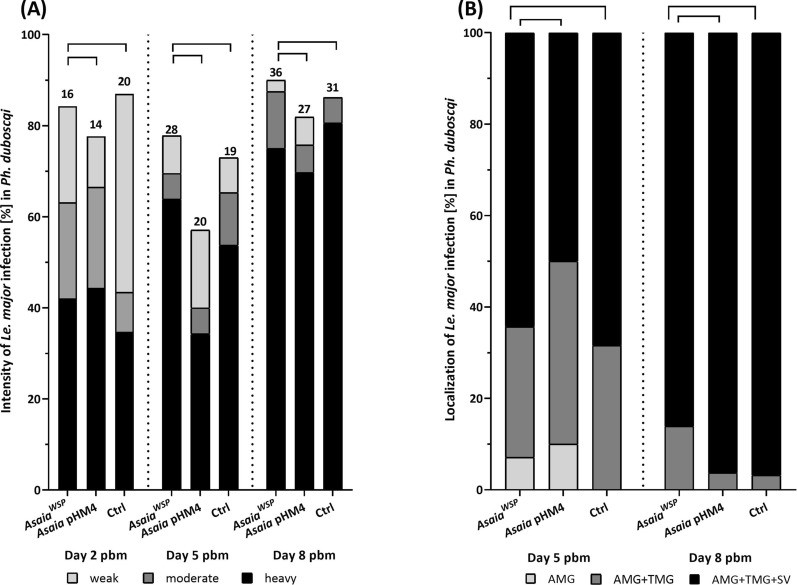 Figure 5
Figure 5 Figure 6
Figure 6- —National Institute of Virology and Bacteriology. Funded by the European Union – Next Generation EU.
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect symbiosis and bacterial influences · Insect and Pesticide Research · Insect-Plant Interactions and Control
Background
In arthropods, microorganisms play a crucial role in various aspects of their biology and life cycle, including nutrition, development, reproduction, immune response, behavior, defense against natural enemies, and speciation (reviewed by [1–3]). From a host’s perspective, interactions with microorganisms can be classified as pathogenic, when the microbes reduce their host fitness (i.e., antagonistic interactions); neutral, with no effect on the host fitness (i.e., commensal interactions); or beneficial, when the microbes enhance host fitness (i.e., mutualistic interactions) [4].
Acetic acid bacteria of the genus Asaia, originally isolated from tropical plants [5–7], are also able to colonize sugar-feeding insects from phylogenetically distant taxonomic groups [8–10]. Among blood sucking arthropods, Asaia sp. were detected in different mosquito and sand fly species [8, 11–16], reduviids [17], Culicoides spp. midges, and the tick Haemaphysalis longicornis [18]. The potential infectivity and risk of Asaia spp. to humans has also been raised as a concern, given that several reports have documented human infections, likely in the context of opportunistic infections, as Asaia does not circulate or establish stable transmission within the human population [19–25]. The most detailed insights into Asaia come from studies conducted on mosquitoes, where Asaia has been identified as a highly promising candidate for the paratransgenic control of vector-borne diseases [11, 26–28]. These bacteria colonize multiple mosquito tissues, including the gut, crop, salivary glands, and reproductive organs of both male and female mosquitoes [11, 12, 29, 30]. Asaia is transmitted between mosquitoes of the genus Anopheles during copulation, vertically from mother to offspring through egg-smearing, and horizontally during cofeeding [11, 12, 29]. Nevertheless, relatively little is known about the physiological role of this bacterium in mosquitoes. In addition to being a component of the microbiome, Asaia influences Anopheles mosquito larval development and adult longevity [31–33], causes transcriptomic changes [32, 33], and induces the activation of mosquito immunity [34]. However, the underlying mechanisms of these effects remain unknown.
Sand flies (Diptera: Psychodidae) are small nocturnal insects; both sexes feed on plant sap, nectar, and honeydew, while females also take blood meals on various vertebrate hosts. These insects have significant veterinary and medical importance as vectors of various pathogens infecting humans or domestic and wild animals. The most important sand-fly-borne pathogens are Leishmania spp., the causative agents of leishmaniasis (reviewed in [35]). There are three main clinical forms of the disease: (i) visceral, which is almost always fatal without treatment; (ii) cutaneous, which causes skin ulcers; and (iii) mucocutaneous, which leads to partial/total destruction of the nose, mouth, and throat mucous membranes. Every year, there are about 700,000 to a million new cases [36]. Although Asaia sp. have been repeatedly found in sand flies [13, 37], their effect on sand flies or transmitted pathogens is unknown.
As far as we know, this is the first study to examine in such detail the tripartite interactions between sand flies, Asaia sp., and Leishmania in vivo. Firstly, we infected females of Phlebotomus duboscqi, a vector of cutaneous leishmaniasis, with Asaia krungthepensis and Asaia siamensis, monitoring their survival and transmission to next generations of sand flies. Subsequently, we assessed the effect of these bacteria on the development of Leishmania major. In addition to using wild-type Asaia strains, we also investigated the effect of Asaia spp. expressing the Wolbachia surface protein (WSP) [28, 38].
Methods
Sand flies
The laboratory colony of Phlebotomus duboscqi (originally from Senegal) was maintained at the Laboratory of Vector Biology at Charles University in Prague under standard conditions [39]. Briefly, sand flies were kept at a temperature of 26 °C, a 14/10 light/dark photoperiod, and had access to a 50% sucrose solution ad libitum.
Leishmania
Leishmania major LV561 (MHOM/IL/67/LRC-L137 Jericho II) was cultured at 23 °C in M199 medium (Sigma-Aldrich) supplemented with 10% fetal bovine serum (Gibco), 1% BME vitamins (Sigma-Aldrich), 2% filtered human urine, and amikacin (250 μg/mL).
Bacteria
Four Asaia strains were used. Wild strains of Asaia siamensis (7132^T^) and Asaia krungthepensis (7333^T^) [6, 40] were provided by the Czech Collection of Microorganisms (Brno, Czech Republic). The genetically modified Asaia bacteria were kindly provided by Prof. Sara Epis from the University of Milan. The gene encoding the Wolbachia surface protein (WSP) was inserted into plasmid pHM4 and introduced into Asaia SF2.1, a wild-type strain derived from mosquitoes, resulting in the recombinant strain Asaia^WSP^. The control strain, Asaia pHM4, carried the empty plasmid [28]. Asaia spp. were cultivated on GLY medium (25 g/L glycerol [Lach-Ner s.r.o.], 10 g/L yeast extract, pH 5; Sigma-Aldrich). Agarose (20 g/L) was added when a solid medium was required to maintain colonies and detect bacteria during experiments. Kanamycin (100 µg/mL) was added to the medium to grow genetically modified bacteria carrying the plasmid*.*
Bacteria preparation for sugar meal infection
Bacterial strains, regardless of species, were inoculated from agar plates into liquid GLY medium and incubated at 30 °C with shaking at 200 rpm until reaching an optical density corresponding to an infectious dose in the range of 1.8–4 × 10^8^ CFU/mL. Afterwards, the cultures were centrifuged twice at 6500 rpm for 5 min and washed with physiological saline solution to stop further bacterial growth. Separately, a 20% solution of cane sugar and distilled water was boiled in a microwave oven three times. The blue food coloring (AROMA a.s.) was added to the cooled solution, which was filtered using a sterile membrane filter (0.22 µM, Millipore Millex). The resulting bacterial pellets were mixed with a 20% sucrose solution, rather than the 50% sucrose used for colony maintenance, to minimize osmotic pressure. The mixture of pellet and sugar was stained with a food dye, which allowed visual confirmation of successful feeding.
Experimental infection of sand flies with Asaia spp.
In each experiment, 120–150 Ph. duboscqi females (1–3 days old) were first starved for 24 h before the experiment and then offered a mixture of sugar solution with Asaia bacteria for another 24 h. Unfed females were removed from the cage, and infected females were maintained in standard conditions with access to a 50% sucrose solution (noninfected). On the second day post sugar meal (psm), 15–30 females per group were dissected to confirm the ability of Asaia spp. to survive in the digestive tract of Ph. duboscqi (Fig. 1A). Females were anesthetized on ice, and their legs were removed. To minimize external contamination, the surface of each sand fly was washed with distilled water containing a small amount of mild detergent, rinsed twice in distilled water, immersed in 70% ethanol for 15 s, and finally washed again with physiological saline. Then, females were dissected in a drop of phosphate-buffered saline (PBS) under a binocular microscope using a clamp and dissecting tools; the head was removed, and the gut (midgut plus hindgut with Malpighian tubules) was pulled out from the abdomen. Each gut was rinsed in a fresh drop of physiological saline, transferred individually into a microcentrifuge tube containing 100 µL of sterile physiological saline, and then manually homogenized with a pestle. From each homogenate sample, 10 µL was inoculated onto agar, and the remaining volume was used for PCR testing to verify both the presence and viability of the bacteria. A sample was considered positive if Asaia DNA was detected by PCR and bacterial growth was observed on solid medium (Fig. 1A).Fig. 1. Experimental design of Asaia spp. colonization and subsequent superinfection with Leishmania major in female Phlebotomus duboscqi. In all experiments, females of Phlebotomus duboscqi were starved (−1 psm) and then on day 0 psm fed for 24 h on a control sugar solution or a sugar with bacteria of the genus Asaia (0 psm) (top panel). Three experiments were conducted on the group (A) ability of Asaia bacteria to survive in Ph. duboscqi, (B) transmission experiments of Asaia spp. in Ph. duboscqi, and (C) superinfection of Ph. duboscqi with Asaia spp. and Le. major. This schematic was created with BioRender.com
On day 6 psm, Ph. duboscqi females were fed for 60–120 min with heat-inactivated sheep blood (LabMediaServis) through a chicken skin membrane on a glass feeder as described by Volf and Volfova [39] (day 0 post blood meal, i.e., day 0 pbm). On days 2, 5, and 8 pbm, 10–30 females from each group were dissected and processed as described above. The experiments were repeated three times for wild-type strains and twice for genetically modified bacteria.
Molecular detection of Asaia spp.
DNA was isolated from homogenized samples using the High Pure PCR Template Preparation Kit (Roche) according to the manufacturer’s protocol. PCR reactions were performed in a total volume of 20 μL, containing 10 μL Emerald Amp GT PCR Master Mix Green (TaKaRa BIO INC.), 1 μL forward primer ASAFOR (5′-GCGCGTAGGCGGTTTACAC-3′; Sigma-Aldrich), 1 μL reverse primer ASAREV (5′-AGCGTCAGTAATGAGCCAGGTT-3′; Sigma-Aldrich), [11], and 2 μL template DNA and nuclease-free water to make up the final volume. The amplification protocol consisted of an initial denaturation at 95 °C for 2 min, followed by 30 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 60 s, and extension at 72 °C for 60 s, with a final extension step at 72 °C for 4 min. The resulting PCR product was analyzed by agarose gel electrophoresis. PCR products were loaded onto a 1% agarose gel prepared from agarose powder (BioReagent, Sigma-Aldrich), 50× TAE electrophoresis buffer (Thermo Fisher Scientific), and SYBR Safe DNA Gel Stain (Thermo Fisher Scientific), which allows DNA visualization. For each sample, 10 μL of PCR product was loaded into the wells of the gel. The first and last wells were loaded with 10 µL of GeneRuler 100 bp DNA Ladder (Thermo Fisher Scientific) as a size standard. Electrophoresis was performed at 90–120 V for 20–55 min, depending on the size of the gel. The gel was visualized and photographed under blue light using a VILBER imaging system and then analyzed.
Asaia spp. detection by cultivation
In total, 10 µL of homogenate were inoculated onto an agar plate with GLY medium and spread with a sterile glass stick over the entire surface to obtain separate colonies. The plates were then incubated in a thermostat for 48 h at 30 °C.
Transmission experiments of Asaia spp. in Ph. duboscqi
To investigate the possibility of transovarial transmission of Asaia spp. in Ph. duboscqi, three experimental groups of 150 females were established. The females were fed on sugar with (1) A. krungthepensis, (2) A. siamensis, or (3) without bacteria (control group). On day 6 psm the sand flies were fed on the blood (0 pbm) (Fig. 1B). On day 6 pbm, individual females were placed into glass vials containing filter paper moistened with distilled water, and the inlet was covered with monofilament. Vials with females were placed in a plastic box, which was also lined with moist filter paper, and maintained at 26 °C and 70% relative humidity. Viability of A. siamensis and A. krungthepensis in females was controlled as described above. As soon as the female died, its carcass and laid eggs were separately sampled, homogenized, and tested by PCR, and inoculated on agar plates. Again, positive detection by PCR must be accompanied by positive cultivation. The experiment was repeated twice.
Vertical transmission of Asaia spp.
After laying eggs, dead females were removed, and the eggs were divided into four groups: (1) nonhomogenized, unwashed clutches; (2) homogenized, washed clutches; (3) homogenized, unwashed clutches; and (4) nonhomogenized, washed clutches. The washing solution was prepared using a modified protocol by Poinar and Thomas [41]. Briefly, eggs were rinsed in distilled water, immersed in 500 µL of 70% ethanol for 5 min, and then in a 10% solution of sodium hypochlorite for the same period. After each chemical treatment, the eggs were rinsed in distilled water for 5 min to remove residual disinfectants. In addition, groups 2 and 3 were homogenized for 5 min using a homogenizer with an iron bead to assess whether Asaia bacteria were localized inside the eggs or merely adhered to their surface. The presence of Asaia in the homogenate was again evaluated by both PCR and agar plates. The experiments were repeated twice.
Superinfection of Ph. duboscqi with Asaia spp. and Leishmania major
Groups of 120–150 Ph. duboscqi females (1–3 days old) were first infected with Asaia spp. using an infection dose of 1.8–4 × 10^8^ CFU/mL via sugar feeding. On day 6 psm/0 pbm, Ph. duboscqi females were fed for 60–120 min with heat-inactivated sheep blood (LabMediaServis), seeded with 1 × 10^6^ cells/mL promastigotes of Le. major, through a chicken skin membrane on a glass feeder. Non-blood-fed females were removed, and blood-fed ones were maintained at standard conditions. Between 15 and 30 Ph. duboscqi females were dissected on days 2, 5, and 8 post blood meal (pbm). Each female was washed as described above, and then the gut was dissected into a drop of PBS under a binocular microscope. The guts were examined microscopically for the presence, intensity, and localization of Leishmania infection. The intensity of infection was graded as (1) no infection, (2) weak (1–100 parasites/gut), (3) moderate (100–1000 parasites/gut), and (4) heavy (> 1000 parasites/gut) [42]. Three different localizations were distinguished, Leishmania either stayed in the abdominal midgut (AMG) or migrated anteriorly and also colonized the thoracic midgut (TMG) and stomodeal valve (SV). Subsequently, the dissected guts were rinsed from the microscope slide and coverslip using a pipette with 100 μL physiological saline solution, collected into a microtube, and homogenized. The homogenate was used for cultivation on agar plates and for PCR analysis to detect and confirm the presence of viable Asaia spp.
In addition, to evaluate Leishmania morphological forms present in the midgut, smears were made from Le. major positive guts using a coverslip. After drying, the sample was fixed with methanol and stained with Giemsa (Sigma-Aldrich).
Promastigotes of Le. major were classified as metacyclic forms when flagellum length was ≥ 2 times body length, leptomonad forms when flagellum length was < 2 times body length and body length was < 14 μm, and elongated nectomonads when flagellum length was < 2 times body length and body length was ≥ 14 μm according to [43]. The experiments were repeated three times for wild-type strains and twice for genetically modified bacteria (Fig. 1C).
Statistical evaluation
The data were tested in StudioR (http://cran.r-project.org) [44]; owing to the type of data, Fisher’s exact or Chi-squared tests were used to assess the statistical significance.
Results
Colonization of Ph. duboscqi with Asaia spp.
All four Asaia strains tested (A. siamensis, A. krungthepensis, Asaia^WSP^, and Asaia pHM4) successfully colonized the gut of Ph. duboscqi following both the sugar meal (psm) and the subsequent blood meal (pbm) throughout the experimental period (Fig. 2). The lowest colonization was observed for A. siamensis, with infection rates of 66.7%, 56.7%, 62.5%, and 40% on day 2 psm and days 2, 5, and 8 pbm, respectively. Asaia krungthepensis showed a high initial infection rate on day 2 psm (93.3%), which dropped to 60% on day 2 pbm but increased again to nearly 80% on days 5 and 8 pbm, following blood defecation. A similar trend was observed for Asaia pHM4, though with slightly lower infection rates ranging from 71% to 85%. In contrast, Asaia^WSP^ maintained relatively stable and high infection rates, varying between 77% and 90% across all tested time points (Fig. 2).Fig. 2. Prevalence of Asaia-positive females over time. Asaia strains (A. siamensis, A. krungthepensis, Asaia^WSP^, and Asaia pHM4) were tested for colonization of Ph. duboscqi midgut on various days post sugar meal (psm) and post blood meal (pbm). The data are shown as prevalence with the standard error of the mean (SEM), which originates from two independent experiments for genetically modified Asaia and three independent experiments for wild-type Asaia
Vertical transmission of wild-type Asaia spp.
Out of 128 females infected with A. siamensis, 78 were positive and laid eggs, yielding 28 positive clutches. The highest positivity (80%) was detected in egg clutches laid on day 9 pbm (Fisher’s exact test, P = 0.001). Out of 128 females infected with A. krungthepensis, 69 were positive and laid eggs, and 41 of the clutches were infected. The positivity of egg clutches (Table 1) did not differ significantly across various days (Fisher’s exact test, P = 0.824) but was significantly higher compared with A. siamensis (χ^2^ = 7.2168, df = 1, P = 0.007). Table 1. Summary of the number of Asaia positive/negative egg clutches per day after blood meal (pbm) laid by Asaia-infected Ph. duboscqi femalesDay pbm67891011121314Total (%)A. siamensisPositive46084222028 (36%)Negative166622806450 (64%)A. krungthepensisPositive57553534441 (59%)Negative26274221228 (41%)
In both Asaia species the vertical transmission was through egg-smearing. Regardless of homogenization, all test groups without egg washing were PCR positive, while washed eggs were all negative.
Leishmania major infection in Ph. duboscqi infected by wild-type Asaia spp.
On day 2 pbm, i.e., before defecation, Le. major promastigotes were present in the endoperitrophic space, and the percentage of infected females ranged between 70% and 80% across all three experimental groups (Fig. 3A). Both the infection rates and parasite loads (i.e., infection intensities) did not significantly differ between females in the control group (without Asaia) and those colonized by A. siamensis (χ^2^ = 0.426, df = 1, P = 0.514; Fisher’s exact test, P = 0.729), nor between the control and the group colonized by A. krungthepensis (χ^2^ = 0.809, df = 1, P = 0.369). Similarly, on day 5 pbm (i.e., after defecation), no significant differences were observed among the groups in either the percentage of Leishmania-infected females or parasite loads (A. siamensis: χ^2^ = 0, df = 1, P = 1; A. krungthepensis: χ^2^ = 0.026, df = 1, P = 0.873; Fig. 3A) or intensity of infection (Fig. 3A) (A. siamensis: Fisher’s exact test, P = 0.297; A. krungthepensis: Fisher’s exact test, P = 0.318).Fig. 3. Intensity (A) and localization (B) of Leishmania major infection in Phlebotomus duboscqi females superinfected with wild-type Asaia strains. (A) The intensity of Le. major infection (%) in Ph. duboscqi females on days 2, 5, and 8 pbm. Three groups of females were compared: those infected with A. siamensis or A. krungthepensis and an *Asaia-*negative control group. Numbers above bars indicate the number of Leishmania–Asaia-positive females in each group. (B) Localization of infection on days 5 and 8 pbm. Only Leishmania-positive females from panel A were included; three categories of localization were distinguished: abdominal midgut (AMG), thoracic midgut (TMG), and stomodeal valve (SV). Data originate from three independent biological replicates
In late-stage infections (day 8 pbm), colonization by A. siamensis significantly reduced the intensity of Le. major infection (Fisher’s exact test, P = 0.002), but the percentage of infected females did not differ significantly (χ^2^ = 0.256, df = 1, P = 0.613). A similar effect was observed with A. krungthepensis, which also significantly reduced the incidence of heavy infections (Fisher’s exact test, P = 0.038), with no difference in percentage of infected females (χ^2^ = 0.014, df = 1, P = 0.905; Fig. 3A).
Regarding infection localization (Fig. 3B), neither Asaia strain had a significant effect on any of the days tested. For A. siamensis, no differences were found on day 5 pbm (Fisher’s exact test, P = 0.538) or day 8 pbm (Fisher’s exact test, P = 0.410). Similarly, for A. krungthepensis, no significant effect was observed on day 5 pbm (Fisher’s exact test, P = 0.705) or day 8 pbm (Fisher’s exact test, P = 0.839; Fig. 3B).
As reported in Fig. 4A, three morphological forms of Leishmania (long nectomonads, leptomonads, and metacyclics) were identified on Giemsa-stained smears from Ph. duboscqi midguts colonized by Asaia. On day 5 pbm, colonization with A. siamensis and A. krungthepensis resulted in a significantly increased proportion of leptomonads at the expense of long nectomonads in both groups (Fisher’s exact test, P = 0.000). The proportion of metacyclic promastigotes was significantly higher in the group infected with A. krungthepensis (9%) compared with the Asaia-negative control group (3%). On day 8 pbm, females colonized with A. krungthepensis maintained the elevated proportion of leptomonads (Fisher’s exact test, P = 0.000), whereas no significant differences in the leptomonad-to-nectomonad ratio were observed between the A. siamensis group (Fisher’s exact test, P = 0.277) and the Asaia-negative control group (Fig. 4A). The proportion of metacyclic forms remained significantly higher in the A. krungthepensis group 17% compared with the control 4% (Fisher’s exact test, P = 0.011).Fig. 4. Morphological forms of Leishmania major in Phlebotomus duboscqi females superinfected with Asaia spp. (A) Proportions of three morphological forms of Le. major on days 5 and 8 pbm in sand fly females infected by two wild type strains (A. siamensis and A. krungthepensis), two genetically modified strains (Asaia^WSP^ and Asaia pHM4), and a control group (without Asaia). (B) Percentage of metacyclic promastigotes in these five groups. Numbers above the bars indicate the total number of measured Le. major morphological forms
Leishmania major infection in Ph. duboscqi infected by genetically modified Asaia
On day 2 pbm, the Leishmania infection rates in sand fly groups superinfected with Asaia^WSP^ and Asaia pHM4 were 84% and 77%, respectively comparable to the infection rate observed in the control group without Asaia at 87% (Fisher’s exact test, P = 0.834; Fig. 5A). On days 5 and 8 pbm, Leishmania infection rates in females superinfected with Asaia^WSP^ and Asaia pHM4 fluctuated between 57 and 90%; however, no significant differences were detected compared to the Asaia-negative control (5 pbm: Fisher’s exact test, P = 0.1657; 8 pbm: P = 0.5731; Fig. 5A).Fig. 5. Intensity (A) and localization (B) of Leishmania major infection in Phlebotomus duboscqi females superinfected with genetically modified Asaia strains. A: The intensity of Le. major infection (%) in Ph. duboscqi females on days 2, 5, and 8 pbm. Three groups of females were compared: infected with Asaia pHM4, Asaia^WSP^, and an *Asaia-*negative control group. Numbers above bars indicate the number of Leishmania–Asaia-positive females in each group. B Localization of infection on days 5 and 8 pbm. Only Leishmania-positive females from panel A were included; three categories of localization were distinguished: abdominal midgut (AMG), thoracic midgut (TMG), and stomodeal valve (SV). Data originate from two independent biological replicates
Asaia^WSP^ did not significantly affect the intensity of Le. major infection in any of the tested time points when compared either with the control group without bacteria (Fisher’s exact test, day 2 pbm: P = 0.131, day 5 pbm: P = 0.796, and day 8 pbm: P = 0.599) or with the group infected with Asaia pHM4 (Fisher’s exact test, day 2 pbm: P = 0.904, day 5 pbm: P = 0.078, and day 8 pbm: P = 0.565).
Similarly, there were no significant differences in Le. major localization among Ph. duboscqi sand flies infected with Asaia^WSP^ and Asaia pHM4 (Fisher’s exact test, day 5 pbm: P = 0.222 and day 8 pbm: P = 0.280), or in the control group without Asaia (Fisher’s exact test, day 5 pbm: P = 0.828 and day 8 pbm: P = 0.354; Fig. 5B). On day 5 pbm, sand flies infected with Asaia^WSP^ showed a higher proportion of leptomonad and fewer nectomonad forms compared with those infected with Asaia pHM4 (Fisher’s exact test, P = 0.005; Fig. 4A), in contrast to the comparison between Asaia^WSP^ and the group without Asaia (Fisher’s exact test, P = 0.409). On day 8 pbm, no significant differences were observed between the Asaia^WSP^ group and the control group without Asaia (Fisher’s exact test, P = 0.912), nor between Asaia^WSP^ and Asaia pHM4 (Fisher’s exact test, P = 0.780; Fig. 4B).
The proportions of metacyclic forms of Le. major in Asaia^WSP^-infected sand flies were similar to those in the Asaia-negative control group on day 5 (Fisher’s exact test, P = 0.721) and day 8 pbm (Fisher’s exact test, P = 0.446). Likewise, no significant differences were observed in the proportion of metacyclic promastigotes between Asaia^WSP^ and Asaia pHM4 groups on day 5 (Fisher’s exact test, P = 0.154) or day 8 pbm (Fisher’s exact test, P = 0.458).
Discussion
In our experiments, all four Asaia species and strains tested were able to colonize the midgut of Ph. duboscqi and persisted throughout the observation period, after infection, and following blood feeding and subsequent defecations. However, their infection and survival dynamics differed over time.
The wild-type species, A. siamensis and A. krungthepensis showed similar colonization patterns, a decrease in infection rate following female blood feeding, a slight increase after defecation, and a subsequent decrease. A decline in microbial richness (i.e., the number of distinct operational taxonomic units) after blood feeding in sand flies was described by Kelly et al. [45], nevertheless, they showed that the richness was restored after defecation to a level seen with the sucrose-fed controls. At first glance, this seems inconsistent with our results, as the infection rate of Asaia continued to decline after defecation. However, while Kelly et al. [45] focused on overall species richness, our data specifically concern Asaia. There is a limited information on the behavior of Asaia in blood-feeding insects. Egyirifa and Akorli [46] described a decrease of A. siamensis in blood-fed Anopheles gambiae, and similar trends were observed in females of Aedes aegypti, Aedes albopictus, Mansonia humeralis, and Asaia sp. [47, 48].
Blood feeding alters the gut environment, inducing a shift in the microbiome abundance. Bacteria that thrive within the blood meal possess large genetic redox capacity to cope with oxidative and nitrosative stress associated with blood meal digestion [49]. It seems that wild-type species of Asaia are not among these species and may prefer non-blood-fed guts or other tissues. In our study, both A. siamensis and A. krungthepensis also infected the sand fly reproductive system, facilitating vertical transmission via contamination of the egg surface. Then the bacteria are ingested by the hatching larvae, a mechanism previously suggested for Asaia in mosquitoes [12] and leafhoppers [8].
In sand flies, similar vertical transmission was proven for Psychodiella gregarines [50]; however, to the best of our knowledge, this is the first study to rule out Asaia penetration through the egg membrane and confirm transmission via egg surface contamination.
It has been shown that native Asaia strains can activate mosquito immunity, potentially affecting pathogen development [34, 51]. Asaia introduction triggered mosquito immune responses that reduced Plasmodium berghei development in Anopheles stephensi [34], moreover, precolonization with Asaia sp. significantly reduced Leishmania mexicana populations in Lutzomyia longipalpis [16]. In our study, we tested the effect of A. siamensis and A. krungthepensis on the development of Le. major, the causative agent of zoonotic cutaneous leishmaniasis, in Ph. duboscqi. Both bacterial species significantly reduced parasite load during the late phase of infection. Notably, A. krungthepensis also altered the composition of the Le. major population, particularly increasing the proportion of metacyclic promastigotes. Leishmania development in sand flies proceeds through a series of promastigote morphotypes, among which, metacyclic promastigotes and possibly haptomonads are the main forms transmitted to mammalian hosts during blood feeding [52]. For transmission to mammals, both infection rate and infection intensity are important factors to consider. The presence of Asaia seems to modify this development, when both species decrease the intensity of infection in the late phase; nevertheless, A. krungthepensis stimulates metacyclogenesis.
Whether this effect is due to immune changes, production of anti-leishmanial molecules, microbiome alteration, or a combination of these factors remains to be determined and warrants further research. Nevertheless, the effect of Asaia spp. is less pronounced than that of the Delftia tsuruhatensis TC1 strain, which strongly inhibited Le. major development in Ph. duboscqi; importantly, Leishmania-infected sand flies fed with D. tsuruhatensis were significantly less able to transmit Le. major parasites and cause disease in mice [53].
Regarding the engineered strains, Asaia pHM4 and Asaia^WSP^, we showed that they retain the ability to colonize sand flies. Interestingly, especially in the case of Asaia^WSP^, abundance increased after the blood meal, contrasting with wild strains. Epis and collaborators [28] observed a similar pattern in Aedes aegypti mosquitoes. Despite Asaia^WSP^-induced activation of the host immune response in mosquitoes inhibiting Dirofilaria immitis in Ae. aegypti [28], in the present study, we found no impact on Le. major infection parameters in Ph. duboscqi or Asaia pHM4, and neither did Asaia^WSP^ affect the infection and development of Le. major in Ph. duboscqi.
Wolbachia has been repeatedly shown to activate the immune system in mosquitoes and negatively affect pathogen transmission [54–57]. However, its effect on sand flies remains unclear. Wolbachia have been repeatedly detected in sand flies [37, 58–60], but data on its impact on vector competence are still lacking. Nevertheless, Rosário and collaborators [61] detected Wolbachia and Leishmania infantum coinfections in 37% of Nyssomyia whitmani collected in the field, suggesting that co-existence in the same vector is possible. Similarly, an in vitro study showed that Wolbachia presence in sand fly cells did not significantly impact Le. infantum infection [62]. However, it should be noted that in our experimental model using Asaia^WSP^, the protein came from the filarial nematode Dirofilaria immitis [28], rather than an insect-associated strain. This exogenous WSP was intentionally used to induce a stronger immune response, but it may not accurately reflect the effects of naturally occurring Wolbachia infections in sand flies.
Conclusions
We provided the first evidence that wild-type Asaia bacteria (A. siamensis and A. krungthepensis) can infect Ph. duboscqi and can be vertically transmitted via egg smearing. These bacteria reduce the late-phase infection of Le. major, which may have important epidemiological consequences and can be used for future vector control strategies. Unexpectedly, the Asaia strain expressing Wolbachia surface protein has no measurable effect on Le. major infection in this sand fly species.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1World Health Organization. Leishmaniasis. 2023 https://www.who.int/news-room/fact-sheets/detail/leishmaniasis. Accessed 25 Jun 2025
- 2R Core Team. R: A language and environment for statistical computing R Foundation for Statistical Computing; 2018 https://www.r-project.org/. Accessed 17 Jun 2025